施氮形态与分株损伤对入侵植物香菇草生长及克隆整合的影响
2022-12-19杨丽娟蔡竟芳穆亚楠沈一娈阿斯哈李红丽
孙 凯,杨丽娟,蔡竟芳,穆亚楠,程 蕊,沈一娈,阿斯哈,李红丽
(北京林业大学生态与自然保护学院,北京林业大学黄河流域生态保护国家林业和草原局重点实验室,北京 100083)
植物入侵对全球的生态环境造成了严重威胁,严重危害了入侵地生态系统的稳定及经济的发展[1-2]。研究发现,许多恶性入侵杂草为克隆植物,而克隆整合作用是体现外来植物生长和入侵能力的重要性状[3-4]。克隆整合是指克隆植物相连分株间能通过间隔子(匍匐茎和根状茎等)共享水分、养分、碳水化合物等资源[5-8]。此外,研究表明,异质环境可以提升克隆植物克隆整合所带来的好处,从而促进克隆植物的生长繁殖[9]。由于施肥、干扰和土壤性质的影响,植物生长所需的土壤养分在生境中的分布往往是异质性的[10]。因此,探究异质环境下入侵植物的克隆整合机制,对防控入侵性克隆植物的入侵尤为重要。
近几十年来,由于人类活动(如化石燃料燃烧、农业施肥),生态系统土壤的氮含量不断增加,显著影响植物的生长和功能[11-13],并且氮水平在区域间具有异质性。研究表明,土壤氮含量的异质性越强,克隆整合对克隆植物生长的积极作用越显著[14-15],但关于氮环境对入侵性克隆植物影响的研究仅基于单一形态氮的供应[9,14-15]。事实上不同形态的氮对植物的生长也有着重要影响。例如,施用一定量的硝态氮可明显促进植物侧根的生长,而过多的铵态氮则会导致根系变短,侧根减少以及根活力下降[16-17]。再如,外用硝态氮较铵态氮更能促进植物光合作用,但是铵态氮更有利于叶片中淀粉的积累[18]。最近一项研究表明,不同入侵植物对不同形态氮偏好不同,生境中氮素形态显著影响其生长及入侵[19]。因此,在探索植物生长对模拟氮沉降的响应时,应该考虑不同氮形态的潜在影响。
此外,植物在生活史中会受到非生物因素和生物因素的双重影响,其中生物因素中,植物经常受到由动物采食造成的分株损伤的影响[9,20]。研究表明分株损伤也显著影响克隆植物的生长和克隆整合[21-22]。一方面,分株损伤可以作为信号在克隆分株之间传输,以便正常分株可以提前应对损伤,从而提升总体的适应力[23],另一方面,分株损伤也可能通过克隆整合传播病原菌等导致植株死亡[24]。然而,关于氮沉降与分株损伤对克隆植物生长及其克隆整合的相互作用影响,目前所知较少。
香菇草Hydrocotylevulgaris是伞形科多年生草本克隆植物,通常由半湿润地带向湿润地带扩散[1,25]。每一个茎节都具有长出横向匍匐茎,产生新的叶柄和不定根的无性系分株的能力。虽然香菇草可以生长出可供有性生殖的花,但其主要是通过茎节繁殖出大量的克隆种群,并迅速扩散[1]。香菇草有较高的表型可塑性、较快的克隆繁殖速度以及广泛的耐受性,能够在环境中占据更宽的生态幅,是一种典型的入侵性克隆植物[9]。
为了探讨不同氮形态、损伤对入侵性的克隆植物生长以及克隆整合的综合作用机制,基于以上理论,我们选择了入侵性的克隆植物香菇草为研究对象,通过控制香菇草分株间连接或者切断,3种氮形态添加处理,以及分株是否损伤设计了三因素控制试验。本试验主要探究施氮形态、损伤、克隆整合三者对香菇草生长生理指标的独立及交互作用。进而为揭示侵入性克隆植物香菇草的入侵机制提供理论支持。
1 材料与方法
1.1 试验材料
本试验中采用的香菇草为浙江省杭州市西溪湿地的野生自然种群,于2014年11月中旬采集后放置于北京林大林业科技股份有限公司的温室(40°40′N,116°20′E)中进行培养。2016年4月12日,首先将试验中需要用到的香菇草置于试验容器(长50 cm,宽50 cm,高20 cm)中进行大量栽种繁殖。将草炭∶蛭石∶石英砂以1∶1∶1比例混合置于试验容器内作为培养基质,并在容器底部放置陶粒,以防止基质流失,培养周期约3个月。2016年7月19日,选取长势相似的香菇草片段,每个片段6个节点。将同一片段各3个节点栽种于两个同样大小的花盆(内径17 cm,外径19 cm,高13 cm)中,所用基质与前述培养基质相同,花盆两两成对摆放。所有容器都放置在开阔的场地,光照充足,温室内平均温度29.1℃,平均相对湿度81.6%。试验期间定期适量浇水,防止植物萎蔫。在移栽后7 d内,死去的植株均被替换。所有容器随机摆放,每月随机交换摆放位置。
1.2 试验设计
如图1所示,该试验采用三因素设计,包括3种氮形态添加(铵态氮、硝态氮、有机氮)和对照处理(去离子水),交叉2种叶片损伤处理(有无损伤)与2种克隆整合处理(连接或断开)。共进行16种处理,每种处理6个重复。
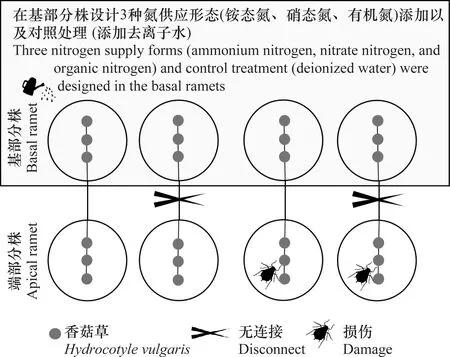
图1 试验设计Fig.1 Experimental design
每个香菇草克隆整合片段有6个节点,克隆整合处理分为连接或切断,即在整个克隆片段的中间部分进行连接或断开处理,其中年老一边的分株作为基部分株,另一边(年轻分株)作为端部分株。在香菇草基部分株设置3种施氮形态添加和对照处理,具体包括:CK(对照组,添加去离子水)、铵态氮[硫酸铵(NH4)2SO4]、硝态氮(硝酸钠NaNO3)、有机氮(Gly甘氨酸C2H5NO2),添加氮含量均为15 g/m2/年。试验中的氮素总量依据香菇草生存区域的大气氮湿沉降量和降水情况进行设置[26]。将各种形式氮养分溶于去离子水中,并分别定容至100 mL,均匀喷洒在土壤表面,对照处理喷洒100 mL去离子水。
试验开始时,在各香菇草端部分株的叶片顶端上投放6~8头桃蚜Myzuspersicae作为损伤处理,并定期检查以保持数量恒定,同时在有无蚜虫处理的一端均摆放120目虫网,以防止蚜虫扩散。桃蚜收集于北京林大林业科技股份有限公司的温室。以往的研究发现桃蚜可以对香菇草造成一定程度的损伤[9]。此外,试验第45 天时,使用直径6 mm的打孔器对叶片进行机械损伤处理,处理面积约占叶片总面积的30%,以进一步增加损伤对香菇草的刺激。
1.3 数据收集
试验过程中,于2016年9月29日上午9:00-12:00,使用LI-6400(Li-Cor,Lincoln,NE)光合仪,随机在每盆样本中各选择一片大小相似的叶片测定样本的净光合速率,测定期间天气晴朗,使用LED 2×3型红蓝光源叶室,叶片温度约为23℃,CO2浓度约为420 μmol/L,光合有效辐射(PAR)约为1 400 μmol/(m2·s)。试验收获时,缓慢倒置试验容器,将香菇草植株完整取出,并将其根部基质冲洗干净。将香菇草分为根、茎、叶三部分,香菇草的叶面积使用扫描仪和ImageJ (National Institutes of Health)软件计算获得,之后放入信封,置于70℃烘箱内烘干72 h至恒重,然后测定根、茎、叶生物量,并且以此计算总生物量。生物量测定完毕之后,先对植物进行H2SO4-H2O2消煮,然后使用连续流动分析仪(SEAL AA3,SEAL,Germany)测定根、茎、叶的氮含量。
1.4 数据分析
所有数据分析均在SPSS 20.0(IBM,Chicago,Illinois,USA)软件中进行。使用三因素方差分析(Three-way ANOVA),检验施氮形态、克隆整合、损伤处理及三者交互作用对香菇草总生物量、根生物量、茎生物量、叶生物量、叶片面积、净光合速率、总氮、根氮、茎氮及叶氮含量的影响。使用最小显著差异法(LSD)进行事后两两比较以分析各处理间香菇草生长生理性状的差异。方差分析之前,检验数据的正态分布和方差齐性,对不满足要求的数据进行对数或开平方根转换。最后用SigmaPlot 12.0(Systat Software,Chicago,IL,USA)绘图。
2 结果与分析
2.1 施氮形态、克隆整合和损伤处理对香菇草整个克隆片段以及前后端分株生物量的影响
克隆整合(连接或者切断)显著影响香菇草整个克隆片段、基部与端部分株的总生物量、根生物量、茎生物量以及叶生物量(P<0.05)(表1)。除添加有机氮且损伤时,连接处理的香菇草整个克隆片段的生物量指标低于切断处理,连接处理的香菇草整个克隆片段的其他生物量指标均高于切断处理(图2a~d)。不同施氮形态显著影响香菇草整个克隆片段的根生物量与叶生物量(P<0.05),也显著影响基部分株的总生物量、根生物量以及叶生物量(P<0.05),但是对端部分株的生物量指标没有显著影响(P>0.05)(表1)。相对于对照而言,添加3种氮形态均提高了香菇草整个克隆片段与基部分株的生物量指标,且在硝态氮添加时生物量达到最高,有机氮添加对香菇草生物量的提升最少,特别是在分株切断且无损伤或连接且损伤时,添加有机氮反而略微降低了香菇草整个克隆片段的生物量(图2)。值得注意的是损伤及各因素的交互作用对香菇草整个克隆片段、基部和端部分株生物量指标均没有显著影响(P>0.05)(表1)。此外,尽管在不同氮形态或损伤处理下,香菇草分株切断或者损伤时的生物量在数值上有所不同,但是基于事后多重比较的结果,氮形态与损伤并没有显著改变克隆整合对香菇草整个克隆片段总生物量的影响(图2a)。

表1 氮供应形态、克隆整合及损伤处理对香菇草整个克隆片段、基部与端部分株生物量影响的方差分析1)Table 1 ANOVA for effects of nitrogen supply form,clonal integration and damage on the biomass of total clonal fragment,basal and apical ramets of Hydrocotyle vulgaris
2.2 施氮形态、克隆整合和损伤处理对香菇草前后端分株叶面积及净光合速率的影响
克隆整合(连接或者切断)显著影响香菇草基部与端部分株的叶面积 (P<0.05)(表2)。连接处理中,基部分株添加铵态氮-连接-无损伤处理及端部分株添加有机氮-连接-损伤处理的叶面积低于对应切断处理,其他连接处理的基部和端部分株的叶面积高于切断处理(图3a,c)。但是,克隆整合对香菇草基部与端部分株的净光合速率没有显著影响(P>0.05)。氮形态显著影响基部分株的叶片面积(P<0.05)(表2)。相对于对照而言,3种氮形态添加均提高了香菇草基部分株的叶面积,在硝态氮添加且无损伤时,叶面积最大(图3a)。氮形态对香菇草基部与端部分株的净光合速率没有显著影响(P>0.05),损伤及各因素的交互作用对香菇草基部与端部的叶面积与净光合速率均没有显著影响(P>0.05)(表2)。
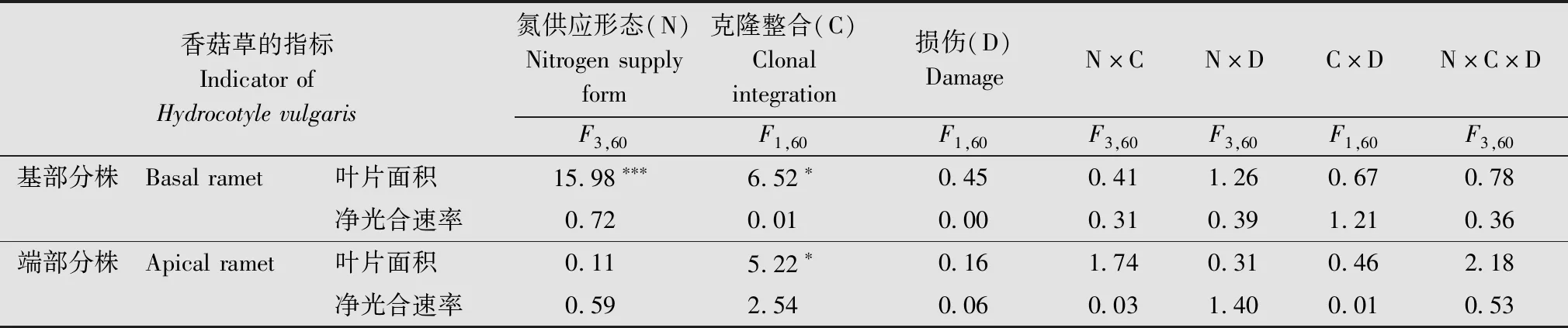
表2 氮供应形态、克隆整合及损伤处理对香菇草基部与端部分株叶面积与净光合速率影响的方差分析Table 2 ANOVA for effects of nitrogen supply form,clonal integration and damage on the leaf area and net photosynthetic rate of the basal and apical ramets of Hydrocotyle vulgaris
2.3 施氮形态、克隆整合和损伤处理对香菇草前后端分株氮含量的影响
克隆整合(连接或者切断)显著影响香菇草端部分株的总氮、根氮及茎氮含量(P<0.05)(表3),总体而言,连接处理的端部分株的氮含量高于切断处理(图4e~h)。但是,克隆整合对香菇草基部分株的氮含量没有显著影响(P>0.05)。不同施氮形态显著影响香菇草基部分株的总氮、根氮、茎氮与叶氮含量(P<0.05);对端部分株的根氮与茎氮也有显著影响(P<0.05)(表3)。总体来说,相对于对照,3种形式的氮添加显著提高了香菇草基部与端部分株的氮含量,并且对基部分株的提升更显著(图4)。此外,损伤处理显著影响香菇草端部分株的根氮与叶氮含量(表3,P<0.05)。具体而言,损伤处理提高了香菇草端部分株根氮和叶氮的含量(图4f,h)。

图2 氮供应形态、克隆整合及损伤处理对香菇草整个克隆片段,基部与端部分株生物量的影响Fig.2 Effects of nitrogen supply form,clonal integration and damage on the biomass of total clonal fragment,basal and apical ramets of Hydrocotyle vulgaris
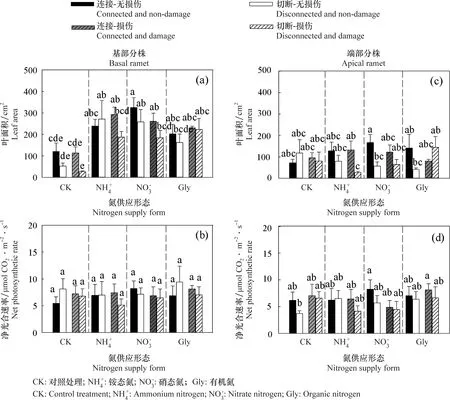
图3 氮供应形态、克隆整合及损伤处理对香菇草基部与端部分株叶面积与净光合速率的影响Fig.3 Effects of nitrogen supply form,clonal integration and damage on the leaf area and net photosynthetic rate of the basal and apical ramets of Hydrocotyle vulgaris

表3 氮供应形态、克隆整合及损伤处理对香菇草基部与端部分株氮含量影响的方差分析1)Table 3 ANOVA for effects of nitrogen supply form,clonal integration and damage on the nitrogen content of the basal and apical ramets of Hydrocotyle vulgaris

图4 氮供应形态、克隆整合及损伤处理对香菇草基部与端部分株氮含量的影响Fig.4 Effects of nitrogen supply form,clonal integration and damage on the nitrogen content of the basal and apical ramets of Hydrocotyle vulgaris
3 结论与讨论
试验结果表明,克隆分株之间的连接显著增加了香菇草整个克隆片段、基部分株以及端部分株的生物量和叶面积(表1,2;图2,3)。这意味着克隆整合对维持香菇草的生长发育具有重要意义。以往的研究也表明,通过克隆整合,克隆植物可以将各个分株间的资源整合并合理分配,从而促进整个克隆片段的生长[27-29]。但是值得注意的是,克隆分株之间的连接显著增加了端部分株的氮含量,但是对基部分株的氮含量并没有显著影响(表3,图4)。这可能是因为位于高资源生境的分株通常作为供体分株,将一些资源转移给位于低资源生境的分株[4,30]。因此,克隆整合促进了整个克隆片段及基部和端部分株的生长,特别是对端部分株的促进作用更显著。总体而言,基于事后多重比较的结果,氮形态与损伤并没有显著改变克隆整合对香菇草整个克隆片段总生物量的影响(图2)。以往的研究表明,氮添加通过提升克隆整合对克隆植物带来好处,进一步提升整个克隆植物片段的生长及适应性[14-15]。这种不一致的结果可能是因为本试验基部分株添加的氮浓度较低,以往的研究也表明,低氮添加对植物克隆整合的积极作用的影响较小[14-15,25]。值得注意的是,添加有机氮且损伤时,连接处理的香菇草整个克隆片段的生物量指标低于切断处理 (图2a),这可能是因为分株损伤诱发了克隆整合的负面效应,导致整个克隆片段生物量降低[24]。
氮是维持植物生长的重要养分[31-32]。本研究发现3种形态的氮添加均显著增加了香菇草整个克隆片段、基部分株以及端部分株的生物量 (表1,图2)。特别是硝态氮对香菇草生物量的积极影响最大。一方面,相较于有机氮,无机氮更容易被大多数植物吸收利用[33-34]。另一方面,我们的研究结果揭示,相较于铵态氮,香菇草可能更倾向于吸收利用硝态氮。以往的研究表明,多数杂草类外来入侵植物,如三叶鬼针草Bidenspilosa、五爪金龙Ipomoeacairica和微甘菊Mikaniamicrantha等均偏好硝态氮[35]。入侵杂草在原产地和入侵地常分布在硝态氮占优势的生境,长期适应高硝态氮的环境使这些外来入侵植物偏好硝态氮[19]。此外,相比于对照,铵态氮与有机氮添加也促进了香菇草的生长,因此,在高硝态氮输入地区应适当加强对香菇草的入侵防控。此外,高铵态氮与有机氮输入地区的入侵防控也不应忽视。
本研究表明,损伤并没有显著抑制香菇草的生长(图2~4)。研究表明,大多数植物具有一定的抵抗损害的能力[36-38],并且入侵植物对损害的耐受性和补偿性更强[39]。另外,不止一项研究表明,克隆植物可以通过合理的资源分配和分株来应对损伤[9,40-41]。值得注意的是,有研究发现分株损伤也可能诱发克隆整合的负面影响[24]。这种不一致的结果可能是由于该研究中的损伤是由长为15 cm的南方菟丝子Cuscutaaustralis寄生引起的[24],它可能需要更多的营养物质来维持生长,超过了克隆分株的抵抗损伤的限度。总之,本研究表明,香菇草作为一种入侵性克隆植物,具有抵抗一定程度损伤的能力。
本研究探讨3种形态氮供应及分株损伤对入侵植物香菇草生长及克隆整合的影响。结果表明,克隆整合对香菇草,特别是端部分株的生长发育有着重要的积极作用。3种施氮形态均促进了香菇草的生长,其中硝态氮对香菇草生长的促进更显著。分株损伤并没有显著抑制香菇草的生长。此外,氮形态与损伤并没有显著改变克隆整合对香菇草生长的积极作用。上述结果表明,入侵植物香菇草的克隆整合在其适应不同形态氮输入以及应对分株损伤中发挥着重要作用。此外,在高硝态氮输入地区应适当加强对香菇草的入侵防控。
