树种和土层对土壤无机氮的影响
2022-12-21韩鑫袁春阳李济宏洪宗文刘宣杜婷李晗游成铭谭波朱鹏徐振锋
韩鑫,袁春阳,李济宏,洪宗文,刘宣,杜婷,李晗,游成铭,谭波,朱鹏,徐振锋
四川农业大学生态林业研究所/长江上游林业生态工程四川省重点实验室/长江上游森林资源保育与生态安全国家林业和草原局重点实验室/华西雨屏区人工林生态系统研究长期科研基地,四川 成都 611130
氮是植物生长过程所必需的营养元素之一,对土壤肥力的形成和植物生长具有重要意义(李志杰等,2017)。土壤氮是森林氮库的重要组成部分,其含量丰缺能反映土壤氮转化与供应能力,并影响植物对氮的利用效率和森林生态系统的生产力(杨起帆等,2021)。土壤氮可通过微生物的生物化学过程转化为无机氮被植物直接利用。土壤无机氮(铵态氮(NH4+-N)和硝态氮(NO3--N))是反映土壤质量和植物生长状况的关键指标,对维持生态系统服务功能和生产力有重大的意义(Li et al.,2020)。研究表明,植物对NH4+-N和NO3--N具有选择吸收的特性,土壤中NH4+-N和NO3--N含量及其比值能反映植物对氮的吸收偏好和吸收效率(张彦东等,2003)。其中,NO3--N能促进植物根系生长,NH4+-N则会抑制植物根系生长,其比值可以反映出近期植物生长状况(Vollbrecht et al.,1992;张彦东等,2003)。因此,研究土壤无机态氮对探究植物生产力和土壤养分利用状况具有重要作用。
土壤无机氮的积累、分布与循环受植物种类、生长环境和土壤性质等综合影响产生差异(戴全厚等,2008)。前人研究表明,植物受到自身化学性质(C/N、木质素含量)影响进而可能会影响土壤氮循环,例如,凋落物分解过程中酚类化合物(如单宁酸)会对土壤硝化、矿化过程和无机氮的积累产生不利影响(Kanerva et al.,2006;张彦东等,2003)。此外,植物根系可以通过分泌物来影响根际微生物的数量与活性,影响植物对土壤氮素的利用效率,进而影响土壤无机态氮含量(丁令智等,2019)。同时,植物生长伴随着根系扩张会驱动土壤团聚体周转,进而改变土壤物理性质(密度和孔隙度),造成土壤NH4+-N和NO3--N含量的差异(Maria et al.,2018)。例如,土壤的黏粒含量将直接决定土壤颗粒表面积的大小和影响土壤透气性,造成 NH4+在土壤团聚体中吸附效果的不同以及土壤中 NO3-的淋溶、气态损失等,最终影响土壤无机氮(Britto et al.,2013)。同时,不同林分由于郁闭度、树木发育等会影响林内微环境,使得土壤温湿度、地温等不同,进而影响到土壤微生物的氮矿化和土壤无机氮(李聪等,2020)。以往有关研究多选择在立地条件相似、土壤基质和树龄大致相近的人工林或采用盆栽试验等来比较树种对土壤无机氮的影响差异,受海拔、林龄、土壤异质性等原因的影响,难以量化树种的真实效应(王薪琪等,2015),同质园试验则能有效避免这种局限性。同质园试验在林龄、土壤基质和经营管理模式相同的均质条件下展开,能有效排除外界干扰环境和原始立地条件差异(Vesterdal et al.,2013)。以往国内外同质园试验多就树种对土壤碳氮养分(Ovington,1954)、水源涵养功能(杨静等,2020)和微生物群落特征(Stefanowicz et al.,2021)等方面的影响展开研究,而就树种对土壤无机氮的影响的研究仍少见报道。
研究区位于四川盆地长江中上游,是中国重要的植被资源和水源涵养保护区,属于亚热带季风气候,夏季雨热同期,降水充沛,植被丰富。合理选择树种造林是重要的林业实践,对提高人工林质量、维持土壤肥力和提升生态效益均具有重要意义(马宏燏等,2017)。常绿阔叶林是四川盆地的主要森林类型,具有生物多样性丰富,生产力高和稳定性强的特点。红椿(T.ciliata)、香椿(T. sinensis)、天竺桂(C. japonicum)、大叶樟(C. phatyphyllum)、香樟(C. camphora)、油樟(C. longipaniculatum)和桤木(A. cremastogyne)是四川盆地常见乡土阔叶树种,具有较高的经济生态效益,因此本研究以上述树种为研究对象,探究树种对不同土壤层次土壤无机氮的影响,以期为今后四川盆地人工造林时选择树种提供科学参考。
1 材料与方法
1.1 研究区域概况
研究区位于四川省崇州市桤泉镇四川农业大学现代农业研发基地(103°49′E,30°55′N)。平均海拔516 m,地势平坦。属亚热带湿润季风气候,年平均日照时数1161.5 h,年均温16.0 ℃,年均降水量约1015.2 mm,平均无霜期285 d。样地内各树种林下植物主要有竹叶草(Oplismenus compositus)和空心莲子草(Alternanthera philoxeroides),其中大叶樟、桤木和香椿样地内随机分布有一两株构树(Broussonetia papyrifera)。撂荒地中主要植物有空心莲子草、茜草(Rubia cordifolia)、狗尾草(Setaria viridis)、蛇莓(Duchesnea indica)和华西凤尾蕨(Pteris occidentalisinica)。
1.2 样地设置
2015年春季,采用随机区组设计,在研究区内建立同质园。同质园内均为人工纯林,造林密度为1600 plant·hm-2。样地前期为农耕地,造林前,对园内土壤进行了旋耕翻土,土壤质地均一,地势平坦。栽植后,除试验前期对样地内树种有浇水来保证树种的成活率,同质园实验区模拟自然森林生长状态,未再做水肥管理。研究选取红椿、香椿、天竺桂、大叶樟、香樟、油樟和桤木7个乡土阔叶树种,每个树种设置了3个重复样地,每块样地面积25 m×25 m,样地间距大于25 m。同时,设置3块撂荒地作为对照。所有树种在栽种时均为2年生幼苗,当年成活率均在90%以上。
1.3 样品采集
2020年7月,在各样方采集土壤样品。每个样方随机选取3个采样点,四分法采样,使用内径5 cm土钻分别采集 0—10、10—20、20—30 cm 土层土壤,将相同样方同一层次样品混合为一个土样后装入消毒自封袋带回实验室,去除石块和动植物残体后过2 mm筛。过筛后一部分置于4 ℃冰箱保存,用于土壤 NH4+-N、NO3--N、土壤微生物生物量碳(MBC)和土壤微生物生物量氮(MBN)测定;另一部分风干后用于含水率、容重、机械组成、孔隙度、pH、C、N、P测定。
1.4 样品测定
土壤NH4+-N和NO3--N分别用氯化钾浸提-靛酚蓝比色法和酚二磺酸比色法测定;土壤MBC和土壤MBN使用氯仿熏蒸K2SO4浸提,过0.45 μm水膜后,由总有机碳分析仪(Multi N/C 2100,Analytik Jena,Germany)测定,以熏蒸样品与未熏蒸样品中可溶性C、N含量的差值表征土壤MBC、MBN,转化系数分别为0.45、0.54;土壤C、N、P含量分别采用重铬酸钾外加热法、凯氏定氮法和酸溶-钼锑抗比色法测定;土壤pH测定采用电位法,水土比 V∶m=2.5∶1;土壤含水率采用烘干法测定;土壤容重和孔隙度使用环刀法测定;土壤砂粒、粉粒和黏粒含量采用马尔文 3000 E激光粒度仪(Malvern Panalytical,Malvern,UK)测定。
1.5 数据分析
采用双因素方差分析(Two-way ANOVA)检验树种和土层及其交互作用对土壤无机氮、NH4+-N和NO3--N及其比值的影响,使用Pearson相关性分析检验NH4+-N、NO3--N及其比值和土壤理化性质的相关性;采用Canoco 5.0软件对影响因素进一步进行冗余分析(RDA),所有统计分析均由 SPSS 19.0完成,显著水平界定P<0.05,所有图片由Origin 2021绘制。
2 结果与分析
2.1 树种和土层对NH4+-N和NO3--N含量的影响
结果表明,树种显著影响土壤NH4+-N含量,且不同树种NH4+-N含量随土层深度变化不同(图1,表1)。除桤木和大叶樟外,其他树种3个土层NH4+-N含量整体均显著低于撂荒地。0—10 cm土层,香樟和红椿NH4+-N含量显著低于其他树种和撂荒地,仅为撂荒地含量的 47.1%和 51.7%。10—20 cm土层,桤木NH4+-N含量显著高于除大叶樟外其他树种,香樟NH4+-N含量相较其他树种最低且显著低于撂荒地,仅为撂荒地含量的47.5%。20—30 cm土层,桤木NH4+-N含量显著高于其他树种,而天竺桂表现为相反趋势。土壤NH4+-N含量整体由高到低为:撂荒地>桤木>大叶樟>油樟>香椿>红椿>天竺桂>香樟
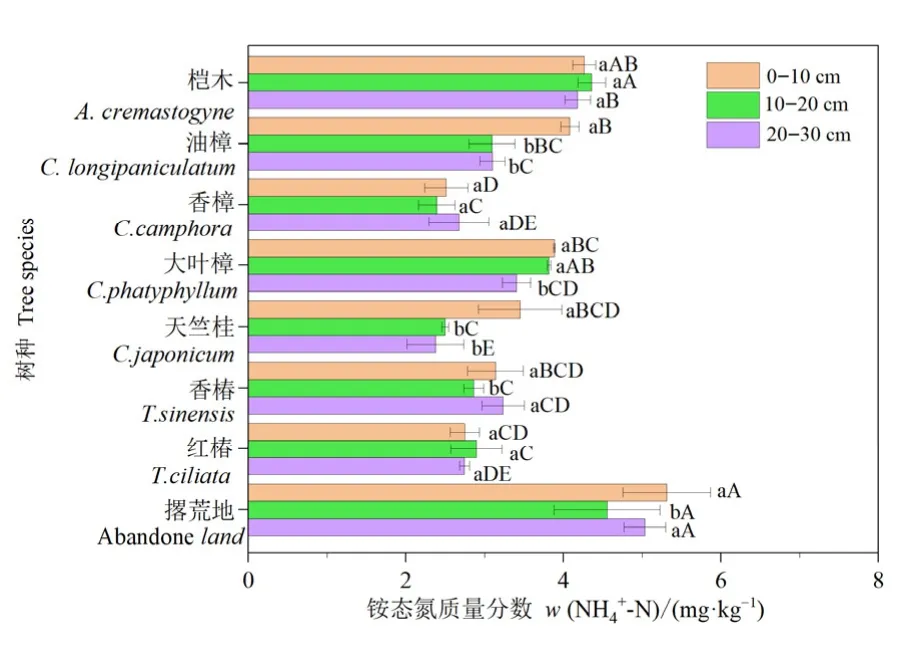
图1 不同树种和土层铵态氮含量Figure 1 NH4+-N content between different tree species and soil layers
土层深度变化对NH4+-N含量的影响在不同树种中表现各异,其中,油樟、大叶樟和天竺桂NH4+-N含量随土层深度的增加而下降,具有表聚现象,而桤木、香樟和红椿NH4+-N含量在3个土层无显著差异。不同土层中,油樟、天竺桂0—10 cm土壤NH4+-N含量显著高于10—20、20—30 cm(图1)。各土层土壤NH4+-N含量平均值分别为3.44、3.13、3.10 mg·kg-1。双因素方差分析表明(表1),树种和土层均显著影响土壤NH4+-N含量且树种效应对其有极显著影响(P<0.001),但树种和土层未对土壤NH4+-N含量产生交互作用。

表1 不同树种和土层对土壤无机氮及其组分的双因素方差分析Table 1 Effects of repeated variance analysis of inorganic nitrogen and its components in different tree species and soil layers
不同树种对土壤 NO3--N含量的影响各不相同,不同深度土层的NO3--N含量变化存在差异(图2;表1)。总体看,除油樟、香樟和香椿外,其他树种各土层NO3--N含量均显著高于撂荒地。0—10 cm土层,天竺桂、红椿和桤木NO3--N含量显著高于其他树种与撂荒地,分别高于撂荒地NO3--N含量的40.0%、37.8%和36.8%,香椿和油樟则显著低于撂荒地。10—20 cm 土层,桤木和红椿 NO3--N含量显著高于其他树种和撂荒地,均高于撂荒地含量的40.6%,香椿NO3--N含量显著低于撂荒地。20—30 cm土层,天竺桂NO3--N含量最高并显著高于撂荒地含量的88.4%。土壤NO3--N含量整体由高到低为:天竺桂>红椿>桤木>大叶樟>香樟>撂荒地>油樟>香椿。

图2 不同树种和土层硝态氮含量Figure 2 NO3--N content between different tree species and different soil layers
土层深度变化显著影响NO3--N含量,如桤木、大叶樟、香椿和红椿的NO3--N含量随土层深度增加而下降,表现出表聚现象(图 2)。不同树种NO3--N含量剖面特征各异,油樟 3个土层间存在显著差异,而香樟3个土层间均无显著差异,天竺桂0—10 cm土壤NO3--N含量显著高于10—20 cm和20—30 cm土壤(图 2)。各土层土壤 NO3--N平均值分别为 22.91、20.22、17.74 mg·kg-1。双因素方差分析表明(表 1),树种、土层及其交互作用(表 1)均对土壤 NO3--N含量有极显著影响(P<0.001)。
2.2 树种和土层对无机氮含量和NH4+-N/NO3--N的影响
结果表明,不同树种和土层显著影响土壤无机氮含量(图 3;表 1)。除油樟、香樟和香椿外,其他树种3个土层无机氮含量均显著高于撂荒地。0—10 cm土层,大叶樟、天竺桂、红椿和桤木无机氮含量显著高于其他树种和撂荒地,相比撂荒地,无机氮含量分别提高了 25.0%、24.5%、19.9%和7.6%。10—20 cm土层,除香椿外,各树种无机氮含量均显著高于撂荒地,其中,大叶樟、桤木和红椿无机氮含量相比撂荒地提高程度大于20.0%,分别为30.8%、23.9%和23.8%。20—30 cm土层,油樟和香椿无机氮含量显著低于撂荒地,其他树种则表现相反,其中,天竺桂无机氮含量最高,比撂荒地含量提升了48.1%。各树种土壤无机氮含量依次为:大叶樟>天竺桂>红椿>桤木>香樟>撂荒地>油樟>香椿。
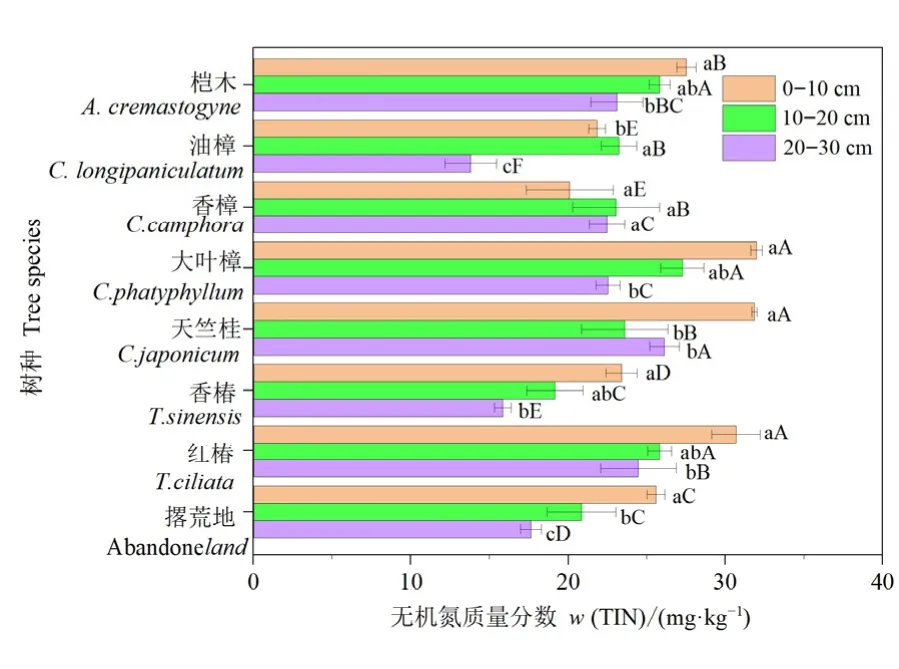
图3 不同树种和土层无机氮含量Figure 3 Total inorganic nitrogen content between different tree species and soil layers
土层深度变化对无机氮含量影响在不同树种间影响不一致,具体表现为桤木、大叶樟、香椿和红椿表现为随土层深度增加而下降,存在明显的表聚现象(图3)。油樟3个土层间均有显著差异,香樟则相反,而天竺桂在0—10 cm土壤无机氮含量显著高于10—20 cm和20—30 cm(图3)。各土层土壤无机氮含量平均值分别为 26.76、23.98、21.19 mg·kg-1。双因素方差分析表明(表 1),树种、土层及其交互作用均对土壤无机氮含量影响极显著(P<0.001)。
树种显著改变了土壤NH4+-N/NO3--N的比值,各树种与撂荒地相比,NH4+-N/NO3--N总体呈下降趋势(图4)。0—10 cm土层,各树种(除桤木外)NH4+-N/NO3--N均显著低于撂荒地。10—20 cm土层,7个树种NH4+-N/NO3--N均显著低于撂荒地。20—30 cm土层,7各树种NH4+-N/NO3--N均显著低于撂荒地,其中香椿NH4+-N/NO3--N又显著高于其他树种,值为 0.323,天竺桂则表现为显著低于其他树种,值为0.103(图4)。整体土壤NH4+-N/NO3--N 值由高到低为:桤木>香椿>油樟>香樟≈大叶樟>红椿>天竺桂。

图4 不同树种和土层铵态氮/硝态氮Figure 4 NH4+-N/NO3--N between different tree species and soil layers
土层深度变化对7个树种NH4+-N/NO3--N影响各异,7个树种和撂荒地未表现出土壤层次差异(图4)。其中,各树种(除红椿和香樟外)NH4+-N/NO3--N随土层深度增加,比值呈“先上升后下降”的趋势,香樟则相反,红椿和撂荒地比值则随土层深度增加而升高。各土层土壤NH4+-N/NO3--N平均值分别为0.197、0.168和0.194。双因素方差分析表明(表1):树种对土壤NH4+-N/NO3--N影响极显著(P<0.001),土层及其交互作用对土壤 NH4+-N/NO3--N无显著影响(P>0.05)。
2.3 土壤无机氮含量与土壤理化性质的关系
相关分析表明:NH4+-N含量与土壤黏粒含量、NH4+-N/NO3--N呈显著正相关关系,与土壤砂粒含量、pH值呈显著负相关。NO3--N含量与土壤砂粒含量、无机氮含量、MBN、N和P含量呈显著正相关,与容重、NH4+-N/NO3--N、C/N和C/P值呈显著负相关。无机氮含量与N、P、MBN含量呈显著正相关,与容重、pH、C/N和C/P值呈显著负相关。NH4+-N/NO3--N与容重、土壤黏粒含量、C/P和C/N值呈显著正相关,与土壤砂粒含量、无机氮含量、MBN和MBC含量呈显著负相关(图5)。
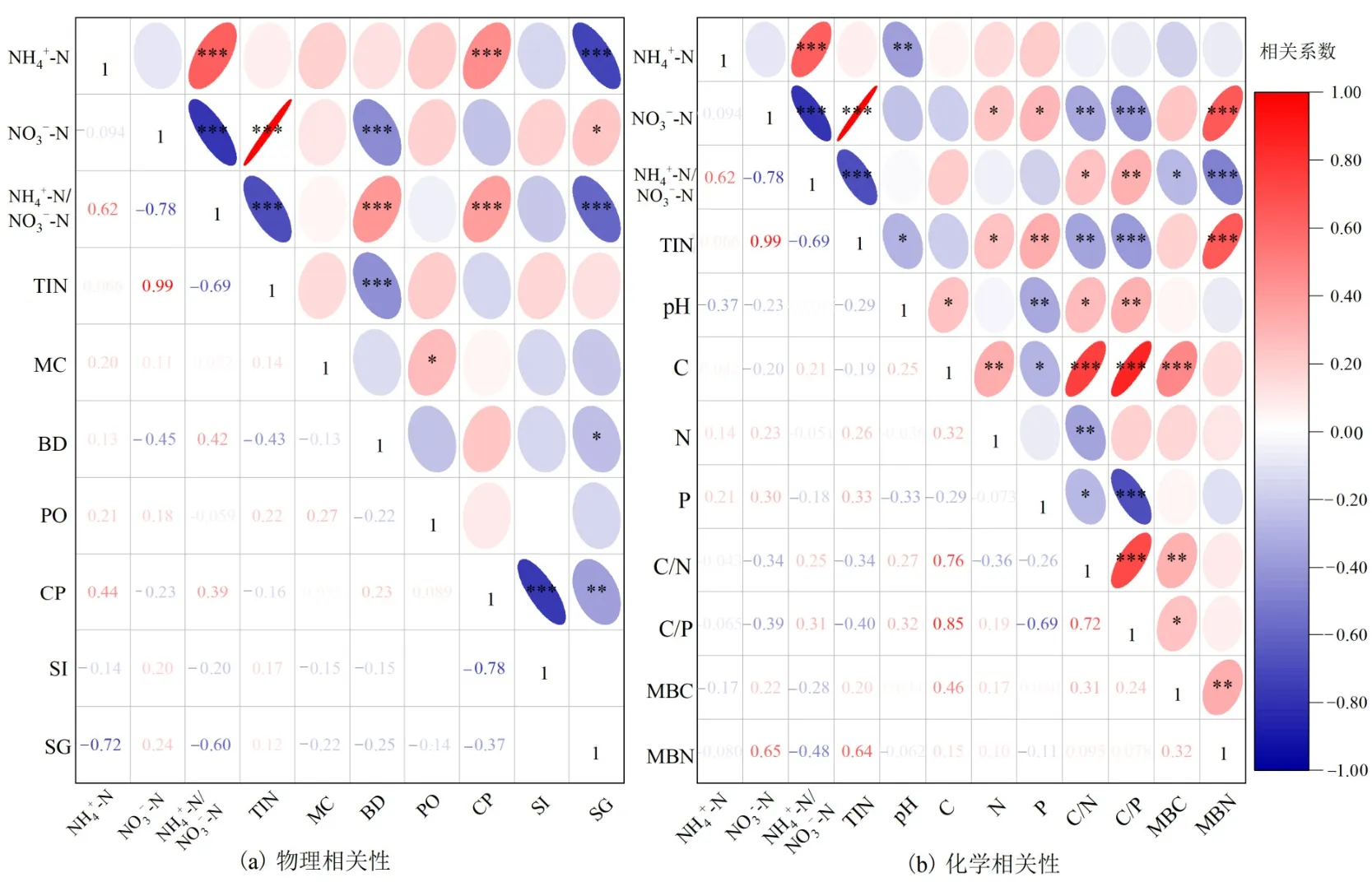
图5 铵态氮、硝态氮及其比值和无机氮和土壤理化参数的相关性Figure 5 Correlations between inoganic soil nitrogen and soil physio-chemical indexes of soil
为进一步验证和解释土壤特征因子与土壤无机态氮素之间的相互作用关系进行冗余分析(图6),结果表明,第一排序轴和第二排序轴分别能解释无机氮含量与土壤特征因子之间关系变异的70.94%和1.58%,Monte Carlo检验后,土壤MBN、土壤黏粒和 N含量与排序轴检验达极显著水平(P<0.01),其中土壤MBN含量与土壤NO3--N和无机氮含量呈显著正相关,与 NH4+-N和 NH4+-N/NO3--N呈显著负相关,土壤黏粒含量与NH4+-N和NH4+-N/NO3--N呈显著正相关,土壤 N与土壤NO3--N和无机氮含量呈显著正相关,这与Pearson相关性分析结果类似。
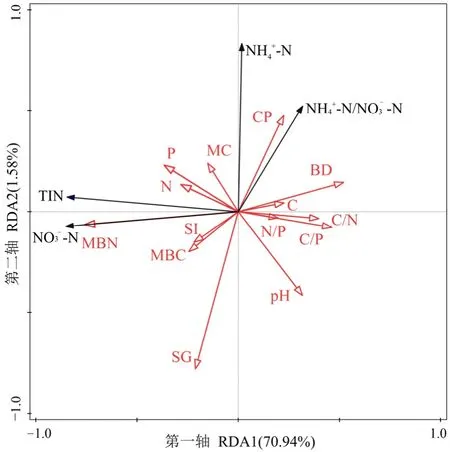
图6 铵态氮、硝态氮及其比值和无机氮含量与土壤理化指标的冗余分析Figure 6 Two-dimensional sequence diagram of redundancy analysis (RDA) between environmental factors and inoganic soil nitrogen contents
3 讨论
3.1 树种对NH4+-N、NO3--N和NH4+-N/NO3--N的影响
不同树种的土壤养分分布规律和空间变异不同,树种通过不同的养分吸收策略,对土壤无机氮的积累和利用不同(张彦东等,2003;戴全厚等,2008)。本研究表明,树种显著影响了土壤NH4+-N、NO3--N、NH4+-N/NO3--N和无机氮含量,树种可以结合自身养分需求来调节和改变对无机氮的吸收利用策略,这与前人研究结果一致,土壤中 NH4+和 NO3-的浓度和比例变化是树种生长的重要驱动力(Chen et al.,2021)。相比撂荒地,7个树种的NH4+-N含量均存在不同程度的下降(图1),本研究采样在7月中旬,此时样地林下环境高温高湿,林木生命活动旺盛,树种需要消耗大量无机氮来满足生长需求,土壤中NH4+-N含量的下降,表明他们均具有优先吸收利用NH4+的特性。研究表明,树种吸收利用土壤中NH4+耗能更少,故树种可通过硝化作用,利用NH4+-N来最大限度的保障生长所需养分,这可能是导致树种土壤NH4+-N含量下降的主要原因(邹婷婷等,2017)。7个树种的土壤NH4+-N 含量下降程度香樟>天竺桂>红椿>香椿>油樟>大叶樟>桤木(图1),并且香樟和天竺桂NH4+-N含量显著低于其他树种,表明香樟和天竺桂相比其他树种对 NH4+吸收速率更高。此外,本研究中 7个树种NH4+-N含量除油樟和香椿外均高于撂荒地(图2),而土壤中NO3-的来源主要是NH4+的氧化过程(Zhang et al.,2021),进一步表明树种通过NH4+氧化产生NO3-这一过程来保障生长必须养分,故土壤中NO3--N含量相较撂荒地出现不同程度的增加。同时,油樟和香樟NO3--N含量显著低于其他树种,油樟和香樟属于重要的植物精油提取源,叶片中富含丰富的次生代谢产物,树种生产这些物质需要消耗利用大量氮源,所以树种的养分分配向叶片倾斜所致(王米雪等,2020;肖祖飞等,2021)。本研究中各树种的土壤无机氮以NO3--N为主,这与Wang et al.(2011)研究结果相近,一般情况下天然森林中土壤无机氮多以NH4+-N为主,而由透气性良好的农耕土或易受干扰的土壤组成的人工次生林土壤,其无机氮多以NO3--N为主。本研究土壤来自农耕土,佐证了这一观点。树种受自身生理学属性和遗传特征的影响对土壤中无机氮的利用程度不同。大叶樟、天竺桂、红椿和桤木土壤无
机氮含量要显著高于撂荒地和其他树种(图 3),整体含量比撂荒地无机氮含量提高了 19.3%—27.6%。其中,桤木属于固氮树种,能通过共生根瘤菌获得额外氮源,故在土壤中无机氮含量较高。对于大叶樟、天竺桂和红椿,这表明他们有利于维持生态系统生产力,对土壤中氮素的利用程度较高,有益于自身生长。土壤NH4+-N/NO3--N是土壤铵挥发、淋溶和反硝化过程的间接体现,能一定程度反映不同树种在土壤中的氮循环(Cui et al.,2007)。本研究中天竺桂、红椿和大叶樟土壤NH4+-N/NO3--N显著低于其他树种和撂荒地(图4)。前期研究表明,土壤 NH4+-N/NO3--N与植物氮利用效率呈负相关关系(Xu et al.,2012),进一步说明大叶樟、天竺桂和红椿能较好地维持和利用土壤无机氮。
3.2 土层对NH4+-N、NO3--N和NH4+-N/NO3--N的影响
本研究7个树种土壤NH4+-N含量随土层深度增加的变化有一定差异(图 1),其中油樟、大叶樟和天竺桂随土层深度增加而含量下降,即具有表聚现象,这可能与植物凋落物归还主要发生在土壤表层,同时土壤表层存在有较多的植物根系和丰富的微生物群落有关(张仰等,2019)。此外,不同树种在凋落物数量和质量及根系分泌物等存在差异,土壤中根系活动深度也各不相同,从而影响土层的氧气有效性和有机质含量分布,造成不同树种对土壤养分利用策略的差异和选择吸收性(戴全厚等,2008;葛晓敏等,2019),也在一定程度上影响了土壤中NH4+在各土层的分布。桤木、香樟、香椿和红椿随土层深度增加,各土层NH4+-N含量变化趋势各异,可能与此有关,是树种生理属性和环境因素的综合作用结果。各树种土壤中NO3--N和无机氮含量在不同土层变化趋势相似(图2、图3),整体上随土层深度的增加含量下降。这与Menyailo et al.(2002)研究结果相近。通常随土层深度增加,微生物能分解矿化的底物也会下降,进而影响植物硝化作用与氮矿化速率,造成土壤中NO3--N和无机氮含量的降低(张仰等,2019;Peng et al.,2020)。与此同时,油樟和香樟无机氮和NO3--N含量在不同土层中的表现与其他树种不同,均为10—20 cm土层最高,这可能是因为它们凋落叶在归还土壤中对表层土壤微生物氮矿化产生了抑制作用所导致(Wang et al.,2011),影响了土壤中的硝化细菌与氨化细菌。各土层不同树种的NH4+-N/NO3--N,天竺桂和红椿均显著低于其他树种,而NH4+-N/NO3--N与植物氮吸收效率密切相关,NH4+-N/NO3--N越低植物氮吸收效率则越高(Xu et al.,2012),故天竺桂和红椿更有利于土壤无机氮的维持与利用。
方差分析表明,土层深度显著影响 NH4+-N、NO3--N和无机氮含量,对NH4+-N/NO3--N影响不显著(表 1)。造成这一结果的原因可能是NH4+-N/NO3--N受到植物遗传属性的影响有关(Xu et al.,2012),因为植物在生长中具有特定的氮吸收偏好。此外,树种和土层对NH4+-N和NO3--N含量均有显著影响,但其交互作用仅对NO3--N含量产生显著影响(表1)。这可能是因为树种对NH4+和NO3-的吸收能力差异导致,在土壤中NH4+的移动速度约是NO3-的10倍,植物根系周围会形成阳离子消耗区来捕获NH4+用于生成NO3-。随土层加深,树种效应呈现“倒金字塔”,根系和淋溶作用向下迁移和扩散减缓,更多的 NH4+集中于土壤表层,NO3-则随植物根系在土层中不断侵入(Jackson et al.,2008;Wang et al.,2011)。同时,方差分析结果表明树种和土层及其交互作用对无机氮有极显著影响,而NH4+-N/NO3--N仅受树种影响(表1),这侧面佐证了 NH4+-N/NO3--N受植物遗传属性影响,随时间推移,受环境因素影响较小。对于土壤无机氮,受到土壤微生物和植物根系活动空间和季节的影响,会在土壤中随时间产生强烈变化(Cui et al.,2017),故会受到土层深度影响。
3.3 土壤理化性质对 NH4+-N、NO3--N和NH4+-N/NO3--N的影响
相关性分析表明,14个土壤理化指标中,3个与土壤NO3--N相关,7个与土壤NH4+-N和无机氮有关(图6)。其中,NO3--N和无机氮含量与土壤密度和土壤黏粒呈显著负相关,土壤密度和黏粒大小表征着土壤孔隙的大小和土壤质地的紧密程度;土壤密度和黏粒含量越大则会导致土壤通气性的下降,直接影响土壤NO3-的淋失程度,土壤中NO3-在缺氧条件下极易挥发(Pang et al.,2011)。这与李聪等(2020)研究结果一致,均表明土壤密度与土壤无机氮存在显著负相关。同时,土壤 NO3--N和无机氮含量与土壤pH、C/N和C/P呈显著负相关,与土壤N、P和MBN显著正相关。土壤pH值影响植物对土壤中NH4+的有效利用程度,当植物根系吸收NH4+时,植物体内H+的流出会增加,为保证土壤电荷平衡,植物根际周围会加强对NO3-的吸附,造成土壤中NO3--N含量升高,最终影响土壤无机氮(Bauhus et al.,1998;张彦东等,2003)。这与朱红霞等(2010)研究结果一致,土壤pH与NO3--N和无机氮的空间分布格局具有较强的相关性。土壤中C、N和P元素均为植物生长发育必需元素,C/N和C/P均能改变土壤有机质的分解进程,其比值低,将有利于土壤中有机质的分解矿化,促进土壤无机氮的积累(Templera et al.,2003)。研究表明土壤中富含P元素能促进植物氮循环,因为土壤中P元素能促进微生物的生长和提高酶活性,最终影响土壤无机氮(Feng et al.,2019)。本研究中土壤无机氮含量与MBN呈显著正相关,这与Xu et al.(2017)研究结果一致。MBN是不稳定氮库的重要源(矿化)和汇(固定),土壤中MBN升高会使得土壤微生物新陈代谢速度加快,造成土壤微生物矿化加速,并释放出可溶性氮供植物利用(张文雯等,2019)。此外,MBN、N/P和C/P等对NH4+-N和NO3--N均有显著的消极影响,表明土壤NH4+-N和NO3--N含量受到土壤中不同种类和数量的微生物对C、N和P养分的需求差异和他们间生态化学计量比的综合作用影响。
4 结论
本研究通过同质园试验消除了立地条件和林龄的差异,研究了7个乡土树种对NH4+-N、NO3--N含量及其比值和无机氮含量的影响。树种和土层均能显著影响NH4+-N、NO3--N和无机氮,且树种和土层对NO3--N和无机氮产生了交互作用,而土壤NH4+-N/NO3--N则仅受树种效应的影响。7个树种和撂荒地土壤无机氮总体有表聚现象,土壤无机氮含量随土层深度增加而下降,这在桤木、大叶樟、香椿和红椿中表现尤为明显,表现出一定的森林土壤特征。相关性分析和冗余分析表明,树种主要通过影响土壤物理参数(机械组成、容重)和化学特性(pH、碳氮比、碳磷比和微生物生物量氮)来影响土壤NH4+-N、NO3--N和无机氮含量,其中土壤密度、pH、C/N和C/P与土壤无机氮含量呈显著负相关,N、P、MBN含量则与土壤无机氮含量呈显著正相关,土壤黏粒、全氮和微生物生物量氮是引起土壤无机氮含量变化的关键影响因子。就土壤无机氮而言,相比其他乡土树种和撂荒地,大叶樟,天竺桂和红椿更适合在四川盆地推广应用。
