大肠杆菌发酵生产L-苯丙氨酸新工艺研究
2022-12-05陈志超李澜潇徐庆阳
陈志超,李澜潇,徐庆阳
(天津科技大学 生物工程学院,天津 300450)
L-苯丙氨酸(L-phenylalanine,L-phe)是人体8种必需氨基酸之一[1],在食品和制药工业中有诸多应用,是新型甜味剂阿斯巴甜的重要前体物质[2],也是部分风味物质的鲜味物质[3],还可合成调味剂苯乙醇[4],后者广泛应用于乳制品与调味品中,在医药方面主要作为抗癌药物的载体[5-6]。常用的生产L-苯丙氨酸的方法为酶法与微生物发酵法[7],目前工业化生产L-phe主要采用具有原料廉价易得、环境污染小、产物纯度高等优点的微生物发酵法[8-9]。
L-phe长势与产量联系密切,是一种与菌体生长部分耦联的产物[10]。维生素H可在脂肪酸生物合成、氨基酸代谢和糖异生的羧化反应中作为酶促辅因子,能够激活相应酶的羧基转移亚基的CO2[11-13];维生素H用量一直是微生物发酵生产的重要参数[14-15]。在大肠杆菌中,过量维生素H会导致菌体代谢旺盛、生长过快,出现糖酸转化率大幅降低的问题;过少或者不添加维生素H会导致菌体生长较慢、生物量较低、产酸效率偏低等问题。氨基酸发酵生产常采用分批补料发酵,发酵培养基一次性加入罐中,过程流加葡萄糖等作为碳源,该工艺势必会导致菌体在发酵前期处于营养物质过饱和的状态[16],造成菌体营养中毒,也会造成营养物质的浪费和代谢流的改变[17]。
为解决上述问题,本研究对发酵培养基中维生素H的浓度进行了优化,旨在葡萄糖亚适量控制工艺[18]前提下,采用全营养流加工艺,即:将发酵培养基按一定比例分为底物培养基和流加培养基,底物培养基供菌体初期生长,流加培养基在发酵过程中按需流加,最终实现了生物量、L-phe产量、糖酸转化率的同步提高,为大肠杆菌发酵生产L-phe提供了新的思路。
1 材料与方法
1.1 菌株
L-phe大肠杆菌生产菌:由天津科技大学代谢工程实验室保存并提供。
1.2 培养基
种子培养基:葡萄糖30 g/L,酵母粉4 g/L,蛋白胨2 g/L,柠檬酸1.5 g/L,MgSO4·7H2O 1.5 g/L,KH2PO42.0 g/L,硫酸铵2.0 g/L,维生素B11 mg/L,FeSO4·7H2O 10 mg/L,MnSO4·H2O 5 mg/L,维生素H 1 mg/L,酪氨酸1.5 g/L,卡那霉素20 g/L。
发酵培养基:葡萄糖20 g/L,MgSO4·7H2O 2 g/L,酵母粉4.5 g/L,蛋白胨1.5 g/L,硫酸铵2 g/L,柠檬酸1.5 g/L,K2HPO4·3H2O 6~10 g/L,酪氨酸1.2 g/L,谷氨酸1.2 g/L,维生素H 0~2 mg/L,FeSO4·7H2O 20 mg/L,MnSO410 mg/L,维生素B1、B3、B5、B12各2 mg/L,微量元素混合液2 mL/L[19],卡那霉素10 mg/L。
1.3 主要试剂与仪器
葡萄糖:食品级;磷酸二氢钾、酪氨酸、柠檬酸、硫酸铵、维生素H、谷氨酸:均为分析纯。
LDZH-100KBS型全自动立式蒸汽灭菌器 天津博鑫生物科技有限公司;5 L自动控制发酵罐 上海保兴生物设备工程有限公司;SBA-40E生物传感分析仪 山东省科学院生物研究所;Agilent 1200高效液相色谱仪 Agilent Technologies公司;KQ-C高压蒸汽发生器 上海奉贤协新机电厂;723分光光度计 上海菁华科技仪器有限公司;Olympus生物显微镜 日本Olympus株式会社;尾气分析仪 美国诺瓦生物医学公司;S-433D氨基酸分析仪 德国赛卡姆公司。
1.4 培养方法
1.4.1 菌种活化
取甘油保菌管中菌株置于斜面培养基上,于37 ℃倒置恒温培养12 h,传代2次。
1.4.2 5 L罐种子培养
初始发酵液体积为3 L,发酵温度为37 ℃,初始通气量为1.6 L/min,初始搅拌转速为200 r/min,后期调节通气量与转速维持DO(溶氧值)在50%左右,通过流加25%氨水调节发酵液pH为6.8~7.0,OD600 nm>25时接种发酵。
1.4.3 5 L罐分批补料发酵培养
接种量为20%~30%,发酵温度为37 ℃,初始通气量为0.65 L/min,初始搅拌转速为200 r/min,后期调节通气量与转速,维持DO在35%~50%,通过流加25%氨水控制发酵液pH为6.8~7.0,当初始培养基中葡萄糖耗尽(发酵4~7 h) 时,流加800 g/L的葡萄糖,控制糖浓度在0~0.5 g/L,并随糖液流加菌株缺陷物质酪氨酸,每升糖液混合1 g酪氨酸,酪氨酸利用NaOH溶液溶解后单独灭菌,待糖液与酪氨酸溶氧均冷却后混匀,发酵50 h。
1.4.4 5 L罐全营养流加发酵培养
发酵控制工艺与1.4.3相同。将发酵培养基按实验设计方案分成底物培养基与流加培养基,在发酵过程中按需补充菌体所需营养物质。
1.5 检测与分析方法
1.5.1 pH、DO、温度的测定[20]
pH、DO、温度均采用梅特勒电极在线实时检测,并配合精密pH试纸(6.4~8.0)人工校正pH。
1.5.2 菌体生物量的测定[21]
每隔2 h取样,适当稀释至吸光值可测浓度范围(0.2~0.9),用紫外可见分光光度计测量样品在600 nm处的吸光值。
1.5.3 L-phe及副产物的测定
发酵液中L-phe产量采用LC-20AT高效液相色谱仪测定,乙酸的检测参考文献[22],谷氨酸采用SBA生物传感分析仪测定。
1.5.4 残糖的测定及耗糖速率、转化率的计算
发酵液残糖浓度利用SBA生物传感分析仪测定。利用天平检测耗糖量,并以下式计算耗糖速率。
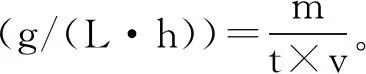
式中:m为单位时间耗糖量,g;t为时间,h;v为发酵液体积,L。
糖酸转化率的计算方法参照文献[23]。
2 结果与分析
2.1 维生素H浓度对L-phe发酵的影响
本部分实验采用分批补料发酵工艺,在发酵培养基的基础上分别额外添加0,0.5,1.0,2.0 mg/L维生素H,并以添加0 mg/L维生素H的发酵组作为对照,探究维生素H对大肠杆菌发酵生产L-phe的影响。
2.1.1 维生素H浓度对菌体生长与L-phe产量的影响
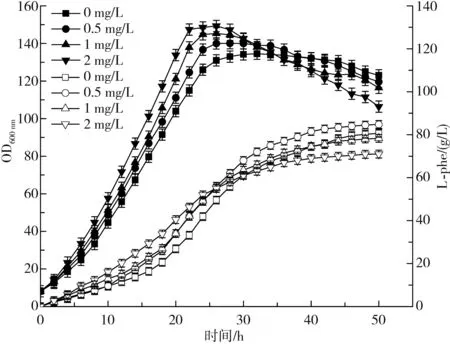
图1 不同浓度维生素H对菌体生长及L-phe产量的影响Fig.1 Effects of different concentrations of vitamin H on the bacteria growth and L-phe yield
由图1可知,维生素H浓度越高,菌体在发酵前期的生长速率越快,最大生物量越大,达到菌体稳定期的时间越短,菌体量稳定期维持时间越短,发酵后期菌体衰亡速度越快,放罐时生物量越少;L-phe产量与菌体量变化趋势略有不同,发酵25 h前,随着维生素H浓度的增加,L-phe积累速度随之加快,25 h后,各组L-phe积累速度均下降,0,0.5 mg/L维生素H添加组下降缓慢,L-phe持续积累,1,2 mg/L添加组下降趋势明显,L-phe积累近乎停滞。
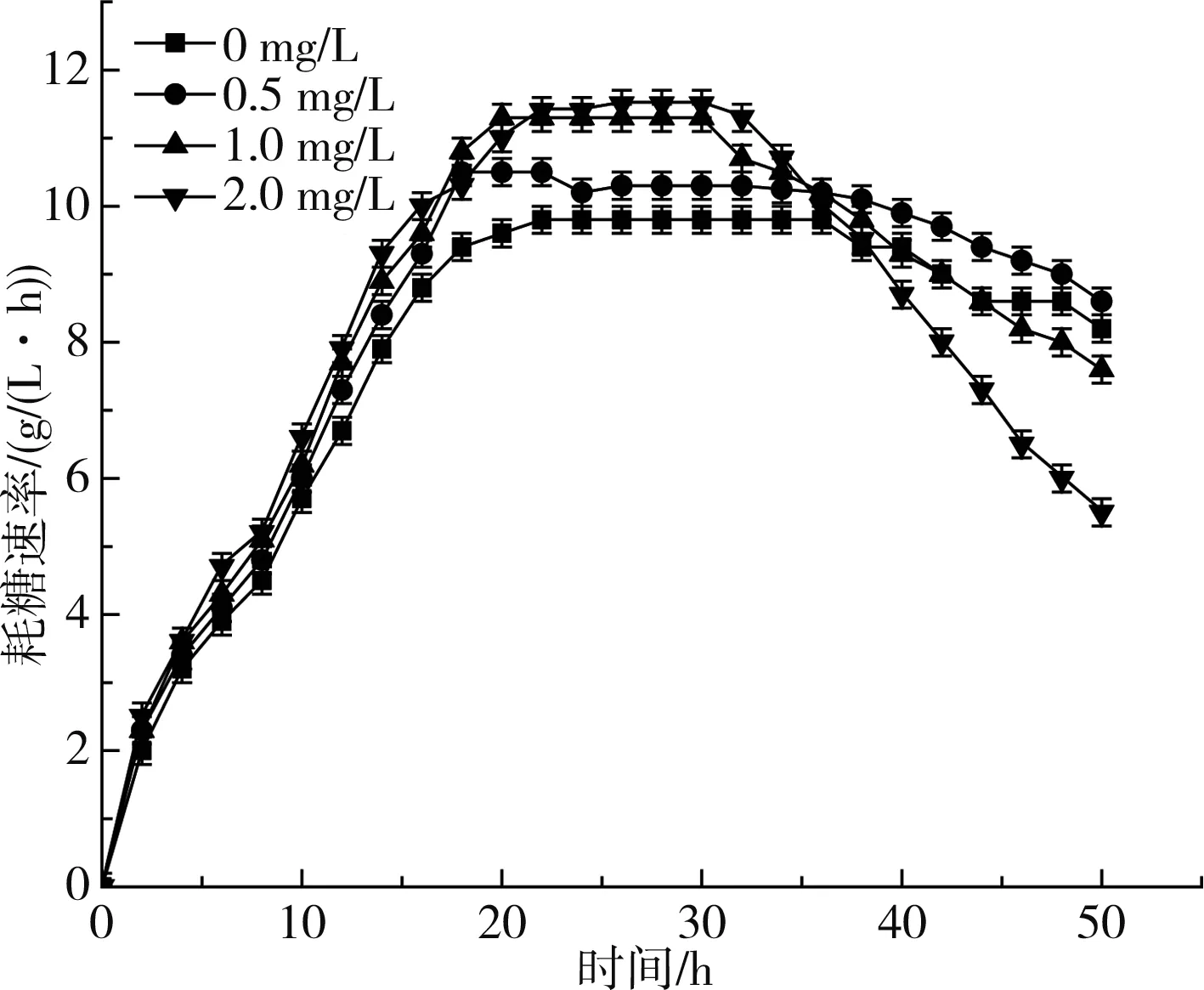
图2 不同浓度维生素H对应的耗糖速率Fig.2 Sugar consumption rates of different concentrations of vitamin H
由图2可知,各组发酵前期耗糖速度均提升迅速,添加维生素H浓度越高,对应最大耗糖速率越大,稳定期越短,后期下降速度越快,这与菌体量变化趋势一致。最终各组(添加0,0.5,1,2 mg/L维生素H)分别积累了80.7,84.8,78.3,71.0 g/L的L-phe,其中以0.5 mg/L表现最优。
2.1.2 维生素H浓度副产物及糖酸转化率的影响
为了更直观地分析维生素H浓度对产酸效率的影响,分别检测了副产物谷氨酸、乙酸的产量,并计算了糖酸转化率,见图3。

图3 不同浓度维生素H对副产物及糖酸转化率的影响Fig.3 Effects of different concentrations of vitamin H on the by-products and sugar-acid conversion rate
由图3可知,随着维生素H添加量的提高,乙酸与谷氨酸均呈明显上升趋势,1,2 mg/L维生素H添加组乙酸较对照组(1.23 g/L)分别提高了70%、135%,谷氨酸较对照组(2.20 g/L)分别提高了90%、140%,0.5 mg/L添加组乙酸与谷氨酸较对照组仅分别提升了8%、9%;糖酸转化率随着维生素H添加量的提高呈下降趋势,0.5,1,2 mg/L添加组较对照组(25.3%)分别降低了2.3%、15.1%、19.7%。
2.2 全营养流加工艺对L-phe发酵生产的影响
全营养流加工艺是指将传统分批补料发酵中一次性投入罐内的发酵培养基分成底物培养基与添加培养基两部分,底物培养基一次性投入罐内,为发酵初期提供营养物质,流加培养基在发酵过程中按需流加,为不同生长阶段的菌体提供最适的营养物质浓度。本研究中,流加培养基体积为400 mL,底物培养基按需求配制。如:配制3 L用量培养基,底物培养基∶添加培养基为2∶8,则用500 mL自来水溶解相应试剂,400 mL为流加培养基,100 mL为底物培养基。
2.2.1 底物培养基与流加培养基比例对L-phe发酵生产的影响

图4 底物培养基与流加培养基比例对L-phe发酵生产的影响Fig.4 Effects of the ratios of substrate medium to fed-batch medium on the production of L-phe by fermentation
本部分研究以2.1中额外添加0.5 mg/L维生素H的发酵组为对照,并在培养基中添加0.5 mg/L维生素H。由图4可知,A组菌体前期生长受到明显的抑制,最大菌体量远低于其他组,但其生长周期为34 h,高于B、C、D组,前期营养物质饥饿状态导致菌体在生长对数期无法最大程度地扩大菌体量,发酵后期产物与副产物的积累导致菌体生长停滞,此时流加培养基无法充分发挥作用,受菌体量影响产量亦低于其他组;D组流加策略并未起明显作用,菌体生长曲线和产物生成曲线与对照组基本一致,50%的营养物质已经足够支撑菌体前期的生长,随着流加培养基添加量的增加,整个过程依旧处于营养过饱和状态,并不能发挥出流加的优势;C、D两组恰好处于营养物质亚适量状态,即发酵各个时期营养物质恰好能够支撑菌体生长,既能保持菌体活力,又能最大程度利用营养物质,最终L-phe产量分别为86.0,87.5 g/L,较对照组分别提高了1.4%、3.1%,最大菌体量分别为144.1,149.1,较对照组分别提高了0.7%、6.5%,达到菌体量稳定期的时间均为32 h,较对照组延长了6 h。
2.2.2 底物培养基与流加培养基比例对副产物及糖酸转化率的影响
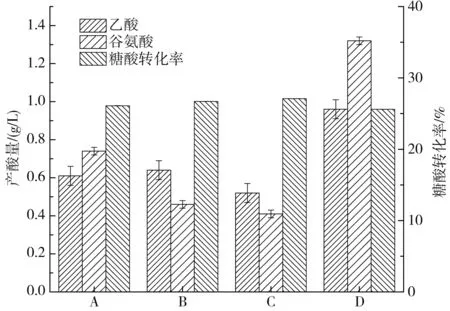
图5 不同底物培养基与流加培养基比例对副产物及糖酸转化率的影响Fig.5 Effects of different ratios of substrate medium to fed-batch medium on the by-products and sugar-acid conversion rate
由图5可知,D组乙酸、谷氨酸产量(0.96,1.32 g/L)分别比对照组降低了28%、45%,糖酸转化率(25.6%)比对照组提高了3.6%,因此即使同样是发酵前期营养过饱和,浓度越高的营养物质对发酵的影响越大;A组副产物均远低于对照组,糖酸转化率(26.1%也得到了很大提升,但其产量并不出众;B、C两组乙酸产量分别为0.64,0.52 g/L,较对照组分别降低了52%、61%,谷氨酸产量分别为0.46,0.41 g/L,较对照组分别降低了80.1%、82.9%,糖酸转化率分别为26.7%、27.1%,较对照组分别提高了7.4%、9.7%。
2.2.3 流加培养基流加时间与流加速度对L-phe发酵生产的影响
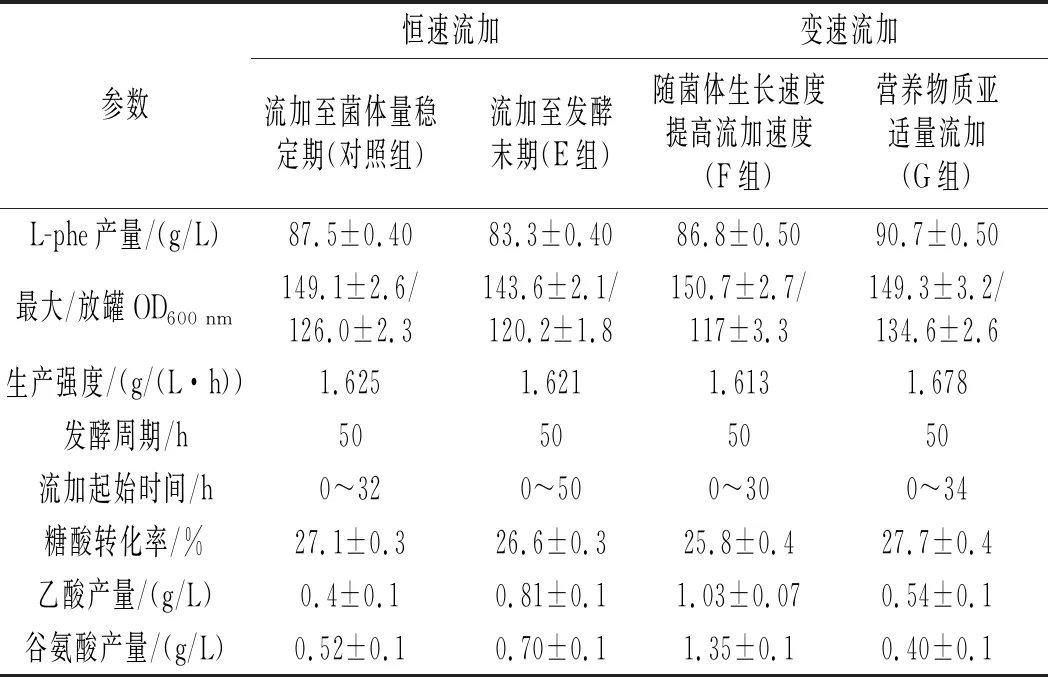
表1 流加培养基流加时间与流加速度对L-phe发酵生产的影响Table 1 Effects of feeding time and feeding speed of fed-batch medium on the production of L-phe by fermentation
以2.2.1中C组(对应表1中恒速流加至菌体量稳定期)为对照组,在前期优化基础上进一步优化全营养流加工艺。
表1中对照组恒速流加至菌体量稳定期指将400 mL流加培养基在32 h内恒速流加完毕,即12.5 mL/h,同理E组指8 mL/h。葡萄糖亚适量是指将罐内残糖浓度控制在0~0.3 g/L,在溶氧上表现为DO围绕某一值上下波动,溶氧曲线波动前行。F组随菌体生长速度对数提高流加速度是指在葡萄糖亚适量状态下,使得DO始终保持稳定的较低状态。G组营养物质亚适量流加是指在葡萄糖亚适量状态下,DO缓慢上升,提高补糖速率DO未见明显下降,流加营养物质DO迅速下降,即认为此时为营养亚适量状态,应提高流加培养基流加速度。
E组OD、产量较对照组分别下降了4.2%、3.6%,达到菌体量稳定期的时间较对照组延长了2 h,全程流加使得单位时间内菌体获得的营养物质变少,生长受限,后期流加液不仅不能为发酵提供营养,反而会占用发酵罐体积,迫使放出部分发酵液,浪费了具有产酸能力的菌体;F组在30 h时达到了最大菌体量150.7,但发酵结束时菌体量仅为117,虽然对L-phe产量的影响并不大,但副产物乙酸、谷氨酸较对照组分别提高了150%、159%,糖酸转化率降低了4.79%,F组使得发酵各个阶段的营养物质相对来说过于丰富,发酵中后期过高的菌体密度不利于溶氧水平和补糖速率的控制,乙酸的积累加快了菌体衰亡的速度,并不适合生长耦联型产物L-phe的积累。
G组最终L-phe产量达到了90.7 g/L,较对照组提高了3.6%,达到菌体量稳定期的时间为34 h,较对照组延长了2 h,糖酸转化率较对照组提高了2.2%,乙酸产量较对照组提高了35%,但乙酸浓度并未达到限制菌体正常生长代谢的浓度[24]。
2.2.4 维生素H添加方式对L-phe发酵生产的影响
本部分研究以2.2.3中G组(对应图6和图7中H组)为对照组,并在对照组的基础上探究维生素H添加方式对L-phe发酵生产的影响。图6和图7中G组的添加方式为将所有维生素H全部加入底物培养基;H组的维生素H添加方式为随底物培养基与流加培养基比例分配;I组的添加方式为将所有维生素H全部加入流加培养基。
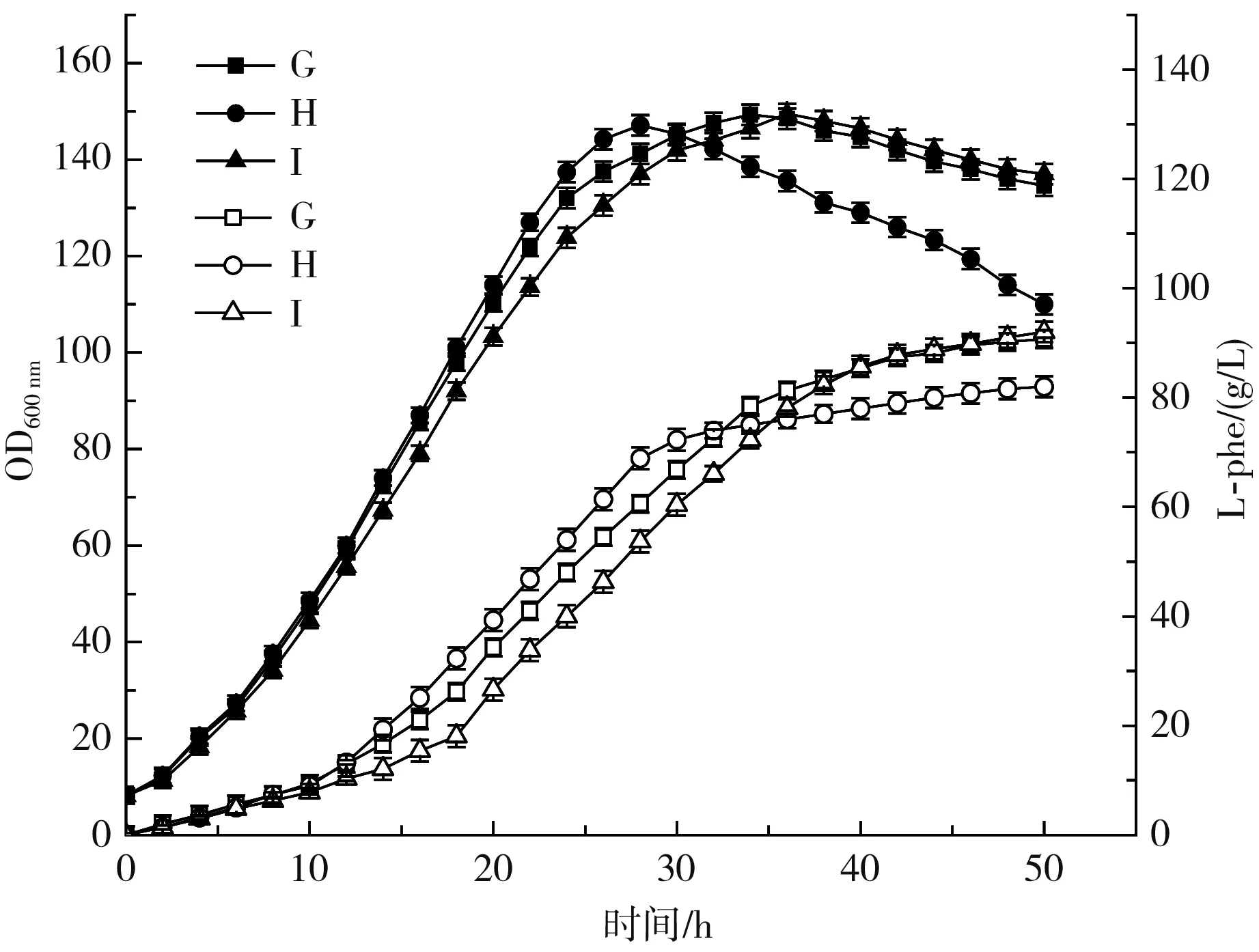
图6 维生素H添加方式对L-phe发酵生产的影响Fig.6 Effects of vitamin H addition methods on the production of L-phe by fermentation

图7 维生素H添加方式对副产物及糖酸转化率的影响Fig.7 Effects of vitamin H addition methods on the by-products and sugar-acid conversion rate
由图6和图7可知,G组菌体在28 h达到了最大菌体量(147.1),与不采取全营养流加策略时相同,前期产酸能力高于H组、I组,但总体产量下降明显,副产物谷氨酸产量达到了2.31 g/L,比对照组提高了477%,这说明代谢流更多地流向TCA循环,一方面降低了糖酸转化率,另一方面加大了提取难度,影响了产品质量;I组达到菌体量稳定期的时间为36 h,较对照组延长了2 h,L-phe产酸量达到了92 g/L,较对照组提高了1.4%,最大菌体量、发酵结束时菌体量、副产物、糖酸转化率与对照组均无明显差异。
3 结论
在大肠杆菌发酵法生产L-phe工艺中,提高转化率、生产强度,抑制副产物生成,是目前亟待解决的问题。本研究对发酵培养基中维生素H浓度进行了优化,确定了维生素H的最适浓度为0.5 g/L,在此基础上对发酵工艺进行优化,确定了底物培养基∶流加培养基为4∶6,流加培养基流加速度随罐内溶氧值变化变速流加至菌体量稳定期,维生素H全部加入流加培养基的全营养流加工艺,最终实现了L-phe产量与糖酸转化率的同步提高,且大幅降低了副产物谷氨酸、乙酸的积累。最终L-phe产量达到了92 g/L,较优化前提升了14.0%,糖酸转化率达到了27.7%,较优化前提升了9.5%,乙酸产量为0.54 g/L,较优化前降低了56.1%,谷氨酸产量为0.4 g/L,较优化前降低了81%。底物培养基和流加培养基的比例决定了发酵前期营养物质浓度,进一步决定了发酵前期菌体生长趋势;流加速度及流加方式决定了发酵过程中菌体的生长速度与生长周期,进一步决定了最终产量与糖酸转化率,是全营养流加工艺的核心。很显然,全营养流加工艺解决了添加维生素H带来的糖酸转化率降低的问题,同时大幅降低了副产物的积累,提高了产品的商业竞争力。
