不同泥炭替代基质对薄壳山核桃幼苗生长及叶绿素荧光特性的影响
2022-11-29季艳红潘平平窦全琴谢寅峰
季艳红,潘平平,窦全琴,谢寅峰*
(1.南京林业大学,南方现代林业协同创新中心,南京林业大学生物与环境学院,江苏 南京210037;2.江苏省林业科学研究院,江苏 南京 210014)
薄壳山核桃(Caryaillinoinensis)为胡桃科(Juglandaceae)山核桃属(Carya)植物,原产美国和墨西哥北部,是世界著名的干果、木本油料和优质用材树种[1-3]。我国引种薄壳山核桃已有100多年的历史,在华东等地引种栽培生长良好并已经开花结实[4]。江苏是国内最早引种薄壳山核桃的省区之一,由于薄壳山核桃适应性强、耐水性好,在长江湖泊滩地表现出较好适应性和生长潜力,是江苏省最适宜栽培的多用途树种。但薄壳山核桃为强深根性树种,主根发达,其长度可达苗高的2~3倍,侧根很少,在采用传统的大田育苗方式时,其主根的生长势旺盛,从而抑制了侧根的发育和生长,形成类似“胡萝卜”的根系,致使造林成活率较低,移植缓苗期长,从而制约了薄壳山核桃规模化种植与产业的发展。容器育苗具有节约种子、造林成活率高、无缓苗期,以及苗木规格和质量易于控制等优点[5-6],是薄壳山核桃良种苗木培育的较佳选择。在容器育苗的基质选择中,相关研究表明泥炭、蛭石混合物是容器育苗的理想基质。但是随着泥炭资源的日益枯竭,国外也在利用当地农林废弃物作为育苗基质材料,如荷兰开发了岩棉替代泥炭,波兰用泥炭与枯枝落叶混合配制成基质;德国用工业啤酒花肥料经过一系列处理作为容器苗培养的基质等[7-10]。
国内在农林废弃物作为基质方面的研究也已取得初步进展[11-16]。在育苗基质研究方面,多位学者研究表明,在基质中添加一定比例的菌渣、醋糟等农林废弃物,基质的理化性质及其培育的幼苗生长量均高于传统土壤基质[17-20]。贺殊敏等[21]研究表明,在基质中添加腐熟的木耳菌渣,有利于番茄(Solanumlycopersicum)幼苗的生长;胡小京等[22]研究发现,菌渣、松树皮、水苔体积比为1∶2∶1的基质配方有利于石斛(Dendrobiumnobile)苗的生长,有效增加幼苗的叶绿素、可溶性糖等含量。薄壳山核桃容器育苗在基质选择方面的研究目前集中于不同基质配比方面,常君等[23]的研究表明,以泥炭、珍珠岩、蛭石体积比4∶4∶2为基质可以获得根系发达、生长健壮的薄壳山核桃容器苗。以上研究表明,在基质中添加菌渣、醋糟等农林废弃物不仅对苗木的生长具有明显的促进作用,并使农林废弃物得到再利用,减少浪费,节省资源。但农林废弃物作为基质用于薄壳山核桃容器育苗的报道尚少。本研究通过在基质中添加菌渣、醋糟等废弃物以减少泥炭用量,分析各基质配方特性及其对薄壳山核桃幼苗生长和叶绿素荧光特性的影响,以筛选出育苗效果好、易获取、成本低、可循环利用的经济环保型基质,为培育薄壳山核桃优质容器苗提供科学依据,也为农林废弃物资源化再利用探索新途径。
1 材料与方法
1.1 供试材料
试验材料为薄壳山核桃品种‘绍兴’的子代1年生苗。种子于2018年1月中旬进行沙藏催芽,3月10日播种于沙床中,4月20日开始萌发,于5月11日移栽至不同基质配比的美植袋中,容器规格为20 cm × 32 cm(口径×高度)。
育苗基质原料为当地易得的、来源稳定的园土、泥炭、菌渣、醋糟4种材料。供试园土为扬州市江都区长江冲积土;泥炭为进口拉脱维亚(QTS,纤维长10~30 mm);菌渣为泰州市森鑫食用菌专业合作社生产经发酵1 a的杏鲍菇废料;醋糟为江苏恒顺醋业股份有限公司发酵制成(表1)。

表1 基质原料的化学性质
1.2 试验设计
试验在江苏省林业科学研究院苗圃进行。采用单因素随机区组试验,设置园土、泥炭、醋糟体积比为4∶3∶3、4∶2∶4、4∶1∶5(分别以A1、A2和A3标记,统称A基质)的3个配方和园土、泥炭、菌渣体积比为4∶3∶3、4∶2∶4、4∶1∶5(分别以B1、B2和B3标记,统称B基质)的3个配方,以园土为对照(CK),共7个处理,每处理小区30株,重复3次。
1.3 测定指标方法
1.3.1 薄壳山核桃容器苗基质理化性质的测定
基质的物理性质测定参考《土壤理化分析》[24],化学性质参考《土壤农业化学分析方法》[25]。土壤理化分析包括移栽定植前各处理的基质含水量、容重、持水量和通气孔隙度的测定,以及基质中有机质、pH、全氮、速效钾和有效磷含量的测定。有机质含量采用重铬酸钾容量法,pH采用电位测定法,全氮含量采用凯氏定氮法,速效钾含量采用火焰光度计法,有效磷含量采用钼锑抗比色法。
1.3.2 薄壳山核桃容器苗生长指标的测定
每小区选生长条件较为一致的20株苗挂牌标记,进行跟踪测定。2018年6月15日—9月15日,每15 d测量1次苗高和地径,共测定7次,以9月15日幼苗生长停滞时的幼苗苗高、地径为当年生长量。
1.3.3 薄壳山核桃容器苗叶绿素荧光参数的测定
于2018年6月12日、7月12日、8月12日和9月12日,采用CF Imager叶绿素荧光成像系统(英国Technologica公司生产)测定各处理的原初光能转化效率(Fv/Fm)、PSⅡ潜在活性(Fv/F0)、PSⅡ有效光化学效率(Fv′/Fm′)、实际光化学效率(ΦPSⅡ)、光化学猝灭系数(qP)和非光化学猝灭系数(NPQ)。选取有代表性的叶片,每个处理选9片,重复3次,用冰盒带回试验室,叶片先暗适应30 min,随后选择光辐射为800 mol/(m2·s),让叶片充分光适应5 min后获得光处理数据。
1.4 数据统计分析
数据采用Origin 8.5作图、DPS 16.05进行单因素方差分析、Duncan法多重比较。
2 结果与分析
2.1 不同配比基质理化性质分析
基质的物理性质与植物的根系生长密不可分,决定根系的生长环境。容重和孔隙度是简单衡量基质物理性质是否符合苗木生长的指标,在本试验中各处理基质的物理性质指标见表2。由表2可知,各处理的基质含水率、最大持水率、最小持水率、基质通气孔隙度含量处理均高于对照。基质含水量指标中,最高的是B1,最低的是B2,其他处理差异不大。对照组基质容重高于其他处理,容重最小的是A3。在最大持水量和最小持水量指标中,B3处理高于其他处理,最低的是对照。所有处理间基质通气孔隙度差异不明显,但是对照处理最低。
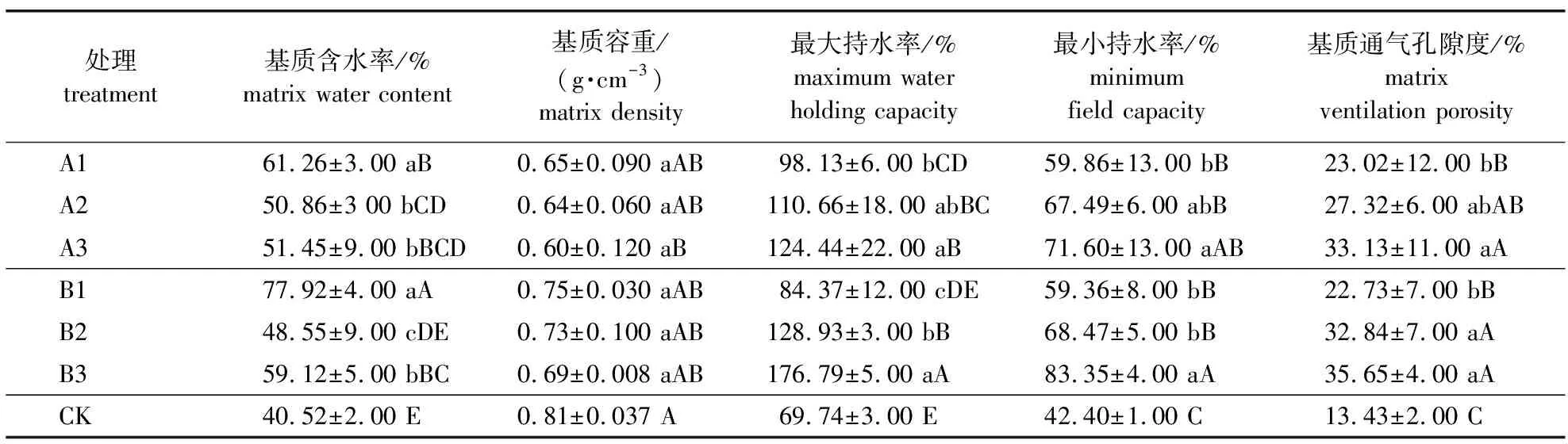
表2 不同配比基质物理性质分析
本试验所配制的基质pH为6.35~6.81。由不同配比基质化学性质(表3)可知,对照组的有机质、氮、有机磷、速效钾含量较低,B2处理有机质含量高于其他处理,A1、B2和B3处理全氮含量较高,分别是对照的1.56、1.67和1.56倍。A3和B3处理速效钾含量较低却显著高于对照。B2和B3处理有效磷含量较高,分别显著高出对照47.48%和48.62%。综合分析可知,不同基质配比和对照相比,基质含水量、最大持水量和最小持水量适中,通气孔隙度好,有机质、N、P、K含量高,可有效促进幼苗的生长。

表3 不同配比基质化学性质分析
2.2 不同配比基质对薄壳山核桃容器苗苗高和地径生长的影响
2.2.1 容器苗苗高和地径分析
由不同配比基质薄壳山核桃容器苗苗高和地径的动态变化(图1A)可知,苗高以及地径均呈现出先快速增长后缓慢增长的趋势,且不同配比基质处理均显著高于对照(P<0.05)。总体来看,B基质中的B2、B3处理效果要比其他处理好,但两者之间的差异不明显。6月15日—7月30日为薄壳山核桃容器苗的苗高快速增长期;8月30日—9月15日苗高增长缓慢且逐渐趋于稳定,在9月15日,B2和B3处理的苗高达到最大值,分别为34.67和33.47 cm,比对照分别高出64.00%和58.33%,可见薄壳山核桃容器苗在后期的高生长基本稳定。
与苗高有所不同,6月15日—7月15日所有处理的薄壳山核桃地径增长速率最大(图1B);7月15日—8月15日,B1、B2和B3处理的地径增长速率依然较大,A1、A2和A3处理的地径增长速率变慢。8月15日—30日,B1、B2和B3处理的地径仍然在增长,而A1、A2和A3处理的地径总体上趋于稳定。8月30日—9月15日,B3、B1、A1、A2和A3处理的地径增长速率变慢且趋于稳定,但B2处理的地径还在增长,说明B基质处理对地径增长速率有很大的促进作用,且以B2最佳。与苗高的动态变化相比,地径停止生长的时间较苗高晚,苗高与地径生长速率具有一定的非同步性,当苗高停止生长后,地径继续增大,最大值可达9.00 mm。由此结果显示,在基质中添加菌渣对于薄壳山核桃容器苗的生长有明显促进作用。
2.2.2 幼苗生长最终形态指标分析
苗高和地径是衡量植株生长的重要指标,由不同配比基质薄壳山核桃幼苗最终苗高和地径(表4)可知,不同配比基质对薄壳山核桃幼苗地上部分生长影响显著。B2处理的1年生幼苗苗高数值最大,为34.50 cm,B3处理的1年生幼苗地径数值最大,为8.60 mm。而苗高的最大值高于对照73.37%,最小值高于对照33.17%;所有处理中,地径最大值高于对照处理73.04%,最小值高于对照处理21.53%。不同处理的幼苗苗高从小到大的排序依次为CK < A3 < A1 < A2 < B1 < B3 < B2;地径从小到大的排序依次为CK < A1 < A2 < A3 < B1 < B2 < B3。以添加菌渣B2、B3基质更适合苗木的生长。

表4 不同配比基质薄壳山核桃幼苗最终苗高和地径
如不同配比基质的薄壳山核桃容器苗形态大小(图2)所示,含有菌渣B配方基质栽培的薄壳山核桃1年生幼苗较醋糟配方A处理生长好,其中以配方B3培育的苗木生长健壮,其次是A3,再次是B2。
2.3 不同配比基质对薄壳山核桃容器苗叶绿素荧光参数的影响
2.3.1 薄壳山核桃叶片Fv/Fm的动态变化
由薄壳山核桃容器苗叶片荧光参数Fv/Fm的动态变化(表5)可知,随着菌渣的不断增加,Fv/Fm数值不断上升。6月薄壳山核桃叶片Fv/Fm在0.778~0.820的范围内,对照的Fv/Fm低于所有处理。7月薄壳山核桃叶片Fv/Fm较6月相比出现下降趋势,A2和A3处理幼苗的Fv/Fm显著高出A1处理1.30%和2.60%,B1和B2处理幼苗的Fv/Fm显著低于B3处理2.47%和2.10%。8月薄壳山核桃叶片Fv/Fm较7月相比出现上升的趋势,A1、A2和A3处理的Fv/Fm上升幅度较大,分别上升了5.19%、3.85%和3.92%。9月薄壳山核桃叶片Fv/Fm较8月出现下降趋势,B1处理幼苗Fv/Fm低于对照,其他所有处理幼苗的Fv/Fm均高于对照,B3处理幼苗的Fv/Fm显著高出B1和B2处理3.04%和1.88%,且B3处理幼苗的Fv/Fm与其他所有处理之间差异显著(P<0.05)。综合比较可知,6—7月B基质处理的Fv/Fm高于A基质处理,8—9月A基质和B基质处理的Fv/Fm之间差别不显著。

表5 薄壳山核桃容器苗叶片原初光能转化效率(Fv/Fm)的动态变化
2.3.2 薄壳山核桃叶片Fv/F0的动态变化
由薄壳山核桃容器苗叶片荧光参数Fv/F0的动态变化(表6)可知,6月B基质处理薄壳山核桃幼苗叶片Fv/F0均显著高于A基质处理(P<0.05)。7月薄壳山核桃叶片Fv/F0为3.22~4.01,以B2处理和A2处理的Fv/F0较高;A2处理幼苗的Fv/F0显著高出A1和A3处理7.61%和13.14%。8月薄壳山核桃叶片Fv/F0均显著高于对照(P<0.05),B处理高于A处理,A基质中A2和A3处理幼苗的Fv/F0分别显著高出A1处理5.70%和6.18%。9月薄壳山核桃叶片Fv/F0在A基质和B基质所有处理中均差异显著,其中在A基质中A1处理幼苗的Fv/F0较高,而在B基质中B3处理幼苗的Fv/F0较高,在基质不同配比比例相同的情况下,B2和A2处理之间、B1和A1处理之间的Fv/F0差异显著,且B2高出A2处理10.64%,B1处理低于A1处理25.27%。综合比较可知,6—8月B基质处理幼苗的Fv/F0高于A基质处理幼苗,9月A基质处理幼苗的Fv/F0高于B基质处理幼苗。

表6 薄壳山核桃容器苗叶片PSⅡ潜在活性(Fv/F0)的动态变化
2.3.3 薄壳山核桃叶片Fv′/Fm′的动态变化
由薄壳山核桃容器苗叶片荧光参数Fv′/Fm′的动态变化(表7)可知,6月所有处理薄壳山核桃叶片Fv′/Fm′均显著高于对照,在A基质中A1处理幼苗的Fv′/Fm′较高,在B基质中B2处理幼苗的Fv′/Fm′较高,但A1处理显著低于B2处理。7月薄壳山核桃叶片Fv′/Fm′较6月出现下降趋势,其中A1、B2和B3处理的Fv′/Fm′下降的幅度较大,分别下降了11.78%、18.29%和22.89%。8月薄壳山核桃叶片Fv′/Fm′均显著高于对照,在基质配比相同的情况下,B基质所有处理的Fv′/Fm′高于A基质处理。除了对照,9月其他所有处理间薄壳山核桃叶片的Fv′/Fm′差异不显著,所有处理的Fv′/Fm′在0.511~0.581的范围内,其中B3和A2处理的Fv′/Fm′分别显著高出对照13.70%和12.33%。综合比较可知,除了7月,B基质所有处理幼苗的Fv′/Fm′高于A基质处理幼苗。

表7 薄壳山核桃容器苗叶片PSⅡ有效光化学效率(Fv′/Fm′)的动态变化
2.3.4 薄壳山核桃叶片ΦPSⅡ的动态变化
由薄壳山核桃容器苗叶片荧光参数ΦPSⅡ的动态变化(表8)可知,6月薄壳山核桃叶片ΦPSⅡ均高于对照,A1处理显著低于其他处理,其他处理之间差异不显著。7月薄壳山核桃叶片ΦPSⅡ为0.356~0.419,A3和B3处理幼苗的ΦPSⅡ较高,分别高出对照12.08%和17.70%。8月A基质中的A1处理薄壳山核桃叶片ΦPSⅡ显著低于A2和A3处理的4.59%和7.65%,在B基质中B3处理薄壳山核桃叶片ΦPSⅡ显著高出B1和B2处理的5.49%和4.44%。9月,A1和B3处理中薄壳山核桃叶片ΦPSⅡ较低,都为0.364,在其他处理中ΦPSⅡ较高。综合比较可知,7—8月B基质处理的ΦPSⅡ高于A基质。
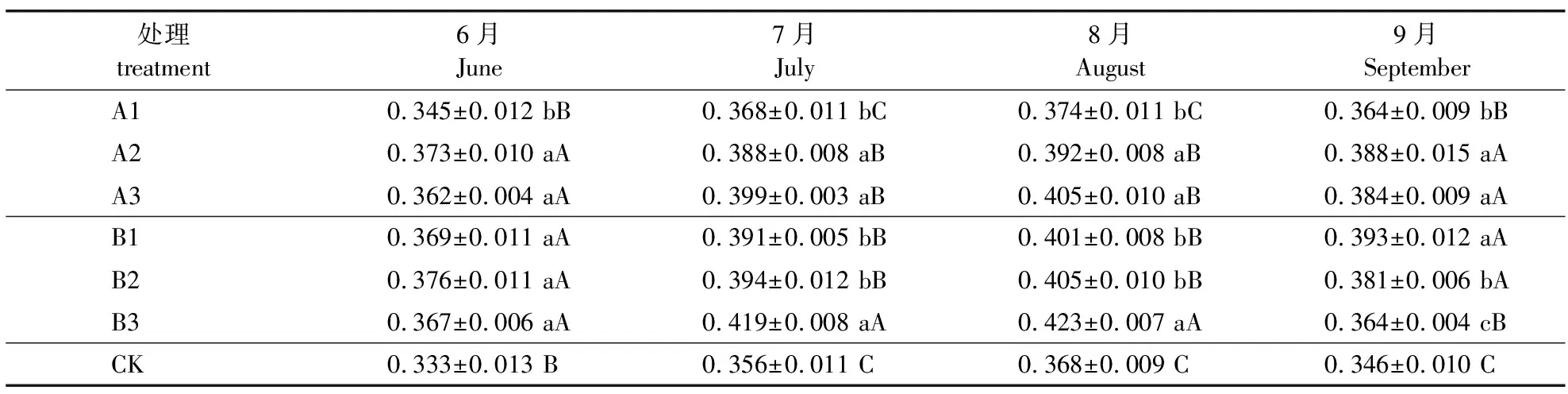
表8 薄壳山核桃容器苗叶片实际光化学效率(ΦPSⅡ)的动态变化
2.3.5 薄壳山核桃叶片qP的动态变化
由薄壳山核桃容器苗叶片荧光参数qP的动态变化(表9)可知,6—7月在同一时间和同种基质配比下,随着醋糟(A基质)或菌渣(B基质)用量的增加和泥炭用量的减少,薄壳山核桃叶片qP呈现升高的趋势。6月薄壳山核桃叶片qP均高于对照,在A基质中A3处理幼苗的qP高于其他处理,在B基质中也有类似的情况。7月薄壳山核桃叶片qP为0.683~0.867,A3、B2和B3处理的qP较高,分别高出对照20.94%、22.06%和26.94%。8月,A2和A3处理的薄壳山核桃叶片qP要显著高于B2和B3处理,A1处理的qP较低。9月薄壳山核桃叶片qP在A2处理中较高,在A1处理中较低,B基质中所有处理幼苗的qP和A3处理之间差别不大。综合比较可知,6—7月B基质处理幼苗叶片的qP高于A基质处理。

表9 薄壳山核桃容器苗叶片光化学猝灭系数(qP)的动态变化
2.3.6 薄壳山核桃叶片NPQ的动态变化
非光化学猝灭系数(NPQ)值越高说明光能大部分以热能散失,数值低则表示光能大部分被山核桃吸收利用。由薄壳山核桃容器苗叶片荧光参数NPQ的动态变化(表10)可知,6月所有处理薄壳山核桃叶片NPQ均显著低于对照,在A基质中A3处理幼苗NPQ显著低于A1和A2处理9.78%和3.66%,B基质中B3处理幼苗NPQ显著低于B1和B2处理10.02%和3.75%。7月薄壳山核桃叶片NPQ为1.889~2.971,B3处理幼苗的NPQ显著低于其他处理(P<0.05),A1、A2和B1处理之间差异不显著。8月,A基质所有处理薄壳山核桃叶片NPQ均高于B基质所有处理。9月B基质中薄壳山核桃叶片NPQ 呈现先降低后升高的趋势,其中B1和B3处理之间差别不大,B2处理低于其他两处理。综合比较可知,6—9月A基质处理的幼苗NPQ高于B基质处理。

表10 薄壳山核桃容器苗叶片非光化学猝灭系数(NPQ)的动态变化
2.4 苗木生长指标与基质理化性质的相关分析
由苗木生长指标与基质理化性质的相关性分析(表11)可知,苗高与最大持水率和有机质含量呈显著正相关;地径与最大持水率呈显著正相关。有效磷与速效钾具有较高的相关性,这与配方基质中pH有关,适宜的pH利于磷酸盐、交换性钾离子的转化,在pH为6.0~6.5时,基质中磷、钾的有效性最高。

表11 苗木生长指标与基质理化性质的相关性分析
2.5 苗木生长指标与叶绿素荧光参数的相关分析
由苗木生长指标与叶绿素荧光参数的相关性分析(表12)可知,苗高和地径与Fv′/Fm′、ΦPSⅡ和qP呈显著正相关,苗高与NPQ呈显著负相关,而地径与NPQ呈极显著负相关。

表12 苗木生长指标与叶绿素荧光参数的相关性分析
3 讨 论
3.1 不同配比基质对于基质理化性质的影响
育苗基质的含水率、容重、持水量、通气孔隙度、有机质含量及各种矿质元素含量等直接影响植物生长[26]。郭世荣[27]和李谦盛[28]研究表明容重在0.1~0.8 g/cm3,通气孔隙度为15%~30%是植物生长较理想的基质。本试验中,添加不同比例的菌渣、醋糟后,基质的理化性质均有改善,配方基质容重、孔隙度等指标均优于园土;基质的化学性质反映对苗木养分供应的能力,化学组分直接影响基质pH,刘帅成等[29]研究发现,基质pH在5.8~7.0时利于植物根系生长,植物易吸收大量元素,基质中的养分含量也受酸碱度的影响,pH降低时,有效磷会随之增高。本试验配比基质的pH低于园土,有机质及N、P、K含量均显著高于对照,其中添加菌渣量较多的B2、B3基质的有机质、全N含量较高。总体上,B2、B3这两种配比基质的理化性质更适宜薄壳山核桃幼苗生长,基质容重较低,良好的通气孔隙度利于幼苗根系的呼吸;适宜的pH有利于基质有效磷、速效钾的释放,促进植物的吸收与生长,有利于根系的吸收与生长。
3.2 不同配比基质对幼苗苗高、地径的影响
苗高、地径是反映苗木生长状况的两个形态指标,从各处理苗木生长情况看,苗高在6月30日后开始出现分化,园土基质培育的幼苗苗高生长量显著低于A、B配方基质,随后的生长基本处于停滞状态,7月30日后含有醋糟配方A基质幼苗高生长较为缓慢,而添加菌渣的B2、B3基质幼苗苗高依然呈快速生长趋势,一直维持生长至8月底;各处理地径生长情况与苗高类似,地径的高峰生长期较苗高滞后15 d左右,添加菌渣、醋糟配方基质中幼苗的地径生长均显著优于园土,其中以B2、B3基质幼苗地径生长量较高,这与其全氮含量高有关,与朱海军等[30]研究认为高N量缓释肥有利于幼苗体内N养分累积量和含量提高的结果相似,氮是构成蛋白质的主要成分,对植物茎叶的生长发育有重要作用,是与产量最密切的营养元素。因此,充足的氮元素是影响薄壳山核桃细胞的分裂和生长的重要调节成分。
本试验结果显示,幼苗苗高、地径与最大持水量呈显著正相关,水分是影响薄壳山核桃生长的重要因素。基质通气孔隙度与含水量相关性较高,较高通气孔隙度有较多毛管孔隙,增加基质持水保水能力和水气协调,利于植物根系伸展和生长,这与国外学者研究得出基质的物理性质较化学性质更为重要的结果一致[31-32];在基质化学性质方面,苗高与有机质含量呈显著正相关,地径与基质其余化学性质间无显著的相关性,表明基质中养分含量较低尚不能满足薄壳山核桃幼苗期生长的需要,潘平平等[33]研究表明,薄壳山核桃1年生容器苗生长所需大量元素为N 3.5%、P 2.8%、K 1%;季艳红等[34]认为3.5 g/株平均施肥方式有利于容器苗的生长。因此,苗木生长过程中所需要的养分需要定期定量施肥进行补充。
3.3 不同配比基质对幼苗叶绿素荧光的影响
光合作用为植物生长发育提供物质基础,是重要的生理生化过程,光合作用等生理指标也可有效地反映植物生长状况。叶绿素荧光参数在一定程度上可以反映植物叶片光合效率和潜在能力[35]。PSⅡ最大光化学效率(Fv/Fm),PSⅡ的潜在活性(Fv/F0)和PSⅡ有效光化学效率(Fv′/Fm′)在不同时间所有处理均高于对照,且B基质处理幼苗的这3个指标数值都高于A基质处理,说明B基质处理的薄壳山核桃原初光能转化效率和实际光能转化效率较高,光合效率和光合潜在能力较强。Fv/Fm常用来判断植物是否受到光抑制,在正常情况下该参数的范围在0.75~0.85,在逆境胁迫和光合器官受到伤害时该值会大幅度降低[36],在不同时期所有处理的Fv/Fm都在0.75以上说明其生长没有受到胁迫和光抑制。实际光化学效率(ΦPSⅡ)、光化学猝灭系数(qP)可反映光合活性的高低,试验中所有处理幼苗的ΦPSⅡ和qP值均高于对照,且8月的ΦPSⅡ和7月的qP均是B基质显著高于A基质,说明B基质的实际光化学效率和光合活性较高。非光化学猝灭系数(NPQ)反映的是当光合系统从环境中吸收的光能超出光合作用光能利用能力时,以热耗散的形式散失的光能[37],可以调节和保护光合作用受到过剩光能的损伤。试验中对照的NPQ值高于其他处理且B基质处理幼苗的NPQ低于A基质处理,说明对照处理的薄壳山核桃吸收的大部分光能以热的形式散失了,而B基质促进薄壳山核桃光能吸收利用效率。荧光参数分析结果显示,不同配比基质处理能够不同程度地提高山核桃容器苗叶片的Fv/Fm、Fv′/Fm′、ΦPSⅡ、qP,同时降低叶片的NPQ,表明不同配比基质处理能够提高山核桃叶片PSⅡ反应中心原初光能的捕捉效率,使有效光量子产量提高、电子传递速率增强,最终使得光合产物的增加,促进植株的生长。
相关分析结果可知,苗高和地径与Fv′/Fm′、ΦPSⅡ和qP呈显著正相关,理化性质较好的基质增强幼苗叶绿素荧光的能力,提高了苗木生长的光合效率;NPQ作为以热耗散的形式散失的光能,苗高与NPQ呈显著负相关,而地径与NPQ呈极显著负相关,表明适宜配比的基质可提高薄壳山核桃苗光能吸收利用效率,促进了幼苗的生长。
本试验旨在筛选出适宜培育薄壳山核桃幼苗的易获取、低成本基质配方,添加菌渣、醋糟混配基质较园土均可促进薄壳山核桃容器幼苗生长。通过比较不同配比基质下幼苗苗高、地径以及叶绿素荧光参数等指标发现,添加菌渣的配方基质对于薄壳山核桃的生长有明显的促进作用,特别是B3和B2基质均减少了泥炭用量,具有利于苗木生长的理化性质,且全N含量较高,薄壳山核桃容器幼苗生长表现良好,因此选择体积比为4∶1∶5的园土、泥炭和菌渣低成本配方基质用于薄壳山核桃容器苗的培育,取得良好的育苗效果,可在生产中推广应用,实现薄壳山核桃容器苗培育的资源节约型和环境友好型生产。
