热带珊瑚岛不同生活型植物叶片性状及其生态适应性意义
2022-12-17覃张芬汤松波旷远文温达志王俊
覃张芬,汤松波,旷远文,温达志,王俊*
(1.中国科学院华南植物园,中国科学院海岛与海岸带生态恢复重点实验室,广东省应用植物学重点实验室,广州 510650;2.中国科学院大学,北京 100049)
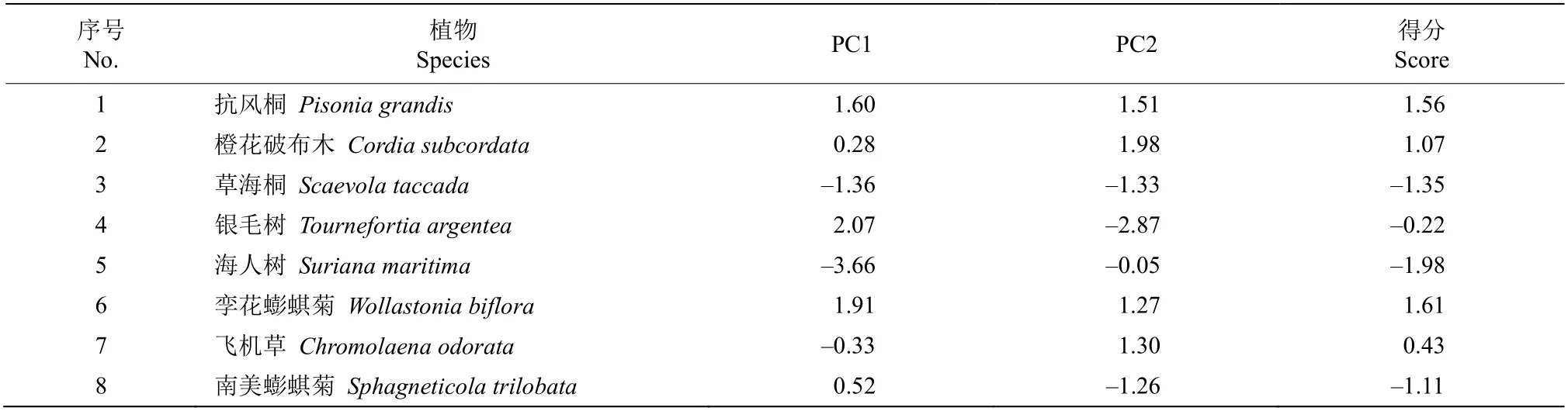
社会经济的快速发展导致人类需求与资源供给之间的矛盾日益突出[1]。植被作为热带珊瑚岛重要的资源,具有供给、调节、文化和支持等生态系统服务功能[2]。由于珊瑚岛独特的地理位置,其植被生境具有异质性、独立性、独特性和脆弱性特点[3]。环境胁迫下植物在叶片功能性状呈现系列变化特征。研究珊瑚岛植物对特殊生境的适应性机理可为其植被恢复和生态服务功能的维持提供重要参考依据[2,4]。
叶片作为连接植物与外界环境的重要桥梁,其功能性状与资源获取、光合产物分配密切相关。不同性状间的关联在一定程度上能反映植物在不同生境中的适应策略[5]。叶片比叶面积(specific leaf area,SLA)与资源(光和水)获取密切相关,影响光合速率,并指示植物碳(C)获取策略,通常,低SLA适应于养分贫瘠、干旱的生境,高SLA 表征植物具有较强的养分保持能力[6]。叶片元素化学计量特征,尤其是碳(C)、氮(N)、磷(P)计量比能反映植物的资源获取策略[7],并指示叶片结构和代谢投资间的平衡关系。植物在生长过程中可以通过改变元素化学计量特性来适应生境变化,如高生长速率的植物具有较高的C/N 和低N/P、C/P[8]。植物叶片的C/N、C/P 常用于表征N、P 利用效率[9]。当N/P<14 (以质量计算)时,表示植物生长受N限制,N/P>16 表示植物生长受P 限制;14 我国南海海域珊瑚岛分布广泛,但岛上植被类型相对简单,且优势种明确[16]。目前,已有从叶片功能性状变化特征角度探究植物对珊瑚岛生境适应的生理生态策略的研究,但多局限于单一物种[17–18]。不同生活型(乔、灌、草)植物对珊瑚岛特殊生境的适应机制是否存在差异还有待深入研究。本研究选取了西沙群岛中8 种优势植物,测定了其常见的叶片功能性状,以探究不同生活型植物叶片性状的差异性和相关性,为揭示不同生活型植物对珊瑚岛生境的适应策略和珊瑚岛植被恢复实践提供科学依据。 西沙群岛是中国南海群岛占地面积最大、生物多样性高及受环境变化较为敏感的区域,该地区受热带季风海洋性气候的影响,干湿季分明,雨季一般为6—11月,全年降雨量约1 500 mm,由于受风吹及强烈光照等因素影响,蒸发量约为降雨量2 倍,2—3月蒸发量甚至是降雨量10 倍之多[19]。珊瑚岛土壤由磷质石灰土和滨海盐土(海边冲积珊瑚砂)等组成,粗砂粒多,缺乏粘粒,硅、铁、铝等元素含量低,钙、磷,盐分含量高,呈强碱性(pH 8.0~9.5)[20]。 从《中国植物志》(http://www.iplant.cn/frps)确定岛上8 种优势植物的科、属、种、生活型和生活史等信息(表1)。每种植物至少选取5 株植株,采集50 片完全伸展、完整且生长良好的成熟功能叶片带回实验室。将新鲜叶片擦除灰尘后,选取10 片完整叶片用叶面积仪(Li-3000A,Li-Cor,Lincoln,NE,USA)测定叶面积(leaf area,LA,cm2),然后于105 ℃杀青,65 ℃下烘干,称取叶片干质量(leaf dry weight,LDW,g),计算比叶面积(SLA,cm2/g)=LA/LDW。 表1 珊瑚岛8 种代表性物种基本信息Table 1 Information of eight species in tropical Coral Islands,China 剩余叶片经干燥、研磨、过筛,用于测定全碳(C)、全氮(N)、全磷(P)和纤维素含量。全C 含量采用重铬酸钾-硫酸氧化法测定,全N 含量采用靛酚蓝比色法测定,全P 含量定采用钼锑抗比色法测定[21]。用同位素质谱仪(Thermo Finnigan,North Pod Waltham,美国)测定叶片同位素比值(δ13C),根据Huang等[22]的方法计算植物的水分利用效率(WUE)。 分别采用Shapiro-Wilk 和Levene检验对数据进行正态性检验和方差齐性分析检验。采用单因素方差分析比较不同物种叶片功能性状的差异; 采用Pearson 相关系数分析叶片性状间的相关性;采用主成分分析(principal component analysis,PCA)对不同物种叶片性状进行排序分析。数据处理和可视化均采用软件R 4.0.2 完成。显著性水平为P<0.05。 3 种生活型植物叶片的全C 含量为401.47~419.82 mg/g,乔木叶片的全N 含量 (28.28 mg/g)明显高于灌木叶片(15.98 mg/g),叶片的全P 含量为2.72~3.07 mg/g。3 种生活型植物叶片的C/P、N/P 和全C、全P含量差异不显著,C/P为142.71~167.43,N/P为3.30~10.02。但C/N 在3 种生活型植物中差异显著,以灌木最高(26.95),乔木最低(14.21)。乔木与灌木叶片的纤维素和全N 含量及SLA、WUE 差异显著,乔木与草本叶片的C/N、WUE 差异显著,灌木与草本的SLA、C/N 和全N 含量差异显著(图1)。 图1 珊瑚岛不同生活型植物叶片性状的比较。柱上不同字母表示差异显著(P<0.05)。SLA: 比叶面积; Cellulose: 纤维素; WUE: 水分利用效率; C: 全碳; N: 全氮; P: 全磷。下同Fig.1 Comparison of leaf functional traits among different life forms in coral islands.Different letters upon column indicate significant differences at 0.05 level.SLA: Specific leaf area; Cellulose: Cellulose; WUE: Water use efficiency; C: Total carbon; N: Total nitrogen; P: Total phosphorus.The same below 从表2可见,叶片的C/N 与WUE、全C 含量呈显著正相关,与SLA、全N 含量和N/P 呈极显著负相关;WUE 与全N 含量呈极显著负相关。C/P与N/P、纤维素含量呈极显著正相关,与全C、全P含量呈极显著负相关。纤维素含量与全C、全P 含量和SLA 呈极显著负相关。叶片全C 含量与N/P呈极显著负相关,与全P 含量呈极显著正相关。全P含量与N/P 呈极显著负相关。SLA 与全N 含量呈极显著正相关,叶片全N 与N/P 呈极显著正相关。 表2 植物叶片性状间的相关性Table 2 Correlations of leaf traits of coral island plants 主成分1 (PC1)和主成分2 (PC2)可解释9 个叶片性状76.3%变异,解释率分别为总方差的40.9%和35.4%。在PC1 上物种得分为银毛树>孪花蟛蜞菊>抗风桐>南美蟛蜞菊>橙花破布木>飞机草>草海桐>海人树;PC2 上得分为橙花破布木>抗风桐>飞机草>孪花蟛蜞菊>海人树>南美蟛蜞菊>草海桐>银毛树(表3)。孪花蟛蜞菊综合得分最高,海人树最低。8 种植物在物种-性状排序图中发生了明显趋异分化,大致分为3 个部分(图2):银毛树种置信区间(95%)位于PC1 正向区域与PC2 负向区域之间,具有较低的C/N、SLA、全C 含量和较高N/P、WUE、纤维素含量; 海人树置信区间位于PC1负向区域,具有较高的全C 含量、C/N 及较小N/P、全N;其他植物的置信区间均分布在整个PC1、PC2正、负向区域。 图2 珊瑚岛植物叶片性状的主成分分析Fig.2 Principal component analysis of leaf traits of species 表3 8 种植物的主成分因子得分Table 3 Scores of 8 species by principal component analysis 植物在长期进化过程中,经过环境过滤和自然筛选,形成一系列组合性状以适应环境变化[23–24]。淡水、土壤养分、光照强度、风力强度等是影响海岛植被恢复的重要生态因子[25]。本研究结果表明,珊瑚岛乔木生活型与草本生活型植物SLA、N/P 和全N、全P 含量均显著大于灌木生活型植物,而C/N、C/P、WUE 和纤维素含量则相反,反映了不同生活型植物叶片功能性状的显著差异。本研究中3 种生活型植物的SLA 均与沙丘生境优势种相当(86~295 cm2/g)[26],说明珊瑚岛3 种生活型植物抗旱能力强; 而全C 含量低于荒漠优势植物(435.07 mg/g)[27],可能是3 种生活型植物为适应海岛恶劣环境,通过降低SLA,防止水分散失,从而影响了叶片光合能力,这与前人[28]的研究结果一致。珊瑚岛3 种生活型植物叶片全N含量(15.98~28.28 mg/g)和全P 含量(2.72~3.07 mg/g)均高于海南岛热带低地雨林植物(0.5~0.8 mg/g),与正常生长植物相当[N (10~50 g/kg)、P (0.3~4 g/kg)],说明3 种生活型生长状况良好,能适应海岛相对缺N 富P 土壤环境[29–30]。珊瑚岛乔木生活型植物叶片的C/N (14.21 g/kg)低于荒漠典型植被,而灌木生活型、草本生活型植物大于荒漠典型植被,以灌木生活型最大(26.96 g/kg),说明灌木生活型植物的N 利用效率最高。而珊瑚岛植物的C/P 显著低于海南岛热带低地雨林植物(1 240~1 260),也低于荒漠典型植物(368.70)[31],灌木生活型植物的C/P 最高,说明珊瑚岛3 种生活型植物对P 利用效率均较低,以灌木生活型植物对P 利用效率相对较高。珊瑚岛3 种生活型植物N/P<14,表明其生长受N 限制,这与热带陆地森林地区(P限制)相反,可能是因为珊瑚岛特殊成土过程,土壤中P 供应大于N 供应有关,也说明叶片的营养元素变化受土壤养分调节[32–33]。3种生活型中,灌木生活型植物SLA、全N 含量最低,而WUE 最高。有研究报道海岛植被种类及面积呈缓慢增加趋势,灌木生活型植物,如草海桐在2006—2018年期间由占据较小面积逐渐发展成单优物种,这可能是因为灌木生活型植物具有低SLA和全N 含量及高WUE 特性而在海岛生境中具有优势[34]。综上,珊瑚岛植被不同生活型叶片性状间差异显著。 珊瑚岛植物叶片的C/N 与WUE,C/P 与纤维素含量、N/P,全N 含量与SLA、N/P,全C 含量与全P 含量呈极显著正关系;而C/N 与N/P、SLA、全N 含量,WUE 与全N 含量,C/P 与全C 和全P含量,纤维素含量与全C 和全P 含量及SLA,N/P与全C 和全P 含量呈极显著负关系,其中C/N 与全N 含量的相关性最强。这表明植物叶片N 含量越低,N 利用效率越高,WUE 越高,叶片厚度较大、面积较小,且叶性状间关联性强,存在碳水化合物储备投资与生长间的权衡关系,这与前人[6,14,35–36]的研究结论相似,但WUE 与叶片C/N、全N 含量间的关系与前人研究结论相反,可能是植物在长期适应珊瑚岛特殊生境过程中采取的耐旱策略,即植物为适应低N 土壤养分、高太阳辐射等胁迫环境而采取的生活策略。WUE 的高低主要取决于植被初级生产力与水分蒸散作用,全N 含量与WUE 呈负相关、C/N 与WUE 呈正相关,可能是因为海岛植物在受到水分、强光胁迫会通过降低蒸腾速率及植被初级生产力来抵抗胁迫[2]。蒸腾速率受气孔密度、叶片结构、叶片碳水化合物含量影响,植物在干旱胁迫下,通常会提高N 含量用于构建叶片细胞壁保护性物质的投入来降低机械损伤,但同时也会降低光合蛋白的投入[2,33,37]。叶片性状间复杂的关联性,体现了珊瑚岛植被存在碳水化合物分配与生长投资之间的权衡关系。 不同物种具有不同的资源利用策略[38]。主成分分析表明,PC1、PC2 主要反映叶片养分投资与回收权衡,PC1 主要反映N、P 分配能力、光合固C能力的保守型生态策略,得分最高的是灌木银毛树,最低的是灌木海人树,说明银毛树叶片更倾向于缓慢生长,叶片全N 含量较高及较低N 利用效率、光合固C 能力的保守型生态策略。PC2 主要反映高SLA、N 储存、低纤维素含量与WUE 的资源获取型生态策略,得分最高的是橙花破布木,最低的是银毛树,说明橙花破布木相对于其他7 种植物更倾向于储存营养物质策略,即具有较高的SLA 和叶片全N 含量,较低WUE 和纤维素含量。当环境中的资源可利用性降低时,植物的根系对地下资源竞争会增大,甚至比地上部分对光资源的竞争更大[39–40]。乔木较倾向于资源获取型策略可能是因为其具有较大、复杂的根系,如抗风桐根部的根瘤菌可以进行生物固N,比较快速地吸收土壤养分且对光资源的需求更为强烈,因此橙花破布木、抗风桐优势乔木在珊瑚岛生境中具有较高的叶片全N 含量及SLA,细胞壁构建物质纤维素含量较低,当植物积累光合物质时,水分因为气孔的开放而降低对水分的利用效率[40–44]。综合分析表明,乔木的抗风桐、灌木的银毛树、草本的孪花蟛蜞菊表现出对热带珊瑚岛生境较强的生态适应性。 对热带珊瑚岛不同生活型植物生态策略的研究表明,不同植物采取不同的生态生存策略,并发生了明显的趋异分化,其中银毛树更倾向于缓慢生长,表现为保守型生态策略,橙花破布木更倾向于储存营养物质的资源竞争策略,即具有较高的SLA和叶片全N 含量,较低的WUE 和纤维素含量。在未来珊瑚岛礁植物恢复实践中,乔木可以优先采用抗风桐,灌木可以优先采用银毛树,但同时要考虑到飞机草、孪花蟛蜞菊、南美蟛蜞菊为我国入侵种(GISD,http://www.iucngisd.org/gisd/; 植物智,http://www.iplant.cn/),比海岛本地种(如海人树)表现出更强的适应性,因此应防止上述物种的扩张。1 材料和方法
1.1 样地概况
1.2 方法

1.3 数据分析
2 结果和分析
2.1 不同生活型植物叶片性状

2.2 叶片性状间的相关性

2.3 叶片性状的主成分分析

3 结论和讨论
3.1 不同生活型植物叶片性状差异
3.2 珊瑚岛植物叶片性状间相关性
3.3 珊瑚岛植物叶片性状间的权衡
