不同马尾松林下光环境中油茶叶片和光合特征比较及其与主要环境因子的相关性
2022-12-16张亚琴郭其强罗丝琼盘金文郭有燕
张亚琴, 郭其强,①, 罗丝琼, 盘金文, 姚 珊, 郭有燕
(1. 贵州大学林学院 贵州省森林资源与环境研究中心, 贵州 贵阳 550025; 2. 河西学院乡村振兴研究院, 甘肃 张掖 734000)
油茶(CamelliaoleiferaAbel)为山茶科(Theaceae)山茶属(CamelliaLinn.)常绿灌木,是一种在中国南方地区广泛种植的优良油用经济树种。油茶籽油富含不饱和脂肪酸,具有抗氧化和增强免疫力等作用[1]。贵州是全国重点油茶产区之一[2,3],并且该地区约30%的油茶种植在马尾松(PinusmassonianaLamb.)林下[4]。然而,随着马尾松个体的不断生长,林下的光照强度和光照持续时间逐渐降低,显著抑制了油茶的生长[5]。如何科学地提高马尾松林下油茶产量成为贵州油茶产业发展中亟待解决的重要问题。
光照是影响植物生长、发育和空间分布的一个重要因子[6],通过植物的形态和光合特性变化能够分析植物的光合能力及其对环境的响应策略[7]。国内学者已经对部分油茶资源的光合能力进行了比较,并发现了一些优质资源。例如:吴晓龙等[8]对2个油茶品种〔‘湘林1’(‘Xianglin 1’)和‘湘林97’(‘Xianglin 97’)〕及6个近缘种叶片光合特性的日变化规律进行了研究,发现供试的2个油茶品种均具有较高的光合效率和光合能力;何一明等[9]认为,疏植油茶的光合效率高于密植油茶;唐炜等[10]比较了油茶品种‘湘林1号’(‘Xianglin No. 1’)、攸县油茶(C.yuhsienensisHu)和宛田红花油茶(C.polyodontaHow ex Hu)的光合特性日变化,发现攸县油茶的净光合速率最高。值得注意的是,虽然光照对植物的生长发育有较大影响,但并不是所有植物都适合生长在强光照环境中,有些植物更适合生长在适度遮光的环境中,例如:桃(AmygdaluspersicaLinn.)和鳞尾木(LepionurussylvestrisBl.)[11,12]。Guo等[13]的研究结果表明:气温、光合有效辐射和空气水汽压亏缺3个环境因子可显著影响桃儿七〔Sinopodophyllumhexandrum(Royle) Ying〕叶片的气孔特征和光合作用,中等强度的光合有效辐射可以促进桃儿七的叶片发育和光合能力。探明马尾松林下不同光照环境中油茶的生长状况,筛选出适宜马尾松林下油茶生长的光照条件,对于提高马尾松林下油茶产量至关重要。
鉴于此,本文对贵州大学林学院实验基地内不同马尾松林下光环境(光下无遮挡处、林缘和林内)中的油茶叶片和光合特征差异进行了比较,对3种林下光环境中光合有效辐射、气温和空气水汽压亏缺的日变化差异进行了比较,并对油茶叶片和光合特征与上述3个主要环境因子进行了Pearson相关性分析,以期阐明不同光照条件下油茶叶片的形态、叶绿素含量、气孔形态和光合特征差异及其与主要环境因子的关系,为油茶复合种植,尤其是马尾松林下油茶产量的提高和科学管理提供实验依据。
1 研究区概况和研究方法
1.1 研究区概况
供试油茶植株种植在贵州大学林学院实验基地(具体地理坐标为东经106°39′20″、北纬26°27′18″,海拔1 116 m)。该区域属亚热带湿润季风气候,年平均气温15.3 ℃,最热月(7月)平均气温24.6 ℃,最冷月(1月)平均气温4.9 ℃,全年积温3 700 ℃~4 710 ℃;年平均降水量1 174.7 mm,年平均空气相对湿度约74%;无霜期约270 d。基地土壤类型为黄壤,pH 5.0。
供试林分内马尾松株龄20 a,平均胸径约22 cm,平均树高约11 m;供试油茶株龄10 a,近7 a基本处于无人经营管理状态。
1.2 材料
于2020年7月下旬,在光下无遮挡处(对照)、林缘和林内选择生长良好且无病虫害的油茶植株各5株,做好标记。利用3415FQF光量子照度双辐射计(美国Spectrum 公司),在连续3 d天气晴朗日的8:00—18:00间每2 h测定1次光照强度,分别测定每株油茶东、西、南、北4个方向距离地面20 cm处的光照强度,结果取平均值。不同马尾松林下光环境中油茶样株的基本情况见表1。

表1 不同马尾松林下光环境中油茶样株的基本情况
1.3 方法
1.3.1 叶片特征观测
1.3.1.1 形态指标测定 于2020年7月25日至30日,采集所有样株一级侧枝中部长势均匀的完整叶片,即枝条顶端向下的第3和第4枚功能叶,每株随机选取10枚叶片,分别测定叶片的形态指标。其中,叶面积和叶周长使用AM-350叶面积测量仪(英国ADC公司)测量;叶片厚度使用数显游标卡尺(精度0.01 mm)测量;比叶面积参照何小燕[14]的方法测定,先将叶片置于85 ℃条件下烘干至恒质量,再使用千分之一电子天平称量叶片的干质量,根据叶面积和叶片干质量计算比叶面积,计算公式为比叶面积=叶面积/叶片干质量。
1.3.1.2 叶绿素含量测定 在测定叶片形态特征的同时,使用SPAD-502叶绿素仪(日本Konica公司)测定上述叶片的叶绿素相对含量(SPAD),根据回归方程y=0.236 0e0.058 8x(r2=0.850 0)计算叶绿素含量,其中,x为SPAD值,y为叶绿素含量[15]。
1.3.1.3 气孔形态观测 在测定叶片形态特征的同时,参照刘明智等[16]的方法,用清水将上述叶片表面冲洗干净,擦干叶片表面的水分,将指甲油均匀涂在叶片表面,置于阴凉、通风处2 h;待叶片表面成膜后,剪取叶片中部面积2 mm×2 mm的小块,用镊子撕下叶片表皮,固定在载玻片上,制成临时玻片;在DM2500光学显微镜(德国Leica公司)下观察气孔形态,选取典型视野,用Leica Application Suite X 3.0.2软件拍照,用Image-J软件测量气孔面积,并统计气孔密度。每株观测30个气孔。
1.3.2 叶片光合特征观测
1.3.2.1 光响应曲线拟合 于2020年7月28日至30日间晴朗日的9:00—11:00,选取供试样株一级侧枝顶端向下生长状况良好且健康的第3和第4枚功能叶,使用Li-6400便携式光合仪(美国LI-COR公司)测定叶片的光合特征参数,每枚叶片重复测定3次。测定时,采用红蓝人工光源叶室,叶室CO2浓度为(370±5) μmol·m-2·s-1,叶室温度为(25.0±0.5) ℃,设定系列光合有效辐射为1 800、1 600、1 400、1 200、1 000、800、600、400、300、200、100、50和0 μmol·m-2·s-1,每个光合有效辐射水平适应120~150 s。根据测定结果,利用非直角双曲线模型[12]拟合光响应曲线,通过拟合的光响应曲线计算最大净光合速率、表观量子效率、光饱和点、光补偿点和暗呼吸速率。
1.3.2.2 光合参数日变化分析 在拟合光响应曲线的同时,选择供试样株一级侧枝顶端向下生长状况良好且健康的第3和第4枚功能叶,使用Li-6400便携式光合仪测定叶片的光合参数日变化,每日测定时间为8:00—18:00,每1 h测定1次,每枚叶片重复测定3次。测定时,叶室CO2浓度为(370±5) μmol·m-2·s-1,叶室温度为(25.0±0.5) ℃,空气流速为(500±1)μmol·s-1。认真记录净光合速率、蒸腾速率、气孔导度和胞间CO2浓度,根据测定结果计算水分利用效率和气孔限制值,计算公式分别为水分利用效率=净光合速率/蒸腾速率和气孔限制值=1-胞间CO2浓度/大气CO2浓度,其中,大气CO2浓度为370 μmol·m-2·s-1。
1.3.3 环境因子日变化分析 在测定光合参数的同时,使用Li-6400便携式光合仪记录光合有效辐射、气温和空气水汽压亏缺。
1.4 数据处理及分析
采用Microsoft EXCEL 2010软件整理相关数据;采用SPSS 26.0软件进行数据统计分析,采用单因素方差分析(one-way ANOVA)法分析各指标在不同林下光环境间的差异(α=0.05),并对油茶叶片和光合特征与3个主要环境因子进行Pearson相关性分析;采用Origin 2018软件作图。
2 结果和分析
2.1 不同马尾松林下光环境中油茶叶片特征的比较
不同马尾松林下光环境中油茶叶片形态特征统计结果(表2)表明:油茶叶片的面积、周长和厚度在林缘最高、在林内最低,且这3个指标在不同林下光环境间差异显著(P<0.05);比叶面积在林缘最高、在光下无遮挡处(对照)最低,且该指标在林缘和林内的差异不显著,但均显著高于对照。
观测结果(表2和图1)表明:油茶叶片的气孔呈椭圆形,仅分布于叶片的下表皮。比较而言,气孔密度在林缘最大(238.6 mm-2)、在林内最小(172.6 mm-2),而气孔面积则在对照中最大(210.73 μm2)、在林缘最小(120.94 μm2),且气孔密度和气孔面积在不同林下光环境间差异显著。
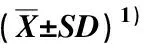
表2 不同马尾松林下光环境中油茶叶片的形态特征
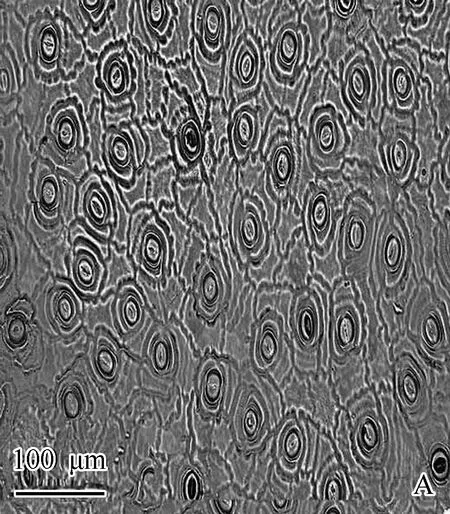


A: 对照(光下无遮挡处)The control (the place without shelter under light); B: 林缘Forest edge; C: 林内Forest inside.
2.2 不同马尾松林下光环境中油茶叶片光合特征的比较
2.2.1 叶绿素含量比较 由不同马尾松林下光环境中油茶叶片叶绿素含量统计结果(表3)可见:油茶叶片的叶绿素含量在林缘最高(15.69 mg·g-1)、在林内最低(12.92 mg·g-1),且在林缘和光下无遮挡处(对照)的差异不显著,但均显著(P<0.05)高于林内。

表3 不同马尾松林下光环境中油茶叶片的叶绿素含量
2.2.2 光响应曲线比较 从不同马尾松林下光环境中油茶叶片的光响应曲线(图2)可以看出,随着光合有效辐射的提高,对照、林缘和林内油茶叶片的净光合速率均呈先急剧升高后趋于稳定的变化趋势。当光合有效辐射低于300 μmol·m-2·s-1时,对照叶片净光合速率在3种林下光环境中最低。当光合有效辐射在300~1 800 μmol·m-2·s-1时,叶片净光合速率在林缘最高、在林内最低,且在林缘的数值明显高于林内和对照。当光合有效辐射为1 800 μmol·m-2·s-1时,林缘叶片净光合速率达6.29 μmol·m-2·s-1,林内叶片净光合速率为3.59 μmol·m-2·s-1。

: 对照(光下无遮挡处)The control (the place without shelter under light); : 林缘Forest edge; : 林内Forest inside.
利用拟合的光响应曲线计算3种林下光环境中油茶叶片的光合参数,结果见表4。由表4可见:油茶叶片最大净光合速率在3种林下光环境中由大到小依次为林缘、对照、林内,其中,林缘叶片最大净光合速率为6.29 μmol·m-2·s-1,显著高于对照和林内,且后2种林下光环境间的叶片最大净光合速率差异显著。3种林下光环境中油茶叶片表观量子效率为0.037~0.055 μmol·m-2·s-1,且在不同林下光环境间差异显著,其中,林缘的叶片表观量子效率最高,分别较对照和林内增加了22.22%和48.65%。3种林下光环境中油茶叶片的光饱和点、光补偿点和暗呼吸速率存在显著差异,其中,叶片光饱和点在林缘最高(1 410.28 μmol·m-2·s-1)、在林内最低(761.57 μmol·m-2·s-1),林缘的叶片光饱和点分别较对照和林内增加了39.86%和85.18%;叶片光补偿点在3种林下光环境中从大到小依次为对照、林缘、林内,其中,对照的叶片光补偿点为25.56 μmol·m-2·s-1,林内的叶片光补偿点为6.63 μmol·m-2·s-1;叶片暗呼吸速率在林缘最高(1.03 μmol·m-2·s-1)、在林内最低(0.54 μmol·m-2·s-1),林缘的叶片暗呼吸速率分别较对照和林内增加了58.46%和90.74%。
2.2.3 光合参数日变化比较 不同马尾松林下光环境中油茶叶片净光合速率的日变化曲线(图3-A)显示:在8:00—10:00和17:00—18:00间,叶片净光合速率从大到小依次为对照、林缘、林内;在11:00—16:00间叶片净光合速率从大到小依次为林缘、对照、林内(14:00除外)。总体来看,3种林下光环境中油茶叶片净光合速率的日变化呈现“升—降—升—降”的趋势,且均有2个峰值,其中,对照和林内叶片净光合速率的2个峰值均出现在10:00和14:00,但出现最高值的时间不同,对照叶片净光合速率在10:00最高,林内叶片净光合速率在14:00最高;林缘叶片净光合速率的2个峰值出现在10:00和13:00,且在13:00最高。
不同马尾松林下光环境中油茶叶片气孔导度的日变化曲线(图3-B)显示:在8:00—10:00和14:00—18:00间,叶片气孔导度从大到小依次为对照、林缘、林内;在11:00—12:00,叶片气孔导度从大到小依次为林缘、对照、林内;在13:00,叶片气孔导度从大到小依次为林缘、林内、对照。总体来看,3种林下光环境中油茶叶片气孔导度的日变化均呈现波动变化趋势,对照叶片气孔导度在8:00最高、在18:00最低,林缘叶片气孔导度在12:00最高、在18:00最低,林内叶片气孔导度在13:00最高、在18:00最低。
表4 不同马尾松林下光环境中油茶叶片光响应曲线的拟合参数



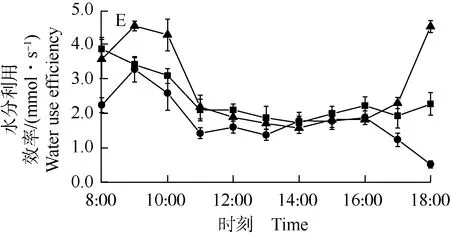
不同马尾松林下光环境中油茶叶片胞间CO2浓度的日变化曲线(图3-C)显示:在8:00—13:00和17:00—18:00间,林内叶片胞间CO2浓度最高,多数时刻林缘叶片胞间CO2浓度高于对照;在14:00—16:00间,对照叶片胞间CO2浓度最高,林缘叶片胞间CO2浓度在14:00—15:00间高于林内、在16:00低于林内。总体来看,3种林下光环境中油茶叶片胞间CO2浓度的日变化均呈现先降低后升高的趋势,最高值均出现在8:00,但最低值略有差异,对照和林缘叶片胞间CO2浓度的最低值出现在13:00,林内叶片胞间CO2浓度的最低值则出现在14:00。
不同马尾松林下光环境中油茶叶片蒸腾速率的日变化曲线(图3-D)显示:在8:00—10:00和14:00—18:00间,对照叶片蒸腾速率基本上最高,多数时刻林缘叶片蒸腾速率高于林内;在11:00—13:00间,林缘叶片蒸腾速率最高,林内叶片蒸腾速率在11:00和13:00高于对照、在12:00低于对照。总体来看,3种林下光环境中油茶叶片蒸腾速率的日变化均呈先升高后降低的趋势,最低值均出现在18:00,但最高值略有差异,林缘和林内叶片蒸腾速率的最高值出现在13:00,对照叶片蒸腾速率的最高值则出现在14:00。
不同马尾松林下光环境中油茶叶片水分利用效率的日变化曲线(图3-E)显示:在8:00和12:00—16:00间,林缘叶片水分利用效率基本上最高,多数时刻对照叶片水分利用效率高于林内;在9:00—11:00和17:00—18:00间,叶片水分利用效率从大到小依次为对照、林缘、林内。总体来看,3种林下光环境中油茶叶片水分利用效率的日变化略有差异,其中,对照叶片水分利用效率日变化表现为“升高—降低—相对稳定—升高”的趋势,林缘叶片水分利用效率日变化表现为“降低—相对稳定—升高”的趋势,林内叶片水分利用效率日变化表现为“升高—降低—相对稳定—降低”的趋势。
不同马尾松林下光环境中油茶叶片气孔限制值的日变化曲线(图3-F)显示:不同时刻对照、林缘和林内叶片气孔限制值无明显规律,但总体来看,3种林下光环境中油茶叶片气孔限制值的日变化趋势与胞间CO2浓度的日变化趋势恰好相反,基本上表现为先升高后降低的趋势。其中,对照和林缘叶片气孔限制值的最低值均出现在8:00,最高值均出现在13:00;林内叶片气孔限制值的最低值出现在18:00,最高值出现在14:00。
2.3 不同马尾松林下光环境中环境因子日变化的比较
不同马尾松林下光环境中气温、空气水汽压亏缺和光合有效辐射的日变化曲线见图4。由图4可见:总体来看,3种林下光环境中气温、空气水汽压亏缺和光合有效辐射的日变化趋势均表现为先升高后降低的变化规律,且气温和空气水汽压亏缺的日变化趋势更为相似。3种林下光环境中气温和空气水汽压亏缺的最高值均出现在13:00或14:00,而光合有效辐射的最高值则出现在12:00;总体来看,气温和空气水汽压亏缺的最低值出现在8:00,且明显低于18:00的数值。



: 对照(光下无遮挡处)The control (the place without shelter under light); : 林缘Forest edge; : 林内Forest inside.
2.4 油茶叶片和光合特征与主要环境因子的相关性分析
油茶叶片和光合特征与上述3个主要环境因子的相关性分析结果(表4)表明:油茶叶片的比叶面积和气孔面积与光合有效辐射呈极显著(P<0.01)正相关,其余油茶叶片形态特征和叶绿素含量与气温、空气水汽压亏缺和光合有效辐射的相关性均不显著。除了气孔导度与气温和空气水汽压亏缺以及水分利用效率与光合有效辐射的相关性不显著外,其余光合参数与气温、空气水汽压亏缺和光合有效辐射的相关性均达到显著(P<0.05)或极显著水平,其中,净光合速率、蒸腾速率和气孔限制值与3个主要环境因子均呈正相关,气孔导度与光合有效辐射与3个主要环境因子也呈正相关,而胞间CO2浓度和水分利用效率与3个主要环境因子呈负相关。

表4 油茶叶片和光合特征与主要环境因子的相关性分析
3 讨论和结论
众所周知,植物叶片形态特征与其受到的光照强度密切相关[5]。在供试的3种林下光环境中,林缘油茶叶片的面积、周长、厚度和比叶面积均最高,且林缘多数指标显著高于光下无遮挡处(对照)和林内;林内油茶叶片的比叶面积显著高于对照,但叶片的面积、周长和厚度均显著低于对照。说明马尾松林缘的光照条件更适合油茶叶片的生长,林内的光照条件则不利于油茶叶片的生长。气孔是植物与外界环境进行水气交换的主要器官[17,18]。油茶叶片的气孔呈椭圆形,仅分布于叶片的下表皮。在供试的3种林下光环境中,林内油茶叶片的气孔密度显著低于林缘和对照,气孔面积显著高于林缘但显著低于对照,说明不同光照条件下,油茶可通过调节气孔密度和气孔面积来维持其叶片的光合能力和蒸腾水平。推测这是油茶对不同光照生境的适应性策略。气孔形态的这种变化不仅促进了油茶的气体交换,还促进了植株对水分的利用[19]。
植物光补偿点能体现植物利用弱光的能力,其数值越低,表明植物的弱光利用能力越强[20];而植物光饱和点越高,表明其光利用率越高[21]。在供试的3种林下光环境中,林内油茶叶片的光补偿点最低,仅为6.63 μmolm-2s-1,林缘油茶叶片的光饱和点最高,达到1 410.28 μmol·m-1·s-1,说明供试马尾松林内油茶叶片利用弱光的能力较强,而林缘油茶叶片的光利用率较高,有利于其体内有机物的积累。
叶绿素是评价植物光合作用水平的一个重要指标[22],能够反映植物叶片的净光合速率水平[23]。比较发现,林缘油茶叶片的叶绿素含量最高,达到15.69 mg·g-1,说明马尾松林缘油茶叶片的光合能力最强,推测这可能是因为在林缘环境中,为了叶片能够获得更多的光能,油茶将体内更多的碳分配给叶片以提高叶片中的叶绿素含量,从而提高油茶叶片的光合作用水平[24]。
叶片净光合速率是衡量植物光合作用能力的直接指标,数值越高,表明植物叶片的光合能力越强,对环境的适应性也越强[25,26]。研究发现,气温、空气水汽压亏缺和光合有效辐射等环境因子通过影响植物的形态特征(如比叶面积和叶绿素含量)和光合参数(如净光合速率、空气导度、胞间CO2浓度、蒸腾速率、水分利用效率和气孔限制值),间接影响植物的净光合速率,其中,气温、空气水汽压亏缺和光合有效辐射为主要影响因子[27,28]。本研究结果显示:油茶叶片的净光合速率与气温、空气水汽压亏缺和光合有效辐射分别呈极显著、显著和极显著正相关,其叶片的蒸腾速率和气孔限制值也与上述3个环境因子呈显著或极显著正相关,而其叶片的胞间CO2浓度和水分利用效率则与上述3个环境因子呈显著或极显著负相关(水分利用效率与光合有效辐射的相关性除外)。说明气温、空气水汽压亏缺和光合有效辐射均对油茶叶片的光合能力有较大影响,应在油茶栽培和推广应用过程中注意栽培地的这些环境因子条件是否适宜油茶生长。
综合分析认为,生长在马尾松林缘的油茶叶片生长较好,光合能力较强。在实际生产过程中,可通过定期疏伐、修枝等措施,改善林下光合有效辐射条件,营造有利于油茶生长的环境条件,为马尾松林下油茶生长和产量提高提供技术保障。
