新甲基橙皮苷二氢查耳酮及合成前体药理作用研究进展
2022-12-06宋若兰王振东钟祥健折改梅
魏 静,宋若兰,陈 翔,王振东,董 英,钟祥健,吕 芳,折改梅,*
(1.北京中医药大学中药学院,北京 102488;2.桓台史氏医院,山东 淄博 256405;3.山东奔月生物科技股份有限公司,山东 东营 257000)
新甲基橙皮苷二氢查耳酮(Neohesperidin dihydrochalcone,NHDC)为高甜度、低热量的甜味剂,于1994 年被欧盟批准作为食品甜味剂在食品行业中应用,其甜度为蔗糖的1500~1800 倍,是目前研究人员所发现的最佳药用甜味剂和屏味剂[1-3]。随着NHDC在食品、营养学等行业中应用增多,近几年NHDC的药理研究逐渐成为关注热点。现代药理学研究表明,NHDC 并不会抑制口服耐受的建立,从而避免诱导食物过敏[2]。此外,除对稳定的自由基和活性氧有显著的剂量依赖性清除作用[4-7],还具有保护肝脏[5,8]和降血脂[9]等药理作用,有望用作保肝剂、治疗或预防脂质水平升高疾病新原料等[1],但其生物学特性和临床应用需要更深入的研究。
目前国内多集中于NHDC 的制备工艺研究,其合成方法日臻成熟,但药理作用及相关机制研究较少[1,3,10-11]。当前多采用橙皮苷(Hesperidin,HSD)以生物合成法或由柚皮苷(Naringin,NRG)以化学合成法制备新橙皮苷(Neohesperidin,NHP),再由NHP在碱性条件下经氢化还原后得到NHDC[1]。NHDC合成前体的药理作用研究较为成熟,涉及心血管系统、神经系统和生殖系统等多个方面,具有多种生物学特性,如抗癌[12-14]、抗氧化[15-17]、抗炎[18-19]、降血糖[20-22]、降血脂[23-25]、保护肝脏[26-27]等作用,对人体健康有益[28-30]。故鉴于其合成前体广泛的药理作用,本文主要对NHDC 及其合成前体代表性文献中的药理作用进行综述并分析,以期为功能性甜味剂NHDC后续开发提供理论信息。
1 NHDC 的合成及合成前体结构
NHDC 为二氢查耳酮类化合物,通常认为是一种半合成的糖苷查耳酮,分子式为C28H36O15,糖基为新橙皮糖,由L-鼠李糖基通过α-1,2-糖苷连接到β-葡萄糖苷残基组成[31]。本研究团队在多叶棘豆植物中发现有天然的NHDC 存在[32],其含量无法满足市场需求。为实现大规模工业化生产,市场多由HSD、NHP 和NRG 等前体进行合成[33]。HSD 分子式为C28H34O15,化学名称为橙皮素-7-芸香糖苷[34];NHP为HSD 的异构体,两者区别在于HSD 糖基中鼠李糖连接葡萄糖的6 位,NHP 糖基中鼠李糖连接葡萄糖的2 位[35];NRG 分子式为C27H32O14,由柚皮素和新橙皮糖组成[36]。NHDC 及其合成前体结构如图1所示。

图1 新甲基橙皮苷二氢查耳酮及其合成前体结构[1,34-36]Fig.1 Chemical structures of neohesperidin dihydrochalcone and its synthetic precursors[1,34-36]
HSD、NHP 和NRG 多存在于芸香科下属柑橘属植物的果皮中,以糖苷形式存在,是柑橘属中黄酮类化合物的主要成分。工业上多以NHP 为原料在碱性条件下开环得到NHDC。随着市场需求量的增加,直接从柑橘果实中提取NHP 已无法满足市场需求,现通常为化学合成所得[37]。合成的方式有2 种:a.生物法合成:以HSD 为原料,在生物酶的作用下将其转化为NHP;b.化学法合成:以NRG 为原料,通过催化剂催化羟醛缩合反应合成NHP,再进一步以常规加氢法或非常规加氢法得到NHDC[33]。
2 药理作用研究进展
在过去的时间里,研究人员开展了各种体内外实验来评价NHDC 及其合成前体的药理作用,包括抗氧化、抑菌、抗炎、保护肝脏和降脂等。
2.1 抗氧化作用
黄酮类化合物多具有较强的抗氧化作用,不仅能直接清除自由基,还能通过激活抗氧化酶[38]、抑制氧化酶[39]、缓解一氧化氮[40]、提高尿酸水平[41]等途径防止组织损伤,预防多种氧化应激损伤或其他机制对组织造成的损害[29]。NHDC、HSD、NHP 及NRG均为黄酮类化合物,具有良好的自由基清除活性。
NHDC 对体内外氧化损伤均具有保护作用。Hu 等[5]实验表明,NHDC 能够有效缓解CCl4诱导的氧化损伤,表现为显著恢复过氧化氢酶(catalase,CAT)、超氧化物歧化酶(superoxide dismutase,SOD)和谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-px)活性以及谷胱甘肽(glutathione,GSH)水平,下调CCl4诱导的核因子-κB(nuclear factor-kappa B,NF-κB)、白细胞介素6(interleukin 6,IL-6)、细胞凋亡蛋白酶caspase-3 和caspase-8 等表达。Choi 等[6]和Suarez 等[7]研究发现NHDC 对不同的活性自由基和氧自由基具有清除作用,对自由基、OH 自由基、非酶性脂质过氧化、H2O2和HOCl 的抑制作用分别为31.53%~84.62%、6.00%~23.49%、15.43%~95.33%、73.5%±0.9%和93.5%±2.9%;其中NHDC对H2O2和次氯酸(HOCl)抑制作用最佳。这些结果表明NHDC 是一种有效的抗氧化剂。
HSD 通过抑制氧化应激产生抗氧化作用。Aksu等[15]在黏菌素诱导的生殖损伤模型中发现HSD 显著降低睾丸氧化应激水平,如丙二醛(malondialdehyde,MDA)水平明显降低,GSH、GSH-px 水平明显升高,睾丸组织中caspase-3 的表达水平降低,但HSD组内无统计学差异。Raushan 等[42]研究发现HSD 在高脂血症模型中可减弱氧化还原稳态的改变。与高脂饮食(high-fat diet,HFD)组相比,HSD 组大鼠细胞内活性氧(reactive oxygen species,ROS)显著降低,MDA、蛋白羰基(protein carbonyl,PCO)和人晚期氧化蛋白产物(oxidation of plasma protein,AOPP)水平显著降低。Estruel 等[43]研究发现HSD 可以预防运动引起的氧化应激,提高运动能力,既防止了额外衰竭试验诱导的ROS 产生,又避免了胸腺和脾脏中SOD 和CAT 活性的降低。Elhelaly 等[44]发现HSD和地奥司明(diosmin,DS)联合使用显著降低了丙酰胺(acrylamide,AA)诱导的血清和组织的生化改变,改善了AA 暴露引起的炎症反应和氧化损伤。
NHP 对DPPH 自由基和ABTS+自由基有一定的清除能力,其半抑制浓度(the half maximal inhibitory concentration,IC50)分别为29.54 mg/mL 和22.54 μg/mL[16],贾富霞等[45]用最小二乘法回归方法,以总抗氧化活性指标结果进行回归分析,相对于柚皮苷、野漆树苷等成分,NHP 与DPPH 自由基、ABTS+自由基和总抗氧化能力正相关的药效组分相关性最强。
NRG 具有较强的体内外抗氧化活性,其体外具有超氧化物自由基清除活性、黄嘌呤氧化酶抑制活性、抑制ROS、抑制脂质过氧化等作用,特别在活性氧相关疾病中发挥重要作用。龙江宜等[46]研究发现NRG 通过降低CC 类趋化因子配体2(chemokine CC motif ligand 2,CCl2)的海马氧化应激反应,抑制caspase-8,caspase-3 信号凋亡通路,下调相关凋亡基因,对抗CCl2引起的神经元结构损伤,改善CCl2诱导的学习记忆损害和认知障碍。Chen 等[17]研究表明,NRG 能够通过调节Nrf2 信号通路,抑制长期氧化应激导致的抗氧化剂消耗,用以维持肺部氧化系统平衡。NHDC 及其合成前体抗氧化通路作用机制如图2 所示。
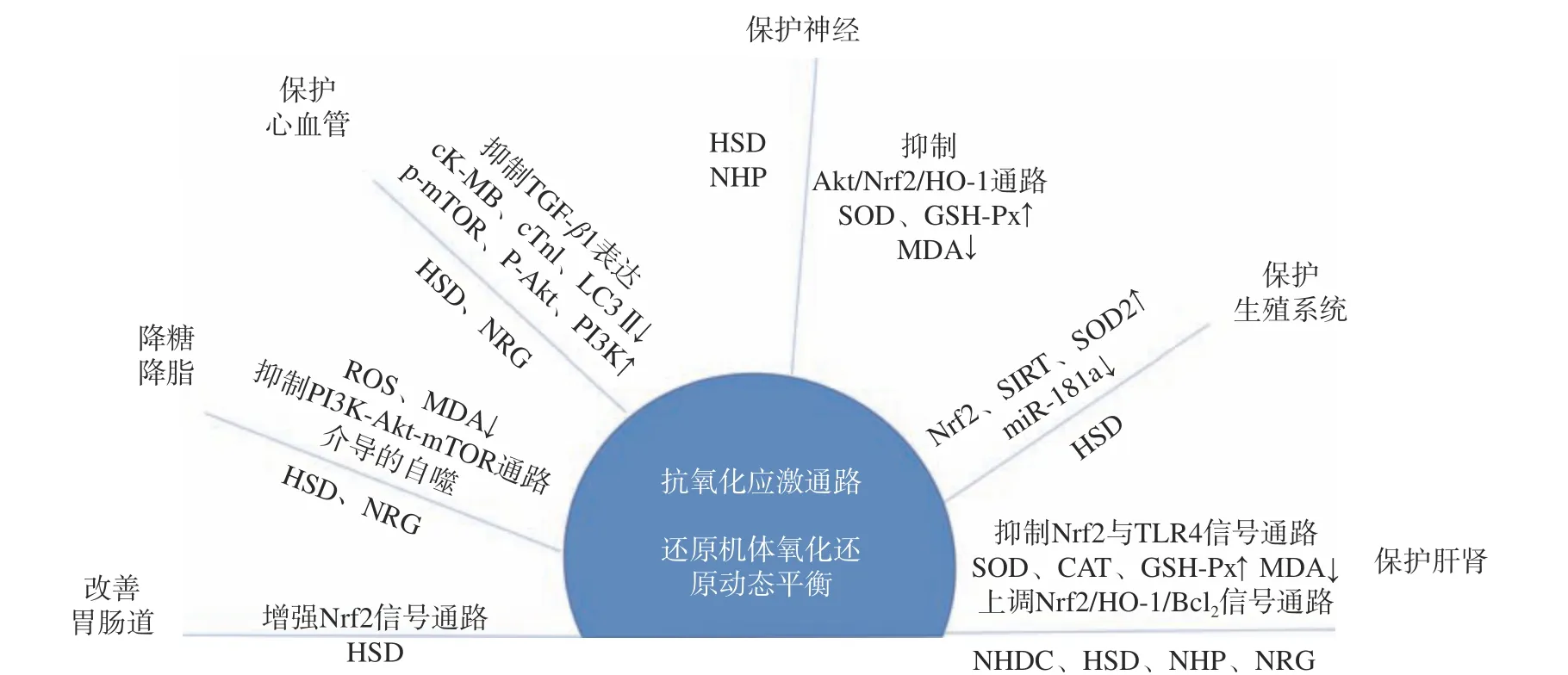
图2 NHDC 及其合成前体抗氧化通路作用机制[5-7,15-17]Fig.2 Action mechanism of NHDC and its synthetic precursors in the antioxidant pathway[5-7,15-17]
2.2 抑菌作用
NHDC 及其合成前体均为黄酮类化合物,具有抗有害微生物的作用。常见的抗菌机制包括:抑制核酸合成、影响生物膜形成而抑制细胞质功能和影响某些关键酶的相互作用等[46]。HSD 抑菌效果无种属特异性,具有广谱的抑菌活性,可作为广谱抑菌药物进行开发。张启焕等[47]和Jin 等[48]采用平板法和琼脂稀释法检测抑菌活性,结果表明HSD 可以抑制青霉菌生长半径和孢子形成,对六种菌(大肠杆菌、金黄色葡萄球菌、伤寒沙门氏菌、阴沟肠杆菌、粪肠球菌、表皮葡萄球菌)均具有抑制作用,效果随浓度的增加而增加。
Du 等[49]用TLIM 脂肪酶对毛孢热霉进行抑菌活性研究,试验结果表明,当NHP 浓度为50 mg/mL时,生长抑制区为24.2 mm;当NHDC 浓度为50 mg/mL 时,生长抑制区为26.3 mm,对抑制革兰氏阴性菌有很大的改善作用,但抑菌作用仍不如新橙皮苷酯衍生物,可能与衍生物较高的亲脂性相关。左龙亚等[50]采用菌丝生长速率法研究柠檬果皮中多酚物质抑菌活性,表明NHP 为柠檬果皮提取物抑菌性检测中最佳作用因子之一,相关系数为0.869。Zhao等[51]研究表明NRG 能显著提高细菌(大肠杆菌和金黄色葡萄球菌)的氧化应激,有效地抑制假单胞菌的生物膜形成,导致细菌RNA 与DNA 的泄露,并对牙周病原菌和常见的口腔微生物也具有一定的抑制作用。
2.3 抗炎作用
炎症是血管组织对有害刺激的复杂生物反应的一部分,如存在病原体、受损细胞或刺激物[52]。尽管炎症是对组织损伤的正常反应,若不加以控制,可能会导致慢性自身免疫性疾病[53]。天然植物中HSD、NRG 和NHP 等黄酮类化合物,已被证明具有良好的抗炎效果。Shi 等[4]发现NHDC 对百草枯(paraquat,pq)所致小鼠急性肝损伤具有很强的抗炎作用,表现为显著抑制pq 诱导的NF-κB 表达和线粒体驱动的凋亡信号,对肝损伤起到有效的保护作用。
HSD 具有抗炎作用,在化学性炎症诱导模型、慢性阻塞性肺疾病(chronic obstructive pulmonary disease,COPD)模型中得到验证,抗炎作用与降低炎症因子过表达等相关。Li 等[18]首次发现HSD 能降低皮肤组织中环氧合酶-2(cyclooxygenase-2,COX-2)的过表达和炎症细胞的浸润,对RAW264.7 细胞的迁移有剂量依赖性的抑制作用。Wang 等[54]研究表明,HSD 能有效减轻COPD 小鼠肺组织炎症反应和细胞死亡,HSD 高剂量组小鼠支气管肺泡灌洗液(bronchoalveolar lavage fluid,BLAF)中IL-6、白细胞介素8(interleukin8,IL-8)水平及肺髓过氧化物酶(myeloperoxidase,MPO)活性均有效下降,肺组织中沉默信息调节因子(silence information regulator 1,SIRT1)、过氧化物酶体增殖物激活受体γ辅激活子1α(peroxisome proliferator-activated receptor-γcoactivator-1α,PGC-1α)、磷酸化p-65(p-p65)的表达水平提高,为HSD 治疗COPD 提供了研究依据。
NHP 在大多炎症模型中具有良好的抗炎作用,如对皮肤炎症表现出良好的抑制作用,具有一定的抗过敏开发价值[55]。但Hamdan 等[56]研究发现NHP 与吲哚美辛联用能显著加重消炎痛给药后胃的损伤,表现为胃溃疡指数升高和胃组织病理改变,加重了溃疡的形成,具有促炎症特性,其部分原因可能是诱导大鼠胃细胞DNA 断裂,并增加了COX-2 的表达,导致胃损伤加重。因此,最好在炎症条件下避免NHP 与吲哚美辛联用,避免溃疡恶化。
NRG 在体内外均可发挥抗炎作用。Ahmad 等[57]研究表明,NRG 能够抑制肿瘤坏死因子(tumor necrosis factor-α,TNF-α)和IL-1β的产生,显著上调转化生长因子β1(transforming growth factor-β1,TGFβ1)的mRNA 表达水平,有效调节抗炎细胞因子加速肺部炎症的修复过程。Gil 等[58]研究表明,NRG通过AMPK、p38 和Nrf-2 信号通路诱导巨噬细胞HO-1 表达,从而在脓毒症中产生抗炎活性。
2.4 保护肝肾作用
NHDC 可通过抗氧化、抗炎和抗凋亡等作用减轻肝脏损伤。Xia 等[8]发现NHDC 通过维持肝功能、减少ROS 生成和减少促炎细胞因子分泌来有效预防脂多糖(Lipopolysaccharide,LPS)诱导的暴发性肝衰竭(fulminant hepatic failure,FHF)。Shi 等[4]研究表明,NHDC 能有效减轻pq 诱导的急性肝损伤,通过显著逆转pq 诱导的促凋亡相关蛋白(Bcl-2-associated X,Bax)上调和B 细胞淋巴瘤/白血病-2(B-cell lymphoma/leukemia-2,Bcl-2)下调,抑制pq增加,减轻氧化应激状态,阻断NF-κB 信号级联,减少促炎症介质生成的增加,减轻肝损伤。Su 等[59]研究表明,NHDC 通过直接清除自由基和间接清除Nrf2/ARE 信号通路激活,对抗CCl4诱导的肝损伤。
HSD 对肝肾损伤具有保护作用,与抗氧化、抗炎和抗凋亡等密切相关。Ramadan 等[26]研究表明,HSD 通过上调Nrf2/HO-1/Bcl2 信号通路和下调NF-κB,降低促炎症和凋亡介质,提高抗氧化能力,保护肝脏免受甲氨蝶呤(Methotrexate,MTX)诱导的氧化应激、脂质过氧化、炎症和凋亡的影响。ABD-Elhakim等[60]利用CCl4诱导的急性肝损伤模型研究发现HSD 单独或与牛磺酸联合使用能使谷丙转氨酶(alanine aminotransferase,ALT)、天冬氨酸转氨酶(aspartate aminotransferase,AST)、碱性磷酸转氨酶(alkaline phosphatase,ALP)、γ-谷氨酰转肽酶(γgltamyltranspeptidase,GGT)等水平显著降低,白蛋白与白蛋白/球蛋白比值分别显著增高,改善肝功能。隗世波等[61]证实了HSD 干预对脓毒症所致急性肾损伤(acute kidney injury,AKI)大鼠肾脏有一定的保护作用,可以改善脓毒症所致肾脏病理和肾功能损伤。Park 等[62]观察到HSD 对缺血致AKI 大鼠的肾功能具有保护作用,与模型组相比,HSD 处理组CAT、SOD、GSH-px 和一氧化氮的表达量较高,而尿素氮和血清肌酐的表达量较低,故能够通过抗氧化作用改善AKI。
NHP 可以减少脂肪酸氧化导致的肝脏脂肪变性。Wang 等[63]检测了肝脏中参与脂肪酸合成和脂肪酸氧化的关键基因的表达,结果表明,NHP 上调了过氧化物酶体增殖物激活受体(proliferator-activated receptorα,PParα)、乙酰辅酶A 酰基转移酶2(acetylcoenzyme aacyltrans-ferase 2,Acaa2)、肉毒碱棕榈酰转移酶-1(carnitine palmitoyl transferase1,Cpt-1)、丙酮酸脱氢酶激酶4(pyruvate dehydrogenase kinase 4,Pdk4)和过氧化物酶体酰基辅酶A 氧化酶1 蛋白(acyl-coenzyme A oxidase 1,Acox1)等HFD 小鼠肝脏中脂肪酸氧化基因的表达,同时下调了固醇调控元件结合转录因子1(sterol regulatory elemental binding transcription factors,Srebf1)、脂肪酸合成酶基因(fatty acid synthase,Fasn)、硬脂酰辅酶A 去饱和酶1(stearoyl-CoA desaturease1,Scd1)和乙酰辅酶A 羟化酶1(acetyl coenzyme A carboxylase 1,Acc1)等脂肪生成基因的表达,最终达到改善HFD 小鼠的肝脏脂肪变性。
NRG 具有抗炎和清除自由基的作用,可防止肝星状细胞的激活和肝纤维化,对乙酰氨基酚、阿霉素、环磷酰胺、顺铂等化学药物或毒素引起的肝肾损伤具有一定的保护作用,以不同的途径抑制肝损伤[36]。如Caglayan 等[27]发现NRG 预处理组显著降低了血清毒性指标,提高了抗氧化酶活性,通过调节氧化应激、炎症、凋亡、自噬和DNA 损伤来保护环磷酰胺诱导的肝毒性和肾毒性。Adil 等[64]证实NRG 对扑热息痛所致大鼠肝肾毒性具有一定改善作用,表现为NRG 预处理组可维持异常抗氧化和活性氧生成,对抗细胞损伤,增强法尼醇X 受体(farnesoid X receptor,FXR)和尿肾损伤因子-1(kidney injury molecule-1,KIM-1)mRNA 的表达。NHDC 及其合成前体保护肝肾作用机制如图3 所示。
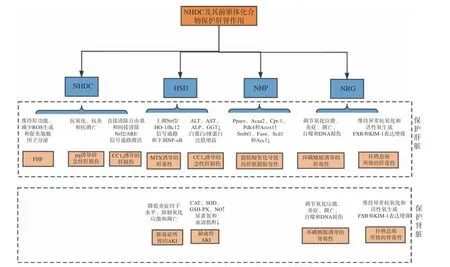
图3 NHDC 及其合成前体保护肝肾作用机制[4,8,26-27]Fig.3 Action mechanism of hepatorenal protective effect of NHDC and its synthetic precursors[4,8,26-27]
2.5 降血糖、降血脂作用
肥胖的发病机制复杂,与多种因素有关。NHDC在高血脂模型中表现出降血脂作用。Han 等[28]实验表明,分化组的油红O 染色在所有组中最高,NHDC组油红O 染色较分化组略少。Bok 等[9,65]实验表明NHDC 可以大大降低血浆胆固醇水平,抑制HMGCOA 还原酶和酰基辅酶A-胆固醇酰基转移酶(acyl coenzyme A-cholesterol acyltransferase,ACAT)的活性,防止动脉内皮细胞中巨噬细胞-脂质复合物的积累,抑制脂肪肝形成;此外,NHDC 能显著提高动物血浆高密度脂蛋白(high-density lipoprotein,HDL)的含量,从而促进胆固醇的代谢。
HSD 通过调节脂质代谢和糖代谢发挥降糖降脂作用[66]。Rehman 等[20]研究表明,HSD 可以通过改善HFD 引起的血脂水平异常,提高炎症介质(瘦素、IL-6 和TNF-α)的血清水平等改善胰岛素抵抗。-Sundaram 等[67]发现HSD 通过调节链脲佐菌素诱导的糖尿病大鼠碳水化合物代谢的糖原合酶和糖原磷酸化酶来改善高血糖。Prasatthong 等[68]在HFD 导致的代谢综合征(metabolic syndrome,MS)中发现,HSD和二甲双胍联合显著减轻了脂肪垫和脂肪肝的堆积,扭转了脂肪细胞的大小和数量,改善了MS 大鼠的心肌肥厚,使胰岛素信号通路IRS/Akt/GLUT4 蛋白表达恢复。
NHP 具有降血脂和降血糖的双重作用。Jia 等[21]研究表明,在糖尿病KK-Ay 小鼠模型中,NHP 能够降低空腹血糖、甘油三酯、瘦素等水平,抑制肝脏脂质积聚和附睾脂肪细胞增大,降低硬脂酰辅酶A 去饱和酶1(stearoyl-CoA desaturase 1,SCD1)、FAS、酰基辅酶A 氧化酶(acyl-coenzyme A oxidases,ACOX)等相关基因表达,从而发挥降糖降脂作用。Zhang 等[69]发现NHP 能够增加HepG2 细胞葡萄糖的消耗,呈剂量依赖性,且与NRG 联合使用比单独使用产生更多的葡萄糖,增强了AMPK 蛋白在细胞中的表达,可能通过糖代谢发挥潜在降糖作用。Wang 等[63]证实了NHP 能够通过促进PGC-1α的表达,增加线粒体生物发生,减轻HFD 小鼠的肝脏脂肪变性,减轻胰岛素抵抗。Lu 等[24]发现NHP 可通过改变HFD 小鼠肠道微生物群的组成,维持肠上皮的完整性,减轻肥胖引起的代谢紊乱,显著恢复肠道屏障损伤、代谢性内毒素血症和全身炎症。
NRG 通过改善内皮细胞功能障碍与肾纤维化发挥预防和治疗糖尿病的潜力,作用机制涉及抑制PI3K-Akt-mTOR 通路介导的自噬[22],抑制HG 诱导的ERK1/2 和JNK MAPK 磷酸化,抑制ERK1/2 和JNK MAPK 信号通路[70]。此外,Bi 等[25]发现NRG通过抑制炎症、凋亡和MAPK 通路,对LPS 诱导的HUVECs 细胞损伤有强大的拮抗作用。Pengnet 等[71]首次发现NRG 可有效抑制高胆固醇血症大鼠主动脉低密度脂蛋白受体-1(lectin-like oxidized lowdensity lipoprotein receptor-1,LOX-1)的表达,下调烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)氧化酶水平,从而降低氧化应激,改善高胆固醇血症诱导的内皮功能障碍,降低高胆固醇血症的致动脉粥样硬化风险。
2.6 改善胃肠道作用
胃肠道中,胃酸分泌和胃运动受5-羟色胺(serotonin,5-HT)调节,两者都是调节食物摄入和饱腹感的复杂网络机制的一部分。Zopun 等[72]实验表明,NHDC 是HGT-1 细胞通过甜味受体T1R3 释放5-HT 的有效诱导剂,但不可调控SLC2A1mRNA 表达水平的影响,其对HGT-1 细胞甜味感知相关基因的调控有待进一步研究。此外,NHDC 能够通过影响肠道微生物减少肠道疾病。Daly 等[73]实验表明,膳食中添加NHDC 可以积极影响乳酸杆菌的数量,改变胃肠道微生物群,增强免疫保护,减少胃肠道炎症反应,维持肠道健康。Shi 等[74]通过宏基因组学、代谢组学和生物学分析的高度创新组合评估糖精、非热量甜味剂(noncaloric artificial sweeteners,NAS)和NHDC 摄入对肠道微生物群的影响。三者相比,NHDC 对细菌群落没有显著影响。
HSD 对肠道中胃肠道传递功能、肠道微生物及肠道炎症有一定的改善作用。Wu 等[75]发现HSD通过5-羟色胺4R/cAMP 信号通路改善洛哌胺诱导的便秘大鼠结肠运动;与模型组相比,HSD 处理组粪便数量和含水量显著增高,肠道转运率升高;5-HT 受体荧光强度和细胞内游离钙离子增加,cAMP/PKA通路和磷酸化cAMP 反应成分结合蛋白(p-CREB)表达上调。Estruel 等[76]证实了HSD 能够通过增加肠道菌群总数来影响肠道微生物群,增加乳酸菌属和双歧杆菌属的比例,以较低剂量增加肠道免疫球蛋白A 来维持肠道内稳态,具有潜在益生元作用。Guo等[77]研究发现HSD 可通过增强Nrf2 抗氧化途径,增加调节性T 细胞(regulatory cell,Treg)种群数量,减少结肠组织损伤,恢复肠道屏障功能来保护肠道炎症、保护肠上皮细胞的存活和屏障的完整性。
NHP 可通过改变肠道微生物群防止直肠肿瘤的发生。Gong 等[78]首次揭示了肠道微生物群在NHP预防肿瘤方面起着重要作用,NHP 通过诱导肿瘤细胞凋亡和抑制血管生成来显著抑制APC Min/+小鼠结直肠肿瘤的发生。袁菱等[79]比较NRG、HSD、NHP分别和芍药苷配伍后相应成分的肠道吸收特点发现,单体成分与配伍成分结合使用对肠道吸收无显著性差异,配伍并不能显著性影响主成分的肠吸收。Lee 等[80]研究表明,枳实中成分NHP 具有抗胃溃疡和胃炎作用,显著降低了胃液分泌量和胃酸排出量,提高了胃酸pH,对HCl/乙醇诱导的胃损伤有55.0%的抑制作用,有望对胃炎起到保护作用。
NRG 对肠道血管屏障损伤具有保护作用。Liu等[81]研究表明,NRG 能够显著抑制TNF-α刺激下RIMVECs 细胞的迁移,使肠血管屏障(gut-vascular barrier,GVB)内皮细胞完整性升高,保护RIMVECs单层屏障的完整性,紧密连接蛋白zona 10 occludin-1、occludin、claudin-1 和claudin-2 的表达和分布。
2.7 心血管保护作用
已有研究表明HSD 能够抑制肾血管性高血压大鼠肾素-血管紧张素系统,具有降压作用。在此基础上,Maneesai 等[82]发现HSD 能够抑制L-精氨酸甲酯(L-arginine methyl ester,L-NAME)诱导的大鼠高血压的发生和心血管重构,抑制TGF-β1 和基质金属蛋白酶(matrix metalloproteinases,MMPs)蛋白表达,对心血管具有保护作用,其机制与减少氧化应激和炎症作用有关。Li 等[83]研究发现HSD 预处理组可显著降低心肌梗死面积、心肌损伤及血清肌酸激酶同工酶(cK-MB)、肌钙蛋白Ⅰ(cTnI)水平,显著下调了微管相关轻链蛋白3Ⅱ(LC3Ⅱ)和自噬基因Beclin1的表达,显著上调了磷酸化雷帕霉素靶蛋白(phosphorylated mammalian target of rapamycin,pmTOR)、磷酸化蛋白激酶B(phosphorylated protein kinase B,P-Akt)和磷脂酰肌醇3-激酶(phosphatidylinositol 3-kinase,PI3K)的表达,通过抑制过度自噬具有保护心肌缺血/再灌注损伤作用。Kuzu 等[84]发现HSD 处理组可显著抑制MDA 和8-羟化脱氧鸟苷(8-hydroxy2′-deoxyguanosine,8-OHdG)的增加,抑制GSH 水平的降低,提高SOD、CAT、GSH-px等抗氧化酶活性。通过减少氧化应激和细胞凋亡,增加抗氧化酶活性,预防炎症,保护心脏细胞结构。
NHP 可能是治疗心肌肥厚和心脏重塑的潜在药物。Zhang 等[85]研究发现,NHP 能显著抑制血管紧张素Ⅱ抗体(angiotensin II,AngII)诱导的心肌收缩功能障碍、心肌肥厚、心肌纤维化、心肌氧化应激和炎症反应,减轻Ang II 引起的心肌肥大和重构。
NRG 可减少心肌细胞的凋亡。Chen 等[86]研究表明,NRG 能够通过增加线粒体降低的GSH 活性和抑制H9c2 细胞中MAPK(p38 MAPK,ERK1/2和JNK)的激活和氧化应激来保护HG 诱导的损伤。刘丹等[87]发现NRG 预处理可降低Caspase-3 活性,下调IRE1α蛋白表达水平,抑制IRE1α途径,对心肌缺氧/复养(hypoxia/reoxygenation,H/R)损伤有保护作用,从而减少内质网(endoplasmic reticulum stress,ER)应激或ER 应激介导的细胞凋亡。
2.8 神经损伤保护作用
HSD 对各种神经退行性疾病如阿尔茨海默氏病、帕金森病、中风、亨廷顿病等发展具有抑制作用,其神经保护潜力是通过改善神经生长因子和内源性抗氧化防御功能,减少神经炎症和凋亡途径介导的[88]。Hajialyani 等[89]通过评估ROS 生成、线粒体膜电位和细胞凋亡水平等发现,HSD 减轻鱼藤酮诱导的神经元损伤与降低氧化应激、改善线粒体功能障碍和细胞凋亡相关。Antunes,等[90]发现HSD通过调节C57BL/6 小鼠黑质致密部(substantia nigra pars compacta,SNpc)中的线粒体功能障碍、凋亡和多巴胺能神经元来防止6-羟多巴胺(6-hydroxydopamine,6-OHDA)注射引起的运动、嗅觉和空间记忆障碍,从而调节行为的改变。Welbat 等[91]证实HSD 既能改善MTX 诱导的成年大鼠氧化应激,也能有效预防MTX 诱导的细胞损伤和海马神经损伤。
NHP 具有活性氧清除和体外神经保护作用。Wang 等[92]研究表明,NHP 可以显著改善神经功能,减轻大脑中动脉闭塞(middle cerebral artery occlusion,MCAO)诱导的梗死体积、病理改变和神经元损失。通过抑制MCAO 诱导的Bax、细胞色素C、cleaved caspase-9 和-3 的上调以及Bcl-2 的下调,抑制凋亡。Wang 等[93]首次证实NHP 通过阻断蛋白二硫键异构酶(protein-disulphide isomerase,PDI)的s-亚硝基化,防止未折叠蛋白(unfolded protein response,UPR)和ER 应激相关的神经毒性,减弱Aβ25-35 诱导的细胞凋亡,对Aβ25-35 诱导的原代培养海马神经元神经毒性具有保护作用。
NRG 具有体内外神经保护作用。Kulasekaran 等[94]证明NRG 显著降低了3-硝基丙酸(3-nitropropionic acid,3-NP)诱导的活性氧和脂质过氧化水平,通过作用于PI-3K/Akt 依赖的Nrf2 信号通路,使PC12 细胞免受3-NP 诱导的神经毒性。Meng 等[95]发现NRG通过淀粉样β代谢、Tau 蛋白超磷酸化、氧化应激和细胞凋亡等多种机制,显著改善氢化可的松诱导的记忆障碍小鼠的认知、学习和记忆功能紊乱。
2.9 生殖系统保护作用
HSD 对睾丸损伤有保护作用。Helmy 等[96]研究表明,HSD 在邻苯二甲酸二(2-乙基己基)酯(Di-(2-ethylhexyl) phthalate,DEHP)诱导的睾丸损伤模型中,HSD 预处理组睾丸3β羟基固醇脱氢酶(3βhydroxysteroid dehydrogenases,3β-HSD)活性和血清睾酮水平显著升高,抗氧化应激水平显著升高,微小RNA-181a(microRNA-181a,miR-181a)表达水平降低,SIRT1 和超氧化物歧化酶2(SOD2)的蛋白表达水平显著上调,Nrf2 和HO-1 蛋白表达水平升高,睾丸器官系数趋于正常。Olayinka 等[97]在非那雄胺诱导的睾丸损伤模型中发现,HSD 处理组通过抑制氧化应激指标、改善精子参数和标记酶活性,抑制非那雄胺与5α-还原酶(5α-reductase,5AR)的结合,改善非那雄胺诱导的睾丸损伤。ABD-Elhakim 等[98]研究表明,HSD 与牛磺酸联合使用可显著抑制CCl4诱导的尿素、尿酸、钾和促卵泡激素水平的升高,减轻了CCl4诱导的氧化应激和炎症,诱发睾丸结构的改变,对CCl4诱导的生殖损伤具有预防性的保护作用。
2.10 抗癌作用
在过去的十年里,人们对HSD 及其苷元橙皮素的抗癌活性进行了大量的研究。HSD 对结肠癌、肿瘤、胃癌、皮肤癌、舌癌、肺癌、肝癌、肾癌和乳腺癌等各种类型的癌细胞具有很强的抑制作用,通过不同的机制在不同组织中诱导肿瘤细胞凋亡[99]。如Tan等[12]试验中与单独接受NScLc 细胞移植的大鼠相比,HSD 治疗的NScLc 细胞移植大鼠的肿瘤体积明显更小,且增加miR-132 的表达,降低ZEB2 的表达,显著降低了NScLc 细胞的集落形成效率,促进NScLc 细胞的凋亡。Kongtawelert 等[100]研究表明,HSD 通过下调三阴性乳腺癌(triple-negative breast cancer,TNBC)中Akt 和NF-κB 信号通路来抑制程序性死亡配体1(programmed death ligand 1,PD-L1)的表达,从而抑制乳腺癌细胞的生长。
Xu 等[13]首次证实NHP 通过激活Bcl-2/bax 介导的信号通路诱导人乳腺腺癌MDA-MB-231 细胞凋亡,并显著上调肿瘤抑制蛋白P53 和Bax 的表达,下调Bcl-2 表达,导致Bax/Bcl-2 比值升高,与MDAMB-231 细胞凋亡的发生相对应。
NRG 能够抑制HeLa 宫颈癌细胞、人三阴性乳腺癌细胞、P388 细胞和H69AR SCLC 等恶性肿瘤的细胞生长;以H69AR SCLC 细胞和宫颈癌HeLa细胞为例,Chen 等[14]首次证实NRG 的抗癌作用为H69AR 细胞的抑制增殖和诱导凋亡,并上调miR-126 表达,抑制了血管细胞粘附分子-1(vascular cell adhesion molecule-1,VCAM-1)蛋白水平,抑制SCLC的生长。Zeng 等[101]证实了NRG 通过抑制NF-κB/COX-2-caspase-1 通路的激活,抑制细胞生长并诱导其凋亡,显示抗癌作用。
2.11 其他作用
NHDC 及其合成前体主要药理作用如图4 所示,具体作用机制及途径总结见表1 所示。除此之外,NHDC 也是哺乳动物、细菌和真菌α-淀粉酶的激活剂,具有不同作用机制,可以稳定细菌的淀粉酶,但不能防止热诱导的展开[102]。

表1 NHDC 及其合成前体药理作用及机制Table 1 Pharmacological effect and mechanism of NHDC and its synthetic precursors
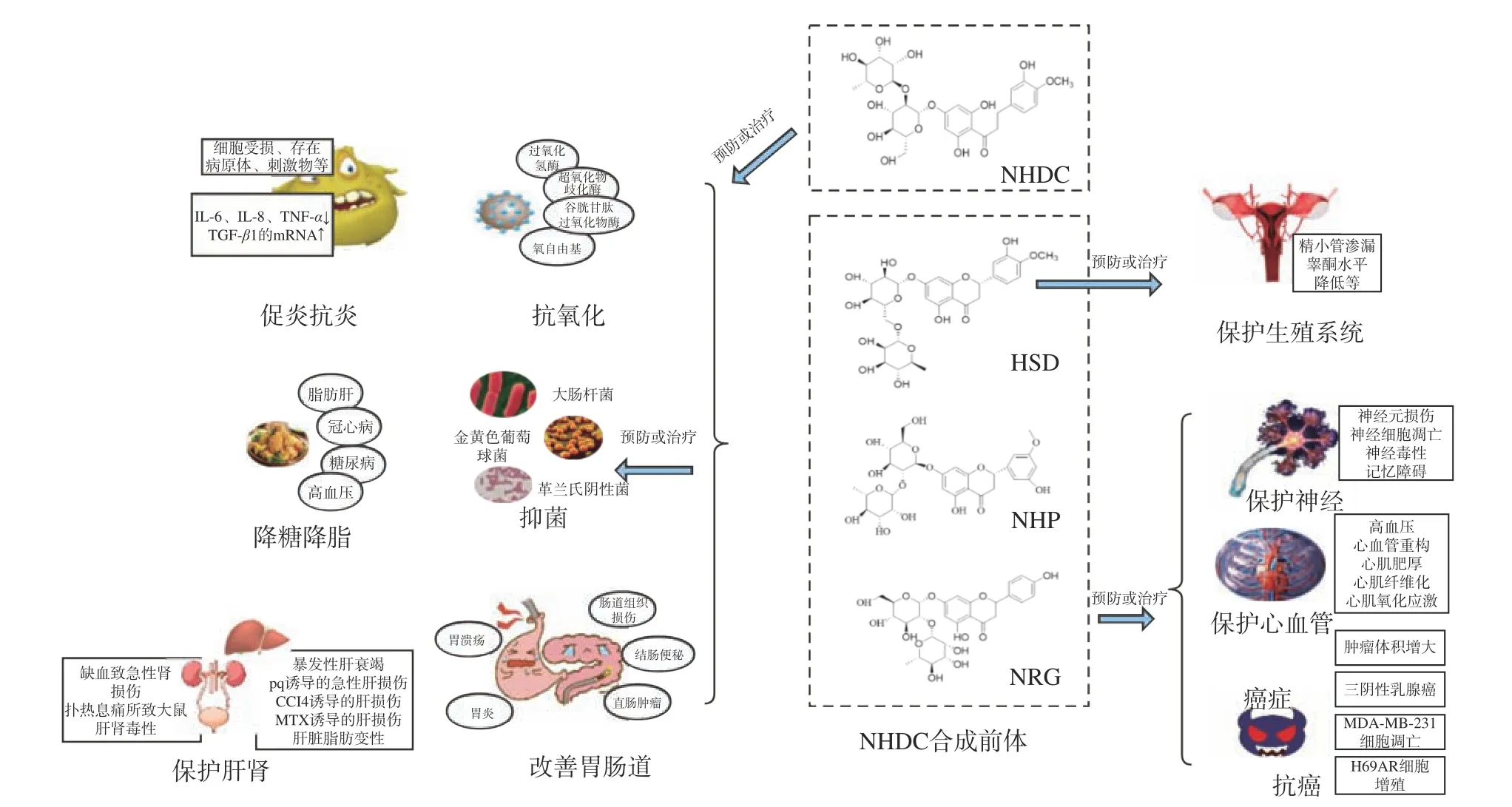
图4 NHDC 及其合成前体药理作用对应疾病[4-8,15,21-27,46-49,54-66,73-96]Fig.4 Pharmacological effects of NHDC and its synthetic precursors correspond to diseases[4-8,15,21-27,46-49,54-66,73-96]
HSD 具有广泛的生物学活性,除以上主要的药理作用以外,还具有抗衰老、镇痛等作用。Lee 等[103]研究发现HSD 通过抑制MAPK 依赖的信号通路,调节MMP-9 的表达,抑制小鼠皮肤厚度增加、皱纹形成和胶原纤维丢失。Ventura 等[104]在小鼠内脏疼痛模型中研究发现,HSD 与安乃近(metamizole)协同使用具有镇痛作用,54.4 mg/kg 的安乃近与42.2 mg/kg 的HSD 组合使用具有70%的镇痛作用,与单独服用100 mg/kg 的安乃近或100 mg/kg 的HSD 镇痛效果接近。
NHP 还具有减轻肺纤维化、抗衰老、溃疡愈合等作用。Guo 等[105]研究表明,NHP 主要通过抑制TGF-β1/Smad2/3 通路、阻断TGF-β1 信号通路,特别是抑制Smad3 激活,参与保护上皮细胞损伤、抑制肌成纤维细胞分化、ECM 沉积和成纤维细胞迁移。Tan 等[106]发现NHP 可通过影响RANKL/RANK-和Ca2+依赖的信号通路抑制破骨细胞分化;通过抑制TRAcP 和组织蛋白酶k 的表达抑制破骨细胞介导的骨吸收等,为骨质疏松症提供了一种潜在的治疗方法。Guo 等[107]研究表明,NHP 以浓度依赖性的方式延长了酵母的实生寿命(CLS)。在降低酵母ROS 积累方面表现出最强的功能,属于抗衰老柑橘类黄酮。
NRG 还具有体内外抗骨质疏松和支气管扩张作用,与可促进破骨细胞凋亡,抑制破骨细胞吸收,有效的增强骨密度、骨小梁厚度等相关[108]。Li 等[109]评估NRG 对骨吸收影响,结果表明NRG 有效抑制OVX诱导的骨丢失,增加骨密度、骨体积和骨小梁厚度,促进成骨细胞分化,有效逆转卵巢切除相关骨质疏松症。Yue 等[110]研究发现NRG 是有显著作用的支气管扩张剂。在体外能使培养的气道平滑肌ASMC 细胞放松,在体内能使小鼠收缩的支气管气道扩张,是一种有效、安全的哮喘扩张剂候选药物。
3 结论与展望
通过检索整理文献发现,NHDC 合成前体发展历程久,研究领域涉及各个系统当中,作用机制的研究包括细胞层次、动物层次和临床相关报道,结合体内外研究广泛应用于临床当中。相比之下,NHDC药理作用有待于进一步研究,当前从氧化应激、肝肾损伤、血脂紊乱和胃肠道疾病等方面对NHDC 的研究居多,特别是氧化应激方面。NHDC 体外研究表明具有HOCl 清除活性,清除能力高于抗坏血酸,对黄嘌呤氧化酶、非酶脂质过氧化有重要的抑制作用,是一种强抗氧化剂[6-7]。体内研究表明,NHDC 的抗氧化特性可能对pq 诱导的小鼠毒性有显著肝保护作用,其清除自由基能力对CCl4诱导的急性氧化损伤也有肝保护作用,有可能被用作肝保护剂[4-5,59]。NHDC 的甜味、低毒、强抗氧化特性使其在药物开发方面具有许多优势,之后需要进一步的研究来阐明NHDC 在抗氧化通路的更多药理作用及相关机制。NHDC 作为甜味剂,多应用于饲料及食品行业,但目前在药物开发方面相比其合成前体仍存在一些问题:a.药理研究涉及领域较窄,现多集中于抗氧化、抗炎等研究方向,未全面向各系统开展药理作用研究,更缺乏临床相关报道;b.实验研究不够深入,其靶点和作用通路尚未阐明,缺乏体内、体外相结合的实验,缺少有力的证据。总之,NHDC 在药物开发方面相比HSD、NRG 和NHP 等合成前体还不够完善,相关药理作用探索还需继续深入研究。本文多方位对三个合成前体的药理作用及机制进行探讨,后续可参考其合成前体研究领域,对NHDC 加大研究力度,借助现代研究手段,挖掘成分新活性或现有活性新靶点,为NHDC 的开发利用提供科学依据,使其能够广泛应用于临床疾病治疗当中。
