林麝及其近缘物种编码区微卫星分布规律及功能分析
2022-12-19赵琪张琪李浩玲兰月鄢行安赵贵军戚文华
赵琪张琪李浩玲兰月鄢行安赵贵军*戚文华*
(1重庆三峡学院生物与食品工程学院,重庆 404100)(2四川大学生命科学学院,成都 610064)(3重庆市药物种植研究所,重庆 408435)
麝属动物(Moschusspp.)是鲸偶蹄目(Cetartiodactyla)反刍亚目(Ruminantia)麝科(Moschidae)中型哺乳动物(Montgelardet al.,1997)。麝属动物雌雄分化明显,始终保持较原始的特征,兼有原始型的鼷鹿科和进化型的鹿科的特征。麂属动物(Muntiacusspp.)属鹿科(Cervidae)麂亚科(Muntiacinae),主要分布于中国南部、南亚、东南亚(Ohtaishi and Gao,1990)。在我国,林麝(Moschus berezovskii)数量较多,分布范围广,主要分布于陕西、四川、青海、宁夏等地的针阔混交林、阔叶林或次生灌林;原麝(Moschus moschiferus)主要分布在黑龙江、吉林、内蒙古等地的针阔混交林(刘志霄和盛和林,2000)。最近,我国首次完成了林麝和原麝全基因组序列测序,并绘制了林麝和原麝全基因组图谱(Fanet al.,2018;Yiet al.,2020)。研究表明,我国林麝和原麝种群遗传多样性低,有待进一步加强种内基因交流,防止近亲繁 育(Fanet al.,2018;Yiet al.,2020)。小 麂(Muntiacus reevesi)在麂属中体型最小,是我国特有的物种,主要分布于中国的亚热带地区;赤麂(Muntiacus vaginalis)体型较大,也是我国重点保护野生动物,主要分布于中国东南、华南、西南等南部地区,在东南亚国家也有分布(Ohtaishi and Gao,1990)。这些麂属动物具有多变的染色体数目和核型,是由鹿科祖先染色体反复串联融合而导致,但其分子机制尚不清楚(Yinet al.,2021)。随着马鹿(Cervus elaphus)基因组测序的完成,Bana等(2018)从马鹿全基因组水平探索其与其他鹿科动物的系统进化关系。
微卫星(Microsatellite),又称简单重复序列(Simple sequences repeats,SSRs),广泛存在于真核生物中,也存在于原核生物和细菌的基因组中,多数分布于非编码区,少数分布于编码区(戚文华等,2013)。SSRs具有高重复性、高多态性,参与染色体组成、蛋白质功能、基因转录、表达与调控等过程。此外,SSRs与动物疾病的发生发展密切联系,如三核苷酸SSRs与人类疾病密切相关(王希恒等,2016)。麝科和鹿科动物具有重要的经济价值,目前关于林麝全基因组以及外显子区和基因间隔区SSR分布规律已有相关的研究(卢婷等,2017;Qiet al.,2020),原麝、小麂、赤麂和马鹿全基因组以及编码区SSR分布规律尚未有相关报道。因此,本文以林麝、原麝、小麂、赤麂和马鹿为代表物种,利用生物信息学方法提取其基因组中的编码区序列,从基因组水平分析编码区SSR分布规律,比较其编码区SSR分布规律的相似性和差异性,并对林麝及其近缘物种的CDS区SSR序列进行GO富集分析和KEGG通路分析,旨在从基因组水平探索林麝及其近缘物种的进化关系及进化特征,揭示富含微卫星序列的基因功能及其富集的信号通路,并探索含SSR序列的关键免疫基因与疾病的关联性。
1 研究方法
1.1 材料来源
林麝基因组序列和注释文件由本实验室提供,原麝、马鹿、赤麂、小麂基因组序列从Ensembl Genome Browser数据库(https://grch37.ensembl.org)下载,均以FASTA格式保存。使用Perl和R语言编程从林麝、原麝、小麂、赤麂和马鹿基因组中提取CDSs序列,以FASTA格式保存。
1.2 数据分析
1.2.1 微卫星识别与鉴定
利用MSDBv2.4(Microsatellite search and building database)(Duet al.,2013)和Krait(Duet al.,2017)软件对林麝、原麝、小麂、赤麂和马鹿编码区中完整型微卫星序列进行识别和鉴定,并分析其特征。搜索和统计标准为单核苷酸(Mononucleotide,Mono-)重复至少12次,二核苷酸(Dinucleotide,Di-)重复至少7次,三核苷酸(Trinucleotide,Tri-)重复至少5次,四核苷酸(Tetranucleotide,Tetra-)、五核苷酸(Pentanucleotide,Penta-)、六核苷酸(Hexanucleotide,Hexa-)重复至少4次;重复序列两端的侧翼序列为200 bp;因为1个氨基酸由3个碱基翻译而来,所以我们将三核苷酸和六核苷酸称为三倍体核苷酸,单核苷酸、二核苷酸、四核苷酸、五核苷酸称为非三倍体核苷酸。其他统计标准及SSRs序列统计术语参考相关报道(戚文华等,2013;蒋雪梅等,2015)。
1.2.2 GO功能富集分析
将林麝及其近缘物种编码区含有SSR的基因进行序列提取,保存为FASTA格式,并对这些序列进行同源性分析。使用TBtools(Chenet al.,2020)将含有SSR的基因与Swissprot蛋白质库进行比对,设置E-value<1E-5,其比对结果利用TBtools进行GO功能注释。按照分子功能(Molecular function,MF)、细胞组成(Cellular component,CC)和生物学过程(Biological process,BP)进行GO功能富集分析。
1.2.3 KEGG通路分析
以分配到KEGG通路所有哺乳动物基因为背景,使用KAAS(Yukiet al.,2007)进行KEGG注释,得到的ko文件再使用TBtools进行KEGG通路富集分析。将富集统计的P值进行Bonferroni校正,以矫正后P值为标准,当P<0.05时认定此KEGG通路为显著富集,并具有统计学意义。从而将含有SSR的基因与相关通路关联在一起,探讨含SSR的基因参与的相关生物功能过程。
2 结果
2.1 林麝及其近缘物种编码区基因组分析
在林麝编码区共计识别和鉴定24 352个基因,其中有1 696个基因包含完整型SSR序列,最多一个基因含有72个SSR位点,大多数基因包含1个SSR位点(1 327个,78.24%),较少基因包含2个及以上SSR位点(369个,21.76%)。在原麝编码区识别和鉴定的29 305个基因中,有2 389个基因包含完整型SSR序列,其中最多一个基因含有11个SSR位点,大多数基因包含1个SSR位点(1 924个,80.54%),较少基因包含2个及以上SSR位点(465个,19.46%)。在小麂编码区识别和鉴定的26 044个基因中,有1 916个基因包含完整型SSR序列,其中最多一个基因含有14个SSR位点,大多数基因包含1个SSR位点(1 510个,78.81%),较少基因包含2个及以上SSR位点(406个,21.19%)。在赤麂编码区识别和鉴定的25 737个基因中,有1 924个基因包含完整型SSR序列,其中最多一个基因含有26个SSR位点,大多数基因包含1个SSR位点(1 545个,80.30%),较少基因包含2个及以上SSR位点(379个,19.70%)。在马鹿编码区共计识别和鉴定的28 103个基因中,有3 005个基因包含完整型SSR序列,其中最多一个基因含有9个SSR位点,大多数基因包含1个SSR位点(2 478个,82.46%),较少基因包含2个及以上SSR位点(527个,17.54%)(表1)。林麝编码区序列数量、SSR序列的数量和长度、丰度和密度与其近缘物种有一定差异(P<0.05,ttest),而五者GC含量和SSR序列占编码区基因组长度的比例基本一致。
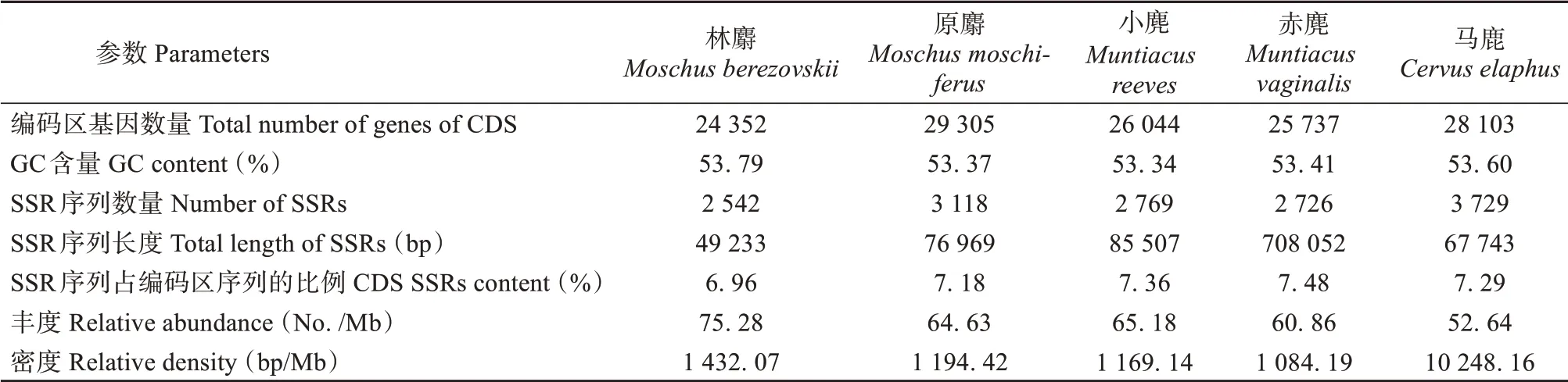
表1 林麝及其近缘物种编码区基因概况Table 1 Overview of the CDS of Moschus berezovskii and its closely related species
2.2 林麝及其近缘物种蛋白质编码区SSR分布规律
林麝和原麝编码区完整型SSR丰度分布模式基本一致,均是三核苷酸SSR最多(93.47%vs.91.34%),其 次 是 六 核 苷 酸SSR(3.38%vs.3.53%),较少的均是单核苷酸(1.02%vs.1.80%)、二核苷酸(0.63%vs.1.96%)、四核苷酸(0.79%vs.0.96%)、五核苷酸(0.71%vs.0.41%)。小麂、赤麂和马鹿编码区完整型SSR丰度分布模式基本一致,均是三核苷酸SSR最多(63.46%vs.62.14%vs.85.55%),其次是二核苷酸(19.25%vs.18.82%vs.3.03%),较少的均是单核苷酸(11.48%vs.12.84%vs.6.60%)、四核苷酸(3.28%vs.3.30%vs.1.96%)、五核苷酸(0.54%vs.0.81%vs.0.37%)、六核苷酸(1.99%vs.2.09%vs.2.09%)。由此可见,林麝、原麝、小麂、赤麂和马鹿的三倍体核苷酸SSRs(三核苷酸和六核苷酸SSRs)占比最高,分别为96.85%、94.87%、65.44%、64.23%和88.04%,非三倍体核苷酸SSRs(单核苷酸、二核苷酸、四核苷酸和五核苷酸SSRs)占比分别为3.15%、5.13%、34.56%、5.77%和11.96%(图1)。

图1 林麝及其近缘物种蛋白质编码区不同重复拷贝类别SSRs丰度比较Fig.1 Comparison of SSR frequency of different repeat types in the CDS regions of the Moschus berezovskii and its closely related species
在林麝及其近缘物种CDS区,不同重复类型SSR序列GC含量的分布模式有所差别。林麝的GC含量的分布模式为三核苷酸SSRs(73.45%)>五核苷酸SSRs(72.39%)>六核苷酸SSRs(69.47%)>四核苷酸SSRs(72.31%)>二核苷酸SSRs(51.83%)>单核苷酸SSRs(38.95%)。原麝的GC含量的分布模式为六核苷酸SSRs(81.53%)>三核苷酸SSRs(75.97%)>四核苷酸SSRs(72.00%)>五核苷酸SSRs(63.70%)>二核苷酸SSRs(52.21%)>单核苷酸SSRs(6.66%)。小麂与赤麂的GC含量的分布模式相同,均为六核苷酸SSRs(79.50%vs.76.61%)>三核苷酸SSRs(73.44%vs.72.51%)>五 核 苷 酸SSRs(53.11%vs.54.95%)>二核苷酸SSRs(48.59%vs.48.72%)>四核苷酸SSRs(47.18%vs.46.10%)>单核苷酸SSRs(26.41%vs.33.76%)。马鹿的GC含量的分布模式为六核苷酸SSRs(77.54%)>五核苷酸SSRs(74.24%)>三核苷酸SSRs(72.94%)>四核 苷 酸SSRs(63.06%)>二 核 苷 酸SSRs(52.66%)>单核苷酸SSRs(41.50%)。由此可知,在林麝及其近缘物种的CDS区,SSR序列的丰度在三核苷酸SSR序列趋于最高,林麝的GC含量在三核苷酸SSR序列趋于最高,而原麝、小麂、赤麂和马鹿的GC含量均在六核苷酸SSR序列趋于最高。
2.3 林麝及其近缘物种编码区SSRs重复拷贝类别比较
在林麝及其近缘物种CDS区,单核苷酸至六核苷酸SSR序列重复拷贝类别的丰度有明显差异(图2,图3)。在单核苷酸SSR重复拷贝类别中,A重复类别的丰度稍高于C重复类别的丰度;在二核苷酸SSR中,AC重复类别的丰度最高,林麝、原麝和马鹿AT的重复类别的丰度最少,而小麂和赤麂CG重复类别的丰度最少。在三核苷酸SSR中,CCG重复类别的丰度最高,AAC重复类别的丰度最少;在四核苷酸SSR中,林麝、原麝和马鹿丰度差异不大,赤麂和小麂CCCG重复类别的丰度为0(图2,图3)。总体上,在CDS区,林麝大部分重复拷贝类别的丰度高于其近缘物种,并且林麝和原麝的重复类别丰度更为相似,小麂、赤麂、马鹿的重复类别丰度更为相似。

图2 林麝和原麝蛋白质编码区不同重复拷贝类别SSRs丰度比较Fig.2 Comparison of SSR frequency of different repeat category in the CDS regions of the Moschus berezovskii and Moschus moschiferus

图3 马鹿、赤麂和小麂蛋白质编码区不同重复拷贝类别SSRs丰度比较Fig.3 Comparison of SSR frequency of different repeat category in the CDS regions of the Cervus elaphus,Muntiacus vaginalis and Muntiacus reeves
2.4 林麝及其近缘物种编码区SSR序列的功能富集分析
2.4.1 GO注释和富集分析
GO富集分析表明,林麝、原麝、小麂、赤麂和马鹿大部分GO功能主要集中在生物学过程,分别 为85.94%、83.52%、82.52%、83.91%和82.32%。具体如下:林麝GO功能富集结果共634条,其中涉及分子功能的有48条(占7.58%),主要涉及细胞核不同结构的结合、DNA结合和DNA结合转录激活因子活性等;涉及细胞组成的有42条(占6.64%),主要有具膜细胞器、染色质和染色体、转录调节复合物等;涉及生物学过程的有544条(占85.94%),主要包括生长发育、各种细胞的分化、代谢过程的调控和正负反馈调节等。原麝GO功能富集结果共634条,其中涉及分子功能的有170条(占9.87%),涉及细胞组成的有114条(占6.62%),涉及生物学过程的有544条(占83.52%)。小麂GO功能富集结果共727条,其中涉及分子功能的有64条(占8.80%),涉及细胞组成的有53条(占7.29%),涉及生物学过程的有610条(占83.91%)。赤麂GO功能富集结果共758条,其中涉及分子功能的有73条(占9.63%),涉及细胞组成的有61条(占8.05%),涉及生物学过程的有624条(占82.32%)。马鹿GO功能富集结果共2 054条,其中涉及分子功能的有189条(占9.20%),涉及细胞组成的有170条(占8.28%),涉及生物学过程的有1 695条(占82.52%)。各类编码区SSR序列的主要功能与林麝的相似,GO条目的显著性有微小的差别。
通过林麝及其近缘物种蛋白质编码区SSR序列进行功能分析表明,在分子功能方面,林麝与原麝最显著富集的转录调控区序列特异性DNA结合(Transcription regulatory region sequence-specific DNA binding)、顺式调控区序列特异性DNA结合(Cis-regulatory region sequence-specific DNA binding)、调控区核酸结合(Regulatory region nucleic acid binding)等30个GO功能条目具有高度一致性,而这些功能条目在小麂、赤麂和马鹿中仅有转录调控区序列特异性DNA结合、调控区核酸结合、分子功能监管机制(Molecular function regulator)不显著;与林麝、原麝相比较,小麂、赤麂和马鹿在这30个最显著富集GO功能条目中更具相似性。
在细胞组成方面,林麝与原麝最显著富集的30个GO功能条目中仅在Viral outer capsid(GO:0039624)不显著,其余29个显著富集的GO功能条目完全一致;与林麝、原麝相比较,小麂、赤麂和马鹿在核染色体(Nuclear chromosome)、生长锥(Growth cone)、树突细胞质(Dendrite cytoplasm)、树突(Dendritic spine)、多线染色体(Polytene chromosome)和神经元棘(Neuron spine)GO功能条目中不显著,其余23个显著富集的GO功能条目高度一致。
在生物学过程中,林麝与原麝最显著富集的氮化合物代谢过程的正调控(Positive regulation of nitrogen compound metabolic process)、RNA生 物合成过程的正调控(Positive regulation of RNA biosynthetic process)、RNA生物合成过程的负调控(Negative regulation of RNA biosynthetic process)等30个GO功能条目在原麝中均显著富集;与林麝、原麝相比较,小麂、赤麂和马鹿在这30个显著富集的GO功能条目高度一致,但有细微差别(表2)。由此可见,林麝与原麝更具亲缘关系,小麂、赤麂和马鹿亲缘关系更近,这与生物学分类相符。

表2 林麝及其近缘物种蛋白质编码区SSR序列的生物学过程的功能富集(top 30)Table 2 The most significantly enriched biological process GO terms of coding SSRs of Moschus berezovskii and its closely related species(top 30)
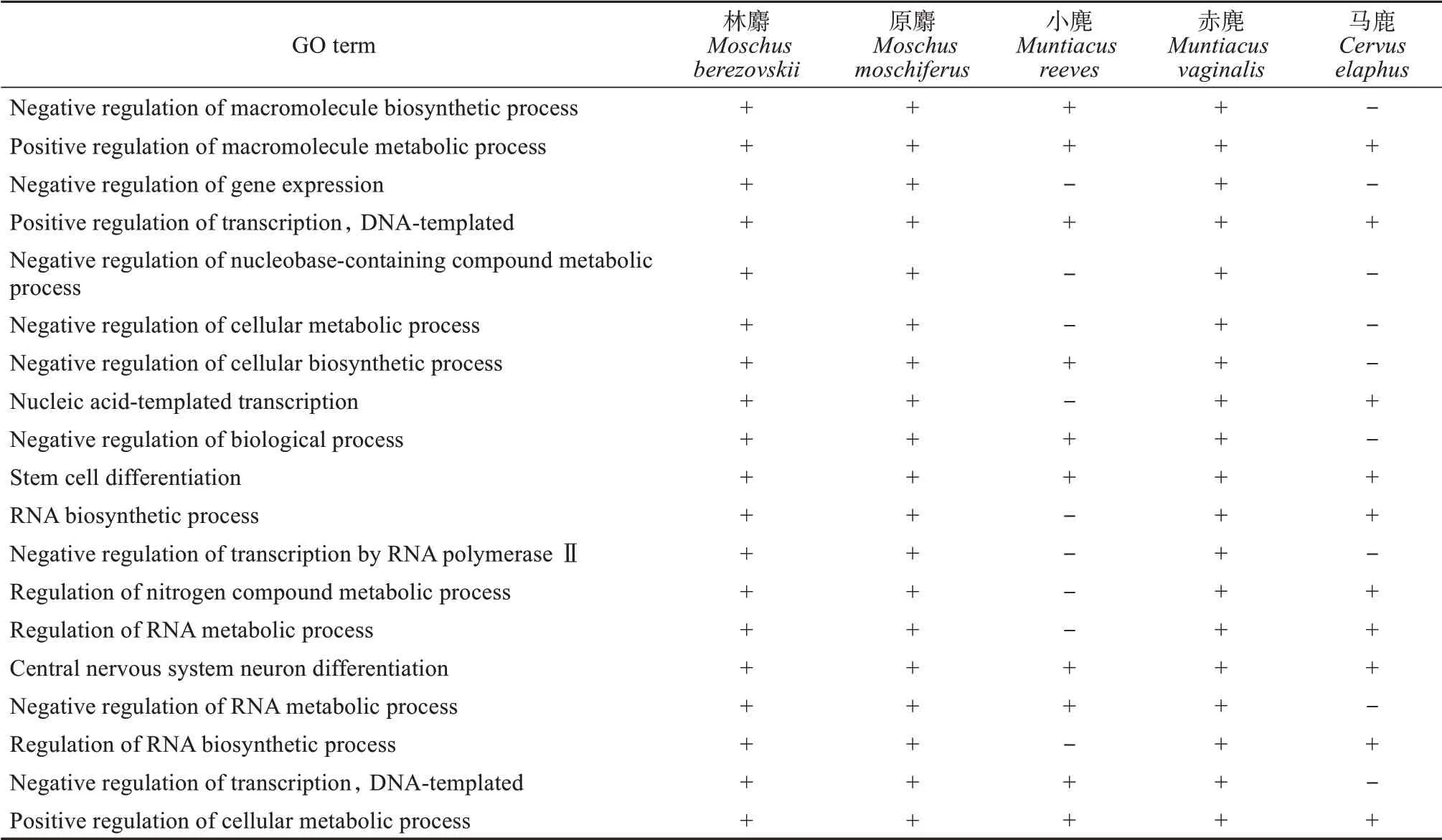
续表2 Continued from table 2
2.4.2 KEGG富集分析
KEGG富集分析表明,林麝、原麝、小麂、赤麂和马鹿编码区SSR序列富集到43个KEGG通路上,这些通路被分成5大类:遗传信息调控(Genetic information processing)、环境信息调控(Environmental information processing)、机体系统(Organismal systems)、新陈代谢(Metabolism)和疾病(Diseases)。在遗传信息调控通路中,这5种动物编码区SSR序列共同显著富集的通路是遗传信息调控蛋白家族(Protein families:genetic information processing)、转录因子(Transcription factors)、染色体及相关蛋白(Chromosome and associated proteins)、剪接体(Spliceosome)和转录机制(Transcription machinery)。在环境信息通路中,这5种动物编码区SSR序列共同显著富集的通路仅有Notch信号通路(Notch signaling pathway)。在新陈代谢中,这5种动物编码区SSR序列没有共同显著富集的通路,林麝、原麝、小麂和马鹿共同显著富集的通路是促性腺激素通路(GnRH secretion),而在赤麂中不显著;在林麝、原麝、赤麂和马鹿编码区SSR序列共同显著富集的通路是赖氨酸降解通路(Lysine degradation),而在小麂中不显著;在原麝和小麂编码区SSR序列中共同显著富集的通路是甲状旁腺激素的合成、分泌和作用通路(Parathyroid hormone synthesis,secretion and action),但在林麝、赤麂和马鹿中不显著;在小麂和马鹿编码区SSR序列显著富集的通路是皮质醇的合成和分泌通路(Cortisol synthesis and secretion),在林麝、原麝和赤麂中不显著;在小麂和马鹿编码区SSR序列显著富集的通路是胰岛素分泌通路(Insulin secretion),但在林麝、原麝和赤麂中不显著。在机体系统通路中,原麝和马鹿编码区SSR序列显著富集的通路是细胞粘连通路(Cell adherens junction),而在林麝、小麂和赤麂中不显著。在疾病通路中,这5种动物编码区SSR序列共同显著富集的通路是成体糖尿病(Maturity onset diabetes of the young)(表3)。由此可见,林麝及其近缘物种蛋白质编码区SSR序列具有共同显著富集的KEGG通路,通过进一步比较发现,林麝与原麝的编码区SSR序列共有显著富集的KEGG通路较多,小麂和赤麂的也较多,但马鹿的编码区SSR序列显著富集的KEGG通路与其他4种动物相比差别较大,这与生物学分类相符合。

表3 林麝及其近缘物种编码区SSR序列的KEGG通路富集(top 30)Table 3 The most significantly enriched KEGG pathway of coding SSRs of Moschus berezovskii and its closely related species(top 30)
2.5 含SSR关键免疫基因分析
通过对林麝编码区含SSR关键免疫基因及其相关联的KEGG通路进行分析,发现有10个含微卫星序列的关键免疫基因对应的KEGG通路与疾病密切相关。其中CEBPB、ZEB1和Bcl2基因参与癌症中的转录失调通路(Transcriptional misregulation in cancer,map05202),ZEB1、Bcl2、HDAC5和Notch4基因参与癌症中的miRNA通路(MicroRNAs in cancer,map05206),Bcl2、JAG2和NOTCH4基因参与癌症通路(Pathways in cancer,map05200)。在这10个基因中,Bcl2基因关联的疾病相关通路较多,包括多重神经退行性疾病通路(Map05022)、癌症中的转录误调节(Map05202)、结直肠癌(Map05210)、胰腺癌(Map05212)、小细胞肺癌(Map05222)、前 列 腺 癌(Map05215)、胃 癌(Map05226)和慢性骨髓白血病(Map05220)等通路(表4)。由分析推测,免疫基因中SSR的重复类型和重复次数与免疫疾病密切相关,这可能由于SSR重复次数的增加或缩减导致原表达蛋白质的功能丧失,从而产生相应疾病。
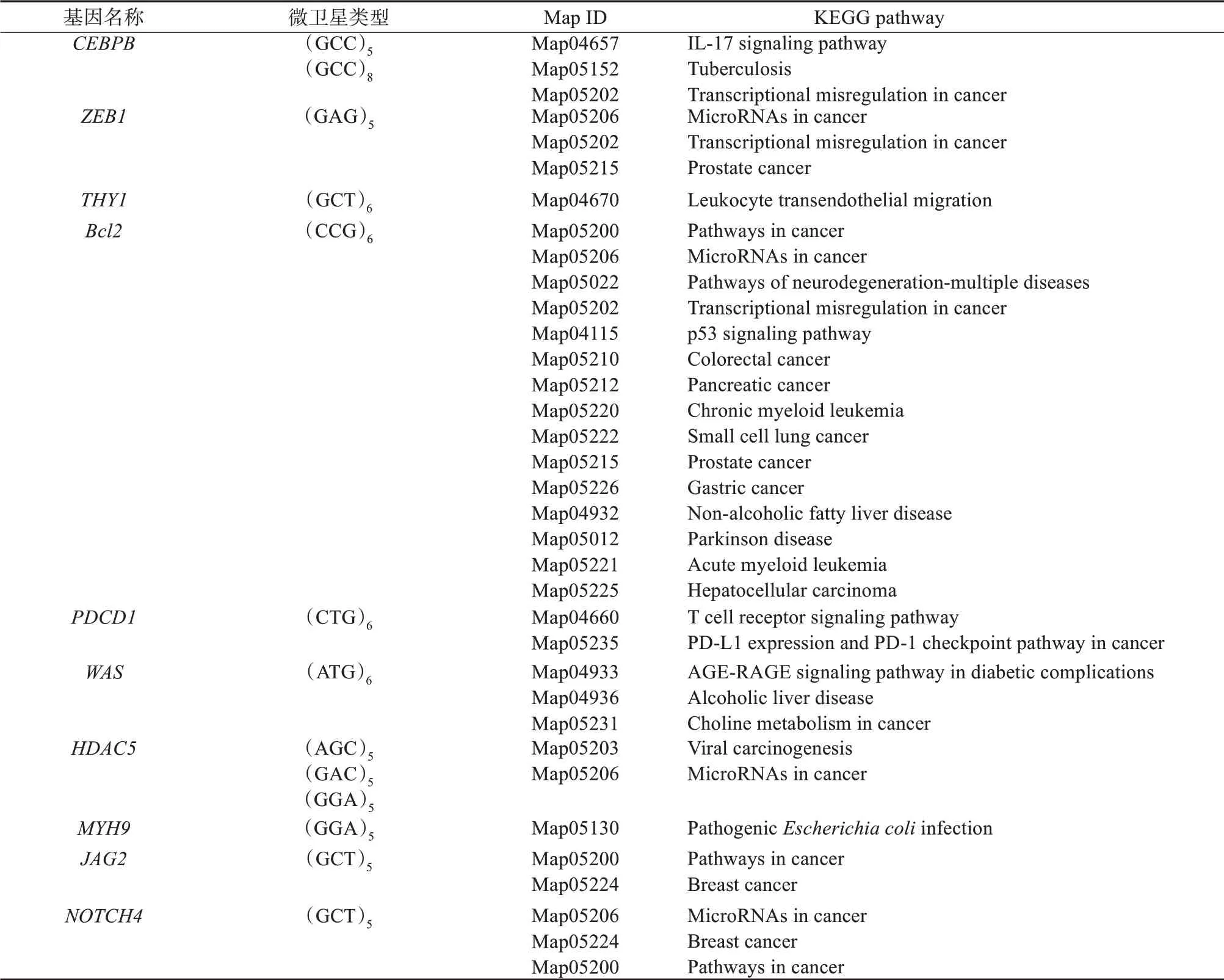
表4 林麝关键免疫基因所含SSR类型及其KEGG通路Table 4 SSR types of key immune genes in Moschus berezovskii and their KEGG pathways
3 讨论
本研究利用生物信息学方法测定和统计了林麝、原麝、小麂、赤麂和马鹿编码区SSR序列的特征,并对CDS区的SSR序列的功能进行了系统分析。林麝、原麝、马鹿、小麂和赤麂编码区含SSR序列的基因数量分别是2 588个、5 772个、3 821个、9 397个和6 590个,其SSR序列所占的比例基本一致,分别为0.15%、0.12%、0.10%、0.12%和0.11%。林麝、原麝、小麂、赤麂和马鹿编码区均以三倍体核苷酸SSRs最多,分别为96.85%、94.87%、65.44%、64.23%和88.04%,这与大熊猫(Ailuropoda melanoleuca)、北极熊(Ursus maritimus)(李午佼等,2014)、牛、绵羊(戚文华等,2013)、肩突硬蜱(Ixodes scapularis)(汪自立等,2013)、人类(Subramanianet al.,2003;O’Dushlaineet al.,2005)等物种编码区中SSR序列研究基本一致。林麝编码区SSRs的分布特征与卢婷等(2017)的研究基本一致。在三倍体核苷酸SSRs中,最多的是三核苷酸SSRs,重复拷贝类别多为AAC、ACC、ACG、AGC、AGG和CCG等,有利于RNA的转录和翻译,可能与防止移码突变的发生有关;六核苷酸SSRs数量较少,可能是由于微卫星长度越长越不稳定,容易发生变异,承受较大的选择压力;而非三倍体核苷酸(即单核苷酸、二核苷酸、四核苷酸、五核苷酸)SSRs极少,非三倍体核苷酸SSRs可能受到强烈的选择压力,与已有研究认为的编码区非三倍体核苷酸SSRs对移码突变的选择压力影响较大相符(Metzgar and Wills,2000)。在CDS区,林麝大部分重复拷贝类别的丰度高于其近缘物种,并且林麝和原麝的重复类别丰度更为相似,小麂、赤麂和马鹿的重复类别丰度更为相似,由此可见林麝和原麝同属麝科,小麂、赤麂和马鹿同属鹿科,具有一定的亲缘性,但是不同物种在长期环境变化的适应中,遗传物质也发生了演化,逐渐产生差异。
随着微卫星研究的深入,发现微卫星序列与蛋白质的表达、功能以及疾病的发生有一定的关联。微卫星可作为染色质折叠的密码。由于核苷酸的差异,不同DNA序列形成的二级结构形状会有差异,一定条件下的GC/CG与甲基化的胞嘧啶及二核苷酸重复的CA/GT易形成Z-DNA结构,执行重要的功能,如结构基因的启动子和重组热点均富含ZDNA结构(杨述林等,2003)。为研究林麝、原麝、小麂、赤麂和马鹿5个物种的编码区SSR序列的功能相似性,对其进行了GO功能富集分析和KEGG富集分析,结果表明编码区SSR序列在林麝及其近缘物种遗传效应上均具有重要的作用:在GO功能中显著富集的均与核系统、遗传物质的成分与进程、机体系统的结构与功能密切相关;KEGG通路富集分析表明,在遗传信息调控通路得到的通路共同显著富集的最多,有遗传信息调控蛋白家族、转录因子、染色体及相关蛋白、剪接体和转录机制,发现编码区SSR序列在遗传信息调控方面发挥重要的作用,可能是由于微卫星序列可以产生多样化的蛋白质功能,影响相应蛋白质的绑定性能(Kashiet al.,1997;Li,2004),进而影响其下游基因的表达,形成可调的功能变化和环境适应。
有研究表明,SSRs重复次数的增加或减少直接影响相关基因的表达,如CDS区域失活修复(MMR)基因(Duval and Hamelin,2002;Vassilevaet al.,2002)、转录调控基因(Duvalet al.,1999)、细胞凋亡基因(Schwartzet al.,1999)。在人类基因中,三核苷酸SSRs重复次数的增加与几种神经系统疾病有关,例如脆性X综合征(Jin and Warren,2000)、肌强直性营养不良(Timchenkoet al.,2001)、亨廷顿氏病和几种共济失调(Sermonet al.,2001)。蛋白质翻译过程中每3个核苷酸形成1个三联体密码,当编码区SSR重复序列增加,会使其基因功能丧失,从而导致产生相关疾病。通过对林麝编码区含SSR关键免疫基因及其相关联的KEGG通路进行分析,发现有10个含SSR的关键免疫基因对应的KEGG通路与疾病密切相关。其中Bcl2基因关联的疾病相关通路较多,包括多重神经退行性疾病通路(Map05022)、癌症中的转录误调节(Map05202)、结直肠癌(Map05210)、胰腺癌(Map05212)、小细胞肺癌(Map05222)、前列腺癌(Map05215)、胃癌(Map05226)和慢性骨髓白血病(Map05220)等。此外,微卫星与端粒和着丝粒相关(丁戈等,2008)。真核生物染色体端粒为重复序列,保护染色体末端。目前关于微卫星功能的研究比较浅显,没有深入研究其中的分子机理,应采用多学科交叉协作,结合遗传学、生物化学、分子生物学等学科知识,融会贯通深入研究其中机理。
致谢:感谢中国科学院成都生物研究所严超超助理研究员提供的帮助。
