中国食用向日葵育种国产化历程及研究进展
2022-11-26冯九焕
冯九焕
(三瑞农业科技股份有限公司 向日葵科学院,内蒙古巴彦淖尔 015100)
向日葵 (HelianthusannuusL.,2n=2x=34,基因组大小~3.5 Gb)属于菊科向日葵属,是为数不多的起源于北美州的作物之一,距今约有4000年历史,进化上属于比较新的物种[1-2]。目前已广泛种植于世界各地,是继大豆、油菜、花生之后世界第四大油料作物。2019年,全世界种植面积约2 736.88万hm2,总产量5 607.27万t。其中俄罗斯种植面积最大,约841.47万hm2;其次是乌克兰、阿根廷、罗马尼亚、坦桑尼亚,中国位居第6[3]。
向日葵属(Helianthus)共有51个种(其中14个一年生种,37个多年生种),包括栽培种和野生种[1]。其中栽培种,除观赏型向日葵外,又分为油用型(oilseed type)和食用型(non-oilseed type)。这种类型的划分主要是基于粒型、含油率及用途的不同按照传统标准人为界定的,不同于生物学上的亚种(subspecies)。油葵通常籽粒短小、黑壳、含油率高(50%~55%),蛋白质含量低(16%~19%),主要用于榨油;而食葵籽粒大、黑底白条纹、含油率通常较低(35%以下)、蛋白质含量高(25%~28%或更高),主要用于嗑食和甜点。
向日葵起源于北美洲温带地区(美国中东部),由当地印第安人从野生种选择单头(花盘)、大籽粒个体用于自食逐渐驯化而来的。直到18世纪前后向日葵被引入欧洲,并在前苏联首次作为一种“作物”规模化种植,成为深受欢迎的“国民零食 (national dainty)”[4]。这一时期,人们一方面选择大粒型用于嗑食;另一方面十分注重选育含油量高的品种。随着高油品种的育成,向日葵逐渐升级为前苏联重要的大田油料作物,相应地嗑食品种的选育渐渐被忽视。相反,美国作为向日葵的起源地,1930s前后,向日葵仅仅是一个小众作物,主要种植前苏联引进的一些农家种。直到1950s末,它才被看作是一种有经济价值的作物。但由于大豆等其他油料作物的竞争,向日葵仅用于烘烤食用或鸟类饲料[5]。然而,人们很快意识到,改良的向日葵品种比大豆产油潜力大[6]。自此,选育高含油量一直是世界各国向日葵育种的主要目标,并取得了巨大成功。含油量从1900s早期的300 g/kg,到1930s的400 g/kg,至1960s的500 g/kg。比如,Peredovik(派列多维克,又称‘先进工作者’)是前苏联种植面积最大的一个代表性品种,1966年达到1.2 Mhm2。该品种于1960年被引进美国,成为当时种植面积最大的油葵常规种[1]。1972年油葵杂交种被引入美国,1974年食葵杂交种被引进,但食葵总体上占比较小,种植面积一直保持在10%~20%。可见,从向日葵早期发展看,人们首先是自选自留大籽粒的嗑食品种(食用型)为主,其后先驱育种家们才逐渐注重高油品种选育(油用型),并因其重要的经济价值使油葵迅速发展。实际上,油葵和食葵在分类学上没有截然的界限,前苏联早期种植的油食兼用型品种(如Lakomka、Borodinskiy等)也印证了这一点[7]。
欧美国家向日葵育种研究无论在种质资源还是人员配置上,主要集中于油葵,只有极少数针对“大籽型(large seed)”、“嗑食型(cracking seed)”的选育项目[8]。食葵育种研究工作开展较早且较为系统的,其一是前苏联油料作物研究所(All-Russia Research Institute of Oil Crops,VNIINK),于20世纪末开始食葵常规种选育,1999年启动食葵杂交育种项目[9]。其二是塞尔维亚大田及蔬菜作物研究所(Institute of Field and Vegetable Crops,Novi Sad,IFVCNS),于1990s中期设立食葵育种项目,如今推出了系列NS食葵品种[7]。近几年来欧洲一些国家逐渐加强了食葵育种,食葵面积有增加的趋势。然而,由于多数育种家主要目标是高油含量,其他类型育种材料被遗弃,向食葵育种转型并非易事。
食用葵花籽经加工后直接嗑食这一休闲方式,已经被许多国家所认同,包括中国、俄罗斯、乌克兰、土耳其、西班牙等;同时葵花籽仁因其高营养价值广泛用于糕点等饮食业。当今,休闲与保健逐渐成为人们的消费趋势,食葵产业作为应运而生的朝阳产业具有广阔发展前景。中国食葵育种起步较晚,面对国内市场的刚性需求和国际市场变化,加之近年来企业育种力量异军突起,中国食葵育种取得巨大进展,培育出一系列优质高产食葵新品种,食葵面积占据绝对优势,使得油葵面积逐渐缩小(几乎没有准确的数据)。据不完全统计,近5年来中国食葵种植面积基本保持平稳,年均种植面积约在56万hm2上下波动,占全世界食葵总面积的一半左右;其他有一定种植面积的国家还有俄罗斯、乌克兰、土耳其、美国等;除此之外,阿根廷、塞尔维亚等也有小面积或零星种植[10](表1)。
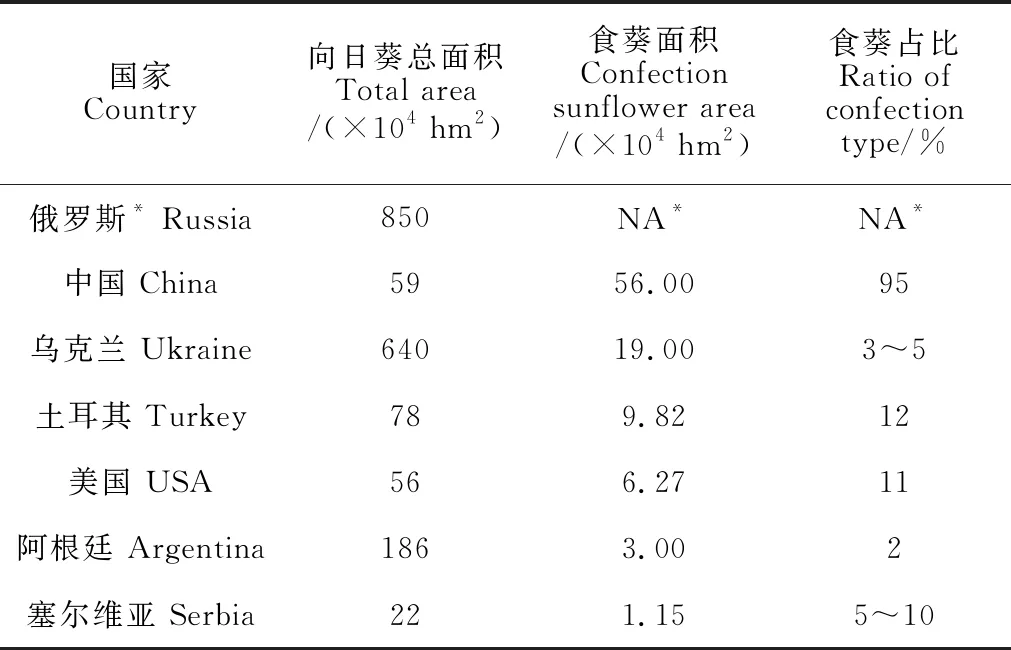
表1 世界向日葵主产国近年来食葵种植面积[10]
中国向日葵生产主要集中在北方冷凉地区,依托主产区逐渐形成了育种、生产、仓储加工、出口贸易以及副产品开发等独具区域优势的完整产业链[11]。近年来随着分子生物学技术的迅速发展,传统育种向分子育种的转型,为中国食葵育种带来了前所未有的发展机遇。然而,中国食葵育种起步晚,种质资源积累欠缺,育种技术单一,尤其是抗性育种重视不够。鉴于此,本研究对中国食葵育种发展历程、抗病育种、生物技术应用等领域的研究进展、存在的问题及未来方向做一梳理和总结,以期为推动中国乃至世界食葵产业的高质量发展提供借鉴。
1 中国食葵育种发展历程
半个多世纪以来,中国食葵育种经历了自留农家种、选育常规种、引进国外杂交种以及发展国产杂交种等循序渐进的发展历程(表2),使食葵成为近年来新崛起的一个重要特色经济作物。

表2 中国食用向日葵育种发展历程
1.1 从零星种植到主栽农家种(landraces)1950-1990年
中国具有栽培和食用向日葵的习惯和历史。向日葵自明代中后期传入中国,至1950年前后,仅零星种植于庭院用于观赏和嗑食[12-13]。自1955年起,中国正式开始向日葵引种鉴定、系统选育和品种杂交育种工作。吉林省白城市农业科学院以引进的‘匈牙利1号(依列基)’为基础材料,经系统选育于1962年育成了油食兼用型‘白葵3号’,在吉林、内蒙古、山东等省区推广[14]。同时,山西省农业科学院以油用型‘苏联2号(VNIIMK 6540)’和食用型‘晋北大马牙’为亲本,于1961年育成了极早熟品种‘太原80天’,在山西等地推广[15]。1965年底中国向日葵育种和栽培研究全部停止,并于1973年再次恢复。与此同时,美国却开始加大了对向日葵这一新型油料作物的育种研究力度,并于1964年在美国召开了第一届“国际向日葵大会(International Sunflower Conference,ISC)”,使得向日葵这一作物迅速得到全世界关注。可以看出,中国向日葵育种研究起步并不晚,但停滞了8年,使得整体落后于欧美国家。在这一时期,尤其是20世纪70-90年代,中国广泛种植的主要是油用向日葵,比如前苏联常规品种Peredovik,1985年种植面积曾达到12.9万hm2[11]。
1969年法国育种家Leclercq首次在野栽杂交后代中筛选出向日葵细胞质雄性不育系(PET1),随后,Kimman(1970)和Leclercq(1971)从多个一年生野生向日葵杂交后代中发现了育性恢复基因,从而使向日葵作物的杂种优势利用成为可能,1974年首批杂交种(FRANSO,LRELAX)在法国登记[16]。利用从加拿大引进的不育系及保持系,吉林省白城市农业科学院于1977年成功选育出中国第一个油葵杂交种‘白葵杂1号’,实现了三系配套[17]。此后,全国相继选育出‘内葵杂1号’、‘辽葵杂1号’、‘沈葵杂1号’等一系列油葵杂交种并推广种植[14,17-18]。自此,常规种逐渐被杂交种所取代,杂交种成为唯一商业化种植的向日葵类型。
可见,建国40年间,限于当时社会发展对食用油的迫切需求,主要科研力量均集中于油用向日葵研究。相比之下,作为休闲嗑食的食用向日葵研究几乎是空白,多数科研院所没有设立专门的食葵项目,即使选育了少数品种,也没有成为主导品种。各地农户仍种植自选自留的农家种(地方品种),包括全国普遍栽培的‘三道眉’(图1)、黑龙江等地的‘黑老鸹嘴’、内蒙古的‘星火花葵’、‘黑白边’,吉林等地的‘白油嗑’、‘长岭向日葵’、‘牙签’,甘南‘黑大嗑’、‘大马牙’、‘黒大片’、‘大青花’、‘黑背’、‘寸嗑’以及硕葵等[11,18-19]。可见,这一时期,食葵仍处于自选自留农家种阶段,油葵已进入了杂交种时代。全国向日葵种植产区从东部迅速向西北发展,1985年达到第一次高峰,种植面积约147万hm2,总产量为173万t,全国平均单产1 175.3 kg/hm2[20]。黑龙江省种植面积和产量居全国之首,油用型面积最高达60%左右[13]。

图1 代表性农家品种‘三道眉’
1.2 规模化种植常规品种(open-pollinated variety,OPV)1991-2000年
农家品种对当地土壤和气候条件具有较强的适应性和抗逆性,但不足之处是品种混杂、有分枝、一致性差、商品性差、产量低[21]。随着市场需求的与日剧增,以科研院所为主体的食葵品种选育研究取得长足进展。黑龙江省甘南县向日葵研究所以农家品种为基础群体,采用系统选育法育成了全省首个食用型向日葵常规品种‘甘葵1号’,结束了一直沿用农家种的局面。该品种1989年审定,5年内推广面积达13.3万hm2,深受种植户及炒货商欢迎[22]。此后,食葵新品种‘甘葵2号’(1998年审定)、‘甘葵3号’(2004年审定),以及吉林省白城市农业科学院以‘匈牙利4号’为原始群体,采用半分法(贮备法)育成的‘白葵6号’(2001年审定),黑龙江省农科院选育的‘龙食葵1号(2002年审定)、2号(2003年审定)和3号(2007年审定)’等常规品种相继推广上市,成为当时的主栽品种。这一时期,向日葵的生产主要集中在黑龙江、辽宁、吉林,以及内蒙古东部赤峰、新疆北部伊犁和塔城等地。全国年种植面积大约在81万hm2左右,总产量约138万t,平均单产约1 730 kg/hm2[20],油葵种植面积逐渐回落至30%左右[13]。
1.3 国外食葵杂交种垄断市场(foreign hybrids)2001-2010年
食葵营养价值高,主要用于嗑食、甜点等行业。由于食葵的休闲食品定位,较之油葵利润高,食葵市场迅速崛起,种植面积迅速扩大。这一时期,作为主要产区的黑龙江省种植面积通常在25万hm2以上,其中食葵面积占比达95%[23]。新疆由于食葵种植效益高,种植区域不断扩展,北疆沿天山一带开始大面积种植食葵,2008年面积上升至1.72万hm2[24]。但国内食葵仍以农家种或常规品种为主,常规种较之自留的农家种产量明显提高,粒大、色泽好,但主要缺点是抗病性差、成熟期晚及一致性差,完全不能满足国内食葵市场快速发展的需要,而这一时期国外食葵已普遍使用杂交品种。
2000年《中华人民共和国种子法》颁布,标志着中国现代种业的诞生。而在中国种业刚刚进入市场化的初期,国际种业巨头已形成全球扩展态势。随着市场全面开放,加之消费市场的刚性需求,种子公司开始从美国、以色列等大量引进食葵杂交种。这些进口包衣的杂交种整齐一致、抗病性好、产量高,迅速占领中国高端市场。杂交种价格比普通常规种高出10倍左右,每千克约100~200元,个别品种如LD5009每千克曾高达400元[25-27]。
这一时期,内蒙古巴彦淖尔市每年大约100多个国外油葵和食葵杂交种被引进和种植,巴彦淖尔地区几乎成为世界向日葵品种试验区。自2000年开始的5年间,内蒙古杂交种面积迅速升至60%~70%[25]。全国其他省区情况类似,2006-2007两年间,参加全国生产试验的9个食葵品种中,3个国内自育品种,6个为美国杂交种[26]。其中,有代表性的包括美国孟山都公司选育的DK119,于2000年审定后成为内蒙古种植的第一个食葵杂交种[28]。另一个代表性品种是内蒙古乌拉特种业2005年引进的美国杂交种LD5009,籽粒商品性好,粒长,无边际效应,抗病性强,丰产性好,仅2007年推广面积达1.33万hm2,成为当时巴彦淖尔市的主导品种[29]。此外,先后引进的国外杂交种包括SH316、SH909、3638C、DC6009、DK88、X3939、RH1122、RH3148、RH3108、HK306、765C和H658等在中国各地广泛种植。杂交种亩均产量由100 kg增加到250 kg左右,亩收入由600元增加到1500元以上,食葵种植面积剧增。这一时期杂交种使用率已达到95%,美国已成为中国最大的食葵种子供应者[26]。2001-2010的10年间,中国向日葵种植面积基本稳定在100万hm2左右,总产量接近175万t,平均单产1 813 kg/hm2以上[20],食葵种植面积升至70%以上,向日葵产区从东北迅速向西北发展,内蒙古成为最大的向日葵种植区[18]。
1.4 国产化食葵杂交种井喷式增长(domestic proprietary hybrids)2011-2020年
随着消费市场对食葵品质要求的提升,国外食葵杂交种缺陷日益凸显出来,籽粒偏短不适宜磕食,香味下降,商品性较差[30]。而且,进口种子价格昂贵,使投入成本增加[21]。为促进国内油料作物发展,2008年国家启动建设“向日葵产业技术体系”,由来自10个省区的科研院所组成的研究团队,成员达110余人,是中国向日葵研究领域有史以来科研人员最多的时期。除科研院所外,一些大中型种业公司,包括三瑞农业科技股份有限公司、甘肃同辉种业有限责任公司、河北双星种业股份有限公司、昌吉州西亚种子有限责任公司、酒泉市同庆种业有限责任公司、哈尔滨丰葵农业科技发展有限公司等也加大了向日葵育种研发力度[11],并针对国内食葵市场的需求特点进行品种选育。
对于食葵这一新的领域,除有限的农家品种‘三道眉’、‘星火花葵’以外,育种资源匮乏成为当时最大的瓶颈。利用国外食葵杂交种(F1)后代成为育种初期行之有效的手段和捷径。比如,从国外品种DK118、LD5009等自交后代中筛选优良个体,与自育的自交系杂交或连续回交,选育了一系列新的恢复系、保持系及相应的不育系[31-36]。据统计,2017-2019年登记的品种中,68%具有外来血缘,也从一个侧面印证了这一育种策略[37]。
由于市场对食葵种子的刚性需求,极大推动了科研院所和种业公司的育种研发,并相继选育出一系列高产、优质、长粒型的自主知识产权的食葵杂交种,使国产杂交种呈爆发式增长。据统计,2017-2019年间,全国共登记食葵新品种1 283份,其中由企业登记的占比达90%[37]。这一方面体现了企业在品种创新中的主导作用,另一方面反映出品种登记制度尚需不断完善,一品多名或套牌现象严重。2021年,全国所有已登记向日葵品种的DNA指纹数据库完成,不仅对拟登记的新品种进行真实性DNA指纹鉴定,而且对已登记入库的全部品种重新检测甄别,对仿冒品种撤销登记资格。据不完全统计,2021年拟劝退的登记品种中,与SH363相似品种约197个,与SH361相似品种约162个[11,38]。DNA指纹数据库的建立为向日葵种业的有序竞争和规范管理提供了技术支撑。
虽然登记品种数量大幅增加,但绝大部分品种没有推广或没有成为主栽品种。市场需求从追求高产逐渐过渡到以质论价。因此生产上普遍种植的仍然以优质高产品种以及新的抗列当系列为主,包括SH363、SH361、‘三瑞9号’、‘三瑞11号’、JK601、‘双星6号’、‘同辉33’、‘先瑞10号’、‘同庆5号’、‘利源1号’、HZ2399和‘西亚一号’等系列品种[37,39]。值得一提的是,中晚熟食葵杂交种SH363(图2),自2013年审定推广以来,以其良好的粒型及独有的香脆口感至今仍是炒货市场最优质的高端商品,成为食葵育种的标杆[40]。此外,2019年登记推出的中熟品种‘三瑞9号’(图3),一个全新的籽粒类型,以其产量突出籽仁香脆为特征,已经成为自SH363之后另一个深受市场青睐的食葵新品种。这一时期,食葵种植面积稳步上升,到2016年增加到128万hm2,创历史最高,总产量提高至320万t,单产最高达2 700 kg/hm2左右[20],其中食葵面积约95%以上,占据绝对优势。

图2 主栽食葵品种SH363籽粒

A.近成熟植株;B. 商品籽粒
近30年来,中国向日葵主要生产地由东向西发生了较大迁移,且生产更为集中,中国向日葵生产中心(以面积而论)由原来的赤峰市向西南方向迁移至巴彦淖尔市的乌拉特中旗。内蒙古产量和面积居全国之首,新疆面积持续增加,菌核病发生以及其他替代作物比如大豆种植政策的调整,一定程度上导致黑龙江、吉林、辽宁、山西和河北面积明显下降[41]。全国向日葵种植面积总体稳定在100万hm2左右,年均总产量基本保持在240~260万t[3]。经过近10年的发展,中国食葵品种完全实现了品种杂交化及自主选育,以美国食葵为代表的国外杂交种基本从中国市场消失[42]。随着生物技术的广泛应用及一系列市场监管措施的出台,兼顾产量、品质及多抗的国产食葵杂交种的选育和推广必将进入良性竞争与有序发展阶段。
2 中国食葵育种研究进展
向日葵因其耐干旱、耐盐碱、耐贫瘠等特性,成为中国西北干旱半干旱地区与粮食作物生态位互补的重要经济作物。为此,选育高产、优质、多抗、广适、宜机收向日葵品种,以满足市场多元化需求成为当今种业发展的重点。尤其是,在注重商品外观品质(粒大、色泽亮、口感香脆)的同时,提升商品营养成份(脂肪酸、蛋白质等)或开发专用型品种将成为未来抢占高端商品的突破口。然而,食葵是针对粒型选育出来的一个特殊类型,育种群体遗传基础狭窄,品种同质性高,与油葵相比抗病性普遍较差,使之存在更大的病虫草害突发性隐患。目前中国向日葵生产中,除少数细菌性和病毒性病害外,真菌性病害尤为严重,其中已报道或鉴定的危害严重的真菌性病害有菌核病、霜霉病、锈病、黄萎病、褐斑病、黑斑病、黑茎病和拟茎点霉病等;此外,寄生性杂草向日葵列当也已成为向日葵生产的首要制约因素[36,43-50],而抗性育种是解决这些问题的最经济途径和最佳手段。
目前,中国食葵抗病性研究绝大多数集中在病原菌生物学特性、病害发生规律、化学防控以及品种资源抗性鉴定等方面,有关抗性基因鉴定、遗传机理解析等研究几乎空白。尤其是对病害发生缺乏预警或有效方案,新选育的品种在推广过程中可能因病害爆发自然被市场所淘汰。究其原因,一方面可能是缺乏抗性资源;另一方面是缺乏可靠准确的检测手段,目前仍主要依赖田间自然发病进行判定,相关的分子标记开发不足。此外,新品种登记时对抗病性没有统一标准要求,缺乏约束性或导向作用。国际上,黄萎病、霜霉病、锈病等抗性基因早已在油葵中成功应用;同时,对危害最为严重的菌核病、茎溃疡病、黑茎病等抗性基因的研究也取得一定进展。面向未来,我们必须正视食葵抗病育种的现状和不足,充分利用和借鉴油葵优异资源或研究成果,普及分子标记辅助选择技术,提高育种效率,并重点加强以下几个方面的研究,加快推进中国食葵生物育种产业化应用。
2.1 启动持续性全国向日葵病害普查
随着向日葵长期栽培和连作种植,缺乏政府引导的强制性轮作制度,许多病原菌与向日葵发生协同进化,导致病虫害日趋严重。目前,全球已报道的向日葵病害至少有30余种[51]。一旦缺乏预警,或育种上缺乏前瞻性研究或抗性品种储备,将会对产业造成毁灭性的影响。国外十分重视向日葵病害田间调查,对于实际生产及规划未来优先研究领域均具有重要指导意义。早在1949年,Sackston[52]就对加拿大向日葵田间病害进行了调查和系统研究。而且,1989-2017年间,加拿大每年对向日葵最大产区曼尼托巴(Manitoba)进行定期病害调查[53]。1970s后期,欧洲多个国家也联合开展病害调查。美国于1984年首次对3个主要产区(北达科他州、南达科他州、明尼苏达州)的10种病害进行全面调查。自2002年起,美国向日葵协会(National Sunflower Association)协调和组织大学、种业企业、农药公司等,对至少10种主要病害进行全国普查并公开发布调查结果(2014年起每2年1次),其中包括霜霉病、茎溃疡病、黑根霉盘腐、锈病、菌核病等[54]。基于此项调查结果,美国农业部于2003年提出了国家菌核病研究启动计划(National Sclerotinia Initative),近几年又将茎溃疡病列为优先研究领域。可见及时跟踪病害的动态变化对向日葵产业可持续发展起着决定性的作用。病虫害发生与地域、气候条件密切相关,不同地区或不同年份的主要病害类型不尽相同,或为全域性的或局部性病害,或由以前的次要病害上升为主要病害[55]。在中国,加强全国性的、持续性的向日葵病害系统调查,并及时公开发布调查结果,不仅有利于向日葵种业企业规划育种方向,而且对种植者选择品种以及农药的研发均具有重要的指导意义,目前这方面的工作明显不足。未来,急需借助专业协会、科研院所及行业力量,开展全国性向日葵病害的持续性调查工作。
2.2 抗病育种
查阅近年来有关新品种选育及品种登记的部分论文,可以看出新品种“抗性评价指标”主要涉及菌核病、黑斑病、褐斑病和黄萎病等,从一个侧面反映出上述4类病害是影响中国向日葵生产的主要病害或较常见病害;除此之外,个别情况下也包括锈病、霜霉病及黑茎病[31-32,34-36,56-63]。国内外大量的遗传研究已揭示,黄萎病、锈病、霜霉病、列当等是由单基因控制,而菌核病、茎溃疡、黑茎病、黑斑病等则由多基因控制,并相应地筛选出一些抗病种质。为加快推动食葵抗性育种,现对国内外抗病研究进展进行简要概述。
2.2.1 菌核病(sclerotinia) 菌核病是由核盘菌[Sclerotiniasclerotium(Lib.) de Bary]引起的全球性的,迄今为止最顽固的一种向日葵真菌病害。菌核病在田间可表现出根腐、茎腐和盘腐3种不同症状,均是由同一病原菌引起。夏季低温多雨是导致该病发生的主要诱因。东三省黑龙江、吉林和辽宁曾是中国最早大面积种植向日葵的主产区,由于大面积重茬种植以及降雨量大等气候因素影响,导致菌核病发生逐年加重[64]。1958年黑龙江嫩江菌核病发病率50%,1959年发病率高达80%,个别地块绝收[65]。2009-2014年,黑龙江年平均发病率在30%~50%,有些地块达到80%以上[66]。菌核病已经成为当地向日葵产业发展的主要制约因素,黑龙江种植面积从2002年前后的22万hm2左右[13],回落到目前不足2万hm2。吉林省由2013年的11.1万hm2下降至7.8万hm2,导致原有的种植老区如洮南市几乎没有规模化种植[67]。目前,内蒙古东部的赤峰地区面临着同样问题,受菌核病影响部分地区已停种[68]。可以预料,若短期内仍无法有效控制菌核病,向日葵在上述产区的种植面积3~5年内将会大幅缩减[45]。抗菌核病品种选育和推广是恢复东北主产区的根本保障。
大量的研究已证实,菌核病是由多基因控制的数量性状,尚未发现高抗或免疫资源[64],抗性育种难度较大。中国自1980s开始菌核病育种工作,多数研究主要集中在病害发生规律、防治措施、表型鉴定及品种抗病性比较等方面[66,69-71]。采用人工接种或田间调查,从野栽后代、地方资源、自交系或杂交种等材料中筛选出一些中抗品种,但未发现高抗或免疫种质[72-74]。国外十分重视菌核病抗性资源挖掘及鉴定,比如美国农业部于2003年启动的菌核病专项,覆盖向日葵、大豆、油菜、菜豆及其他豆类作物,系统地开展菌核病抗性机理研究,取得了一定进展。从不同向日葵野生种,包括H.petiolaris(PI435843)、H.argophyllus(PI494573)、H.praecoxsubsp.runyonii(PI468853)与栽培品种的杂交后代中,多份耐菌核病遗传材料被鉴定和登记(HA-BSR1、HA-BSR2、HA-BSR3、HA-BSR4、HA-BSR5、HA-BSR6、HA-BSR7、HA-BSR8)[75-77]。利用PAC2/RHA266构建的RILs群体,鉴定出位于14个连锁群(LG)上的36个盘腐病抗性QTLs,其中位于LG1、LG10和LG15的QTLs在多次田间试验中被检测到[78]。Pogoda等[79]利用由地方品种及自交系组成的218份多样性群体进行GWAS分析,鉴定出位于9个LG上的15个菌核病抗性QTLs,但遗传效应较小,而LG7和LG10上检测到多个QTL位点。Filippi等[80]对135个自交系进行关联作图分析(AMP),鉴定出13个抗性位点,其中4个自交系51084/C820、RHA801、HA441和51084表现明显的菌核病抗性,为育种提供了珍贵的抗性材料。由此可见,不同研究所鉴定的菌核病抗性位点不尽相同,一方面反映出菌核病的多基因遗传的复杂性,另一方面反映出环境条件对该性状影响较大。菌核病可发生在植株的多个部位,极易受湿度等环境条件影响。为避免田间自然发病不稳定性的影响,人们采用菌丝块、菌丝体悬浮液、孢子悬浮液、草酸浇灌等方法,分别接种于叶片(离体)、茎秆或花盘上进行人工鉴定[66,69,81],但目前尚缺乏一种规范可靠的抗性鉴定方法,导致缺乏准确的可重复的表型鉴定结果,也就难以找出真正的抗性资源和抗性基因。因此,建立科学可靠的表型鉴定方法,对重点野生资源、地方品种进行精准筛查,是菌核病抗性育种突破的首要环节。此外,组学技术为揭示菌核病抗性基因表达及抗病机理提供了新的工具和方法[82-84]。
2.2.2 茎溃疡病(phomopsis stem canker) 茎溃疡病其病原菌为DiaporthehelianthiMunt.-Cvetk., Mihaljc. and M. Petrov,其无性态为PhomopsishelianthiMunt.-Cvetk.etal,也称拟茎点霉病。该病于1980年在南斯拉夫首次被发现,目前已报道至少10个不同生理小种[85]。主要症状是茎秆和叶柄结合处有灰褐色病斑,逐渐变成暗灰或黑色[86](图4)。与菌核病类似,均属于腐生菌,二者发病原因相似,进化中有可能同时被选择。但不同于菌核病的是,茎溃疡病在1980s前并没有造成世界性的病害[87]。美国于1982年首次在德州报道,发病率仅为5%和11%[88]。但随后调查发现,自2001-2012年的十多年间,在美国北部该病发病率增加了16倍[89];而且主产区南达科他州、北达科他州/明尼苏达州地区2009-2015年发病率分别攀升至50%和56%。为此,美国将此病列入年度病害普查项目及优先研究领域[54]。此外,在大多数向日葵生产国包括塞尔维亚、法国、西班牙、罗马尼亚、俄罗斯等均有发生,是最具破坏性的真菌病之一。然而,中国对茎溃疡病研究几乎是空白,且多数文献报道中国迄今尚未见茎溃疡病[55,86,90]。但在陈为民等[46]报道中,新疆伊犁调查发现了11种主要向日葵病害,其中‘褐色茎腐病(Phomopsishelianthi)’,发病率高达65%,主要症状为叶片出现棕色病斑,叶柄形成灰色水浸状斑,最终造成茎部溃疡和髓部解离。根据上述症状,可以推测上述的‘褐色茎腐病’即为茎溃疡病。由于该病叶片或茎秆症状极易于与其他病害(比如黑茎病)相互混淆难以鉴定[89],加之命名上的不严谨或混乱,故而认为中国未见茎溃疡病的报道有待考究,在引用这些文献时,尤其是早期文献需十分谨慎,避免误导。

图4 茎溃疡病(茎秆上出现褐色病斑)[54]
目前,茎溃疡病已经成为许多主产国向日葵减产的主要病害之一,因此,抗性材料鉴定及基因挖掘日益引起人们高度重视。Gulya等[91]通过对1 106份多样性群体(499份来自前南斯拉夫,607份来自美国)的茎溃疡病自然发病率进行鉴定,筛选出23份材料(占2.08%)发病率低于10%,表现出较好的抗性。Talukder等[92]对260份多样性资源在2年3个地点鉴定,结果发现包括PI531389(Slovenska Siva)和PI531366(Lengyel A)在内的13份材料对茎溃疡病和盘腐病均表现出较好的抗性。Mathew等[93]在温室接种条件下,利用2个小种D.helianthi和D.gulyae对49份资源进行鉴定,与对照HA288相比,其中PI552939对2个小种均表现较好抗性。可见,虽现有资源中尚未发现高抗和免疫材料,但不同材料间存在一定的抗性差异。
相比于菌核病,茎溃疡病发现较晚,有关该病的遗传解析及基因鉴定研究较少。已有研究表明茎溃疡病是由多基因控制的数量性状[94]。Bert等[95]利用双亲群体检测到15个QTLs,解释表型变异的7.2%~34.7%。Langer等[96]在RILs群体中检测到8个QTLs,其中位于LG15的QTL解释46%的表型变异。利用HA89(感) 和HA-R3(抗)构建的RIL群体,共鉴定出位于11个LGs上的15个QTLs,遗传效应为5.24%~17.39%[97]。茎溃疡病与菌核病均属于腐生菌,一旦爆发对产量将造成毁灭性影响,可导致40%以上的减产[54]。鉴于命名混乱、田间症状易混等原因,中国对此病尚缺乏系统研究,因此,开展准确可靠的表型鉴定和病害预警事关向日葵产业安全,亟待高度重视。
2.2.3 黄萎病(verticillium wilt) 黄萎病主要由大丽轮枝孢菌(VerticilliumdahliaKleb)病原菌引起。典型症状是叶片出现黄色边缘的斑块,由下部叶片逐渐向上部发展,并通过维管束传播,导致维管束组织褐化并萎缩变黑。黄萎病在中国向日葵产区均有发生,且发病面积有逐年增加的趋势。调查发现,新疆黄萎病发病率约10%,严重时达70%;宁夏灌区普遍发生严重,一般在40%左右;而内蒙古主产区发病率通常在10%~45%[45,49,98-101]。针对黄萎病,目前尚无有效的杀菌剂[102],是影响向日葵产量的主要病害之一。
对黄萎病的遗传研究可以追溯到1950s,Putt[103]在加拿大育种材料中首次发现了黄萎病抗源,并指出黄萎病由显性单基因V1控制。随后Miller等[104]发现,HA89(油葵保持系,广泛用于群体构建)含有同一抗性基因V1,并利用HA89选育和登记了首批食用向日葵抗黄萎病自交系HA312、HA313、HA314和HA315。可见,50年来抗性品种选育一直是控制黄萎病的最有效措施,直到出现了新的生理小种。Martín-Sanz等[105]发现,HA89对东欧V2-EE小种表现抗性,但对西欧V2-WE小种表现感病;而另一自交系INRA2603的抗性与HA89正好相反。新的小种演变从一个侧面也解释黄萎病日趋增加的原因。
中国十分重视黄萎病的研究,包括病原菌致病力、黄萎病发生规律及防治措施、抗性材料筛选等方面,并已成功鉴定出一些高抗的食葵资源,如免疫品种SH338、SH361、JK601和LS009;高抗品种JK103、JK107、JK108、‘巴葵138’和‘甘10138’[100,106-108]。此外,利用RNA-seq等组学技术解析黄萎病抗性机理也取得一定进展[102]。鉴于黄萎病属于单基因遗传,而且现有资源中已鉴定出免疫或高抗种质,未来新品种选育中应重视抗黄萎病基因的协同转育,避免该病的爆发和流行。
2.2.4 锈病(rust) 锈病由病原菌锈菌(PucciniahelianthiSchwein.)引起,典型症状是叶片产生铁锈色病斑,是全世界普遍发生的病害之一。锈病最早发现于1960s[52]。但是,在最初的几十年间,由于大多数油葵和食葵品种均具有良好的抗性,锈病并没有给生产上造成严重危害。直到1990s早期,新的锈病小种出现致使原有品种失去了抗性。目前,共鉴定出39个锈病生理小种,广泛分布于北美洲、澳大利亚、阿根廷、南非、俄罗斯、印度和中国等地区,其中334和336是主要优势小种,而777是目前毒力最强的小种。国际上已建立了一套共9个向日葵鉴别系(含不同抗性基因)用于区分不同生理小种[109]。
锈病一直是美国最流行的病害之一,某些地区超过50%的田块发生病害[54]。在中国,锈病主要发生在吉林、辽宁、黑龙江、内蒙古和新疆等地,主要生理小种为100、300、500和700。在新疆阿勒泰地区,2016年被调查的田块中约70%发生锈病,病株率达81.2%~100%[110]。而且,食葵品种比油葵更易感病[111],随着食葵大规模种植,一旦气候条件适宜,极易导致锈病的爆发,因此锈病抗性品种选育不容忽视。
已有研究表明,锈病由单基因控制,部分栽培品种或野生种,如H.annuus、H.petiolaris和H.argophyllus等资源中均含有抗性基因[112]。目前已鉴定出17个抗性基因,包括R1-R5、R10-R12、Pu6、Radv、R13a、R13b和R14-R18,分别定位于5个连锁群(R3和R10除外,未鉴定)。值得注意的是,其中9个抗性基因均位于LG13的2个基因簇中[109,113]。据报道,油葵自交系RHA397(含R13b)和TX16R(含R16),以及食葵自交系HA-R6(含R13a)均抗所有生理小种[109],这就为食葵锈病抗性育种提供了重要的种质资源。
2.2.5 霜霉病(downy mildew) 霜霉病由病原菌Plasmoparahalstedii(Farl.) Berl. & de Toni引起,苗期表现萎缩和矮化,节间缩短,叶片出现褪绿黄斑,叶背面出现白色霉层[114]。霜霉病是一种最古老、遗传机制研究最清晰,抗性育种利用最有效的一种病害,但偶尔仍造成毁灭性的影响。霜霉病最早于1882年在美国东北部发现,并对此开展了一系列研究[115]。中国于1963年首次在黑龙江发现霜霉病[116]。迄今为止,全世界霜霉病已发现40个生理小种,遍布世界所有向日葵种植区,其中欧洲和美国尤为严重[117]。在美国连续12年的全国病害调查中,霜霉病属于重点调查的病害之一。2005和2011年美国被调查的田块40%发现了霜霉病,病株率3%~12%[54]。鉴于大量新的小种出现,Gulya等[118]建立了包含9个自交系的一套霜霉病标准鉴别系。之后鉴别系不断更新和完善,目前分别为Mycogen 270(CK)、RHA265、RHA274、DM-2、PM17、803-1、HA-R4、HA-R5和HA335[119]。
绝大多数情况下,霜霉病主要由单显性基因(Pl)控制,这就使得早在1960s抗霜霉病品种已成功用于商业生产[112]。迄今为止,已鉴定出36个Pl基因(Pl1—Pl35、PlArg),其中30个已分别定位到6个连锁群上(LG1、LG2、LG4、LG8、LG11、LG13),部分成簇存在。几个重要基因位点包括Pl17、Pl18、Pl19、Pl20的关键候选基因已被成功克隆[117,120-121],加深了人们对霜霉病抗性机理的认识。
在中国,除个别地区个别年份外,一般情况下霜霉病仅零星发生,利用种衣剂处理可以有效地控制霜霉病[122],未见造成大面积毁灭性灾害的报道。因此,中国对霜霉病的研究相对较少,抗性育种一直未引起足够重视。王富荣等[122]通过对247份油葵和食葵品种进行鉴定,发现28份油葵表现高抗,但食葵中无抗性品种。随着大面积非抗品种的推广,若遇气候条件适宜,霜霉病的发生仍存在极大隐患[113-114]。
众所周知,油葵和野生资源含有丰富的抗性基因。研究表明,49个野生向日葵种中仅有11个种出现感病,是最有效的抗性来源[115]。利用油葵或野生抗源基因,一系列兼抗霜霉病或锈病的食葵自交系,包括HA-DW1(Pl18)、HA-DM2(PlArg/R12)、HA-DM3(Pl17/R13a)、HA-DM4(Pl18/R13a)、HA-DM5(Pl19)、HA-DM6(Pl35)、HA-DM7(Pl20)和HA-DM8(Pl35)已被成功转育并登记[123-126],为食葵抗霜霉病育种提供了重要的抗性资源。
2.2.6 黑茎病(phoma black stem) 黑茎病其病原菌有性态为LeptosphaerialindquistiiFrezzi,无性态为PhomamacdonaldiiBoerma。1964年该病首次在加拿大被鉴定和报道,目前已经遍布世界各地,其中法国尤为严重[127]。1990年法国发生大面积黑茎病[128],是仅次于霜霉病之后的第二大病害,减产10%~30%[129]。2005年,中国首次在新疆采集并鉴定出黑茎病[130-131],2010年列入入境检疫性有害生物名录。通常情况下该病仅零星发生,但遇多雨高湿条件可造成大面积发生,新疆北部、黑龙江、内蒙古等省区都有发生。2007年,新疆新源县发病田块约占51%,田间发病率50%[128,132]。中国对黑茎病研究主要集中在病原菌生物学特性、致病性、发生规律、防治措施等方面,对于抗性育种未见报道[128,133]。研究认为,向日葵不同品种间抗病性存在一定差异,绝大多数资源表现高感,一些品种具有部分抗性,但未见免疫或高抗品种[127,133-134]。利用PAC2/RHA266的F9代重组自交系,人工接种幼苗鉴定出具有中等抗性的QTL,贡献率6%~20%,表明其微小多基因遗传机制[129]。因此,建立和优化科学可靠的表型鉴定方法,如生长箱接种、温室接种、田间自然观察等,对抗性基因的准确鉴定至关重要[135]。
2.2.7 其他病害除上述病害外,褐斑病、黑斑病等在一定程度上的发生同样会影响向日葵生产。褐斑病(斑枯病)遗传抗性研究极少[136-137],早期少数研究表明,一些食葵农家品种如‘九连灯’、‘太原80天’等,在田间人工接种条件下表现免疫或高抗[138],但近期相关研究报道很少。黑斑病是与其类似的另一类叶斑病,其病原菌为向日葵链格孢菌Alternariahelianthi(Hansf.) Tubaki et Nishihara。1966年在吉林首次发现,目前在向日葵产区普遍发生,可造成严重减产甚至绝收[50]。牛庆杰等[139]曾利用野生种开展远缘杂交,获得了耐黑斑病后代个体,田间及接种的发病率较对照明显降低,但目前尚未发现高抗或免疫品种[50,140-141]。有关上述两种病害的抗病遗传机理报道较少,故不在此展开。植物病害研究需要长期积累,跟踪调查病原菌的演化,否则一旦爆发以往的防治措施或抗性品种可能失去效果而造成严重影响。
2.3 抗列当及抗除草剂育种
向日葵列当(OrobanchecumanaWallr.)为一年生专性寄生植物(图5),属于高度危险级别的检疫性有害生物,其综合风险评价值R为2.473[142]。中国于1959年首次在内蒙古乌兰察布发现列当,寄生于蒿属植物根上[143]。其后1979年在对吉林白城地区向日葵病害调查中也发现列当大量寄生[144],据此推测列当在中国早有发生,确切来源及发生时间无从考究[145]。由于列当与向日葵寄主的协同演化,列当出现明显的生理小种分化,国际上已鉴定出A、B、C、D、E、F、G、H等不同小种。目前,中国向日葵列当已蔓延至所有向日葵产区,且绝大多数地区出现了高级别的F和G小种[146]。国际上列当生理小种的命名尚未完全统一,G生理小种还有待验证。中国以食葵种植为主,食葵整体上抗性比油葵差,可能是造成列当快速蔓延的原因之一[147]。

A.内蒙古武川田间列当;B.温室盆栽条件下的列当
向日葵对不同生理小种列当的抗性主要由显性单基因控制,分别为Or1—Or7,而野生向日葵中含有丰富的抗性基因。单基因控制以及丰富的抗性基因来源(野生种)使得抗列当品种选育一直较为成功。早在1920s前后,国外已育成抗不同小种的油葵常规品种,如Kruglik A-41(抗A),J8281(抗B)、VNIIMK 1646(抗B)和Record(抗C)等[148-150]。国内的一些食葵杂交种如‘谷丰1号’、TP3313、TP3316、JK601也具有良好的列当抗性[151-152]。由于抗列当品种推广,人为形成高选择压力,使得高毒力小种成为优势小种,导致原有的抗性品种逐渐失去抗性[146]。2018年,在中国78个被调查的田块中,60个田块均发生了列当;其中内蒙古最高寄生率高达100%,寄生强度达38.5;新疆次之[147]。且内蒙古地区的列当65%以上为高毒力的F和G小种[146]。列当寄生可导致减产30%~60%甚至更高[153]。因此,近几年来,各种抗列当品种受到市场青睐,如‘双星6号’、‘三瑞11号’、‘同辉33’、HZ2399、‘乐丰3.0’等。对于列当重发区,抗列当成为种植户选择的首要因素,甚至对品种的产量、品质指标均降低了要求。可以预计短期内抗列当仍然是种子销售的“指挥棒”。
由于列当生理小种的快速演变,不仅导致抗列当品种使用期限缩短,并且出现一些新的小种。据报道,新疆列当种群与国内其他地区列当差异较大,具有最多的特有等位基因和最丰富的多态位点,暗示新疆列当可能是向日葵列当与当地其他列当科物种杂交的结果,使得新疆列当种群在某些向日葵品种(如LD5009)上寄生数量远高于其他品种[145]。这一现象也被我们反复多次的盆栽接种试验所验证,即新疆列当种群与内蒙古列当种群存在明显的差异(未发表)。这就意味着抗列当品种的推广存在一定的潜在风险,而利用非转基因途径选育抗除草剂品种应该是一个有效的替代途径[153]。利用除草剂对列当进行防除不受生理小种的局限,抗咪唑啉酮类(IMI)的Clearfield(2003年商业化)及换代产品Clearfield Plus早已在油葵中广泛应用[150],而食葵中除草剂的应用几乎是空白。利用成熟的分子标记技术快速转育兼具Clearfield Plus或Syngenta AIRTM性状的新品种[154-155],有望成为开发抗除草剂食葵领域新的增长点。
2.4 抗籽粒锈斑育种
籽粒锈斑(Seed scarring)即指向日葵成熟后果壳表面出现的浅灰色斑痕(图6),严重影响向日葵的商品性。目前,籽粒锈斑主要发生于食葵,尤其是核心种植区内蒙古河套灌区,其他地区较轻(比如内蒙古赤峰、新疆),油葵中未见有报道。内蒙古河套灌区自20世纪70年代末大面积种植向日葵,至今已有近40年的历史,种植面积一直居全国首位[28]。由于气候干旱、土壤盐渍化严重,除少部分地块可轮作玉米外,多数中低产田连续种植向日葵,轮作倒茬困难,导致病虫草害逐年加重,尤其是向日葵籽粒锈斑病已成为近年来制约河套灌区向日葵产业的最大障碍。目前有关向日葵籽粒锈斑病的研究报道较少。
初步研究认为,开花期蓟马(图6)锉吸幼果可能是造成籽粒锈斑的主要原因[156-157]。蓟马对果蔬、农作物、花卉等花果外观品质的危害已有较多报道,例如,柑橘中的花皮果或风癣果[158]、苹果白斑症[159]等。目前,喷施杀虫剂仍然是控制蓟马的唯一有效措施。然而,摆脱对农药的依赖是食葵走向品质化及绿色健康的必然要求。因此,从长远目标看,培育籽粒锈斑抗性品种仍将是根本上减少籽粒锈斑危害的最经济有效的途径,但目前在向日葵中尚未见相关的研究工作。包选平等[160]通过对大豆品种‘齐黄34’(感)与‘冀豆17’(抗)的重组自交系群体进行遗传分析,初步在大豆中鉴定了3个抗黄蓟马(Thripsflavus)的遗传位点,其中rtf1位点贡献率达15.8%。由于不同品种的叶片厚度、蜡质层、茸毛等性状存在一定差异,有可能影响黄蓟马取食的偏好性。特木尔布和等[161]对紫花苜蓿形态特征及抗虫特性研究表明,不同苜蓿品种对蓟马存在抗性差异。抗蓟马品种具有叶色深绿、叶片厚、茎叶茸毛密度大且短粗的特点,进而影响蓟马采食和生存性。抗虫机理的研究为向日葵籽粒锈斑病的探索提供了重要的参考和启示。向日葵包括野生和栽培共51个种,具有十分丰富的原始基因库。因此,筛选潜在的籽粒锈斑抗性材料,探讨籽粒锈斑遗传机理,选育符合市场需要的籽粒锈斑抗性新品种,真正实现食葵品质化和健康化。

A.瘦果表面出现浅灰色斑痕;B. 舌状花上的蓟马
2.5 品质育种
食葵消费主要以嗑食直接消费为主,对市场需求敏感。消费者普遍喜欢大粒、光泽度好、籽仁饱满、口感香脆的葵花籽,并在很大程度上引导着商品的收购价格。由于强烈的市场效应,使得育种家一直追求粒大、外观好的品种,而籽仁营养品质始终被忽视。向日葵油因富含不饱和脂肪酸(油酸、亚油酸等)和维生素E一直被公认为是仅次于橄榄油的高品质健康油。因此,高油酸育种及维生素E的多元化改良早已成为油葵育种的热点,而这一育种目标同样适用于食用向日葵,但目前几乎是空白。面对市场同质化严重、商品销售难的问题,未来应将外观品质与内在营养品质协同考虑,将营养健康、专用型指标放在首位,使优质营养成为食葵育种研发新的风向标。
2.5.1 油酸和亚油酸向日葵籽仁主要由脂类、蛋白质和多糖类物质组成。其中脂类又分为饱和脂肪酸(棕榈酸、硬脂酸等)和不饱和脂肪酸(油酸、亚油酸等)。油酸(oleic acid,C18∶1,属于omega-9)是一种单不饱和脂肪酸,氧化稳定性高,天然存在于动植物油脂中。亚油酸(linoleic acid,C18∶2,属于omega-6)和亚麻酸(linolenic acid, C18∶3,属于omega-3)为多不饱和脂肪酸,二者均为人体无法合成的必需脂肪酸,但氧化稳定性差。由于双键数目不同,油酸∶亚油酸∶亚麻酸氧化速率约为1∶10∶20[162-164]。这意味着亚油酸的氧化速率是油酸的10倍左右,降低亚油酸和亚麻酸的含量将有利于提高油脂的氧化稳定性,延长货架期;而且可以降低高温烹饪时产生的不良化学成分,有利于食品工业高温煎炒。因此,提高油酸的相对含量,增加油酸/亚油酸(O/L)比值,早已成为油葵育种的主要目标[163]。前苏联早在1970s就开始了高油酸向日葵的开发,美国1984年高油酸向日葵已开始商业化规模化种植[165],1997年正式推出的高油酸努桑油(NuSun)以无氢化饱和脂被视为美国日常用保健油,2000年已占市场份额的15%。此外,近20年来,法国在高油酸育种领域取得了重大进展,约60%以上的品种为高油酸,其中2019年高油酸品种占76%。在乌克兰和俄罗斯,2018年,高油酸品种推广面积约42万hm2[166]。提高油酸含量也一直是中国油葵遗传改良的重要目标之一。利用高油酸和低油酸品系构建的F2分离群体,克隆了3个控制油酸的QTL,分别位于LG9(表型贡献率12.05%)和LG6(表型贡献率5.81%、5.18%)[164]。利用无损的近红外技术预测向日葵籽仁品质成份(如粗脂肪、蛋白质、油酸、亚油酸等),为品质育种前期大量优异种质筛选提供了快速测定技术[167]。相比之下,人们对食葵油脂组份特征研究较少。研究发现,油用向日葵中油酸含量相对较高,相反地,绝大多数食葵中亚油酸含量较高,达50%以上[168-169]。因此,相比油葵,食葵更易氧化,产生油脂酸败,尤其是炒制的葵花籽(高温加速氧化)。通过调控脂肪酸组成,延长食葵保质期尤其炒制葵花籽保质期就显得十分必要,因此选育高油酸食葵品种值得高度重视。
2.5.2 维生素E维生素E(Vitamin E)是生育酚及生育三烯酚的统称,分别包括4种生育酚(α、β、γ和δ-生育酚)和4种生育三烯酚(α、β、γ和δ-生育三烯酚)。维生素E只能由光合生物合成,是最重要的天然抗氧化剂之一。维生素E的含量和组成也是油脂品质评价的重要指标。维生素E是保护油脂的风味、延长油脂储藏时间、增强油脂营养品质的关键因子之一。在抗氧化、抑制胆固醇合成及肿瘤细胞生长、改善动脉粥样硬化及预防心血管疾病等方面具有独特的生理功能(辅助药物)。葵花籽油中α-生育酚含量最高,占维生素E总量的96.7%[162,170]。但α-生育酚抗氧化能力最低,因此,提高β、γ和δ-生育酚含量,提高植物油的氧化稳定性,一直是育种的重要目标之一。研究发现,野生向日葵种子生育酚平均含量328 mg/kg,其中99%为α-生育酚,0.7%β-生育酚,0.3%γ-生育酚,也检测到少数野生种含有较高的β-生育酚(eg.11.8%,H.debilis)或γ-生育酚(14.6%,H.nuttallii);栽培向日葵种子生育酚平均含量669 mg/kg,其中,92.4%α-生育酚,5.6%β-生育酚,2.0%γ-生育酚[171]。通过化学诱变(EMS)或生物技术途径,改变各类生育酚的含量(如提高γ-生育酚含量至940 mg/kg),进而提高其氧化稳定性是完全可行的[171-173]。
总之,随着植物脂肪酸或维生素E合成代谢通路的解析以及关键酶基因的克隆,借助基因编辑技术,集高油酸、高亚油酸、低亚麻酸、高β-或γ-型生育酚等众多优良品质于一身的油用或食用向日葵育种一定会是未来品质育种的热点[163,174]。
3 种质资源创新利用
食葵育种历史较短,育种基础群体遗传背景相对单一,育种主要目标是高产和优质。因此,相比油葵,食葵抗性总体较差。如何从大量的种质资源中挖掘有利抗性基因,推出多抗优质新品种,才能确保在无法预测的各种病虫害环境下高产稳产。中国收集保存有3 200余份种质资源,主要以国内收集的系选种和农家种为主,其中食葵种质约占72%,油葵及油食兼用占27%,野生种质较为缺乏(约20份)[175]。在2017-2019年间登记的品种中,68.3%食葵品种是利用国外亲本或杂交后代选育而来,国外品种主要集中于从美国、以色列、西班牙等国引进的LD5009、3638C等10余份材料,而国内材料主要涉及‘三道眉’、‘星火花葵’等农家品种,育种基础群体的遗传背景相对狭窄[37]。
近几年,一些育种单位或种业公司加强了种质资源的收集和利用,包括野生种、地方品种以及栽培品种(图7),极大丰富了向日葵种质资源。然而,对种质资源普遍存在重视收集保存,但创新利用不足。尤其对野生种利用,因杂交困难、回交转育周期长、不良性状连锁等不利因素,难以被育种家所重视。然而,野生资源正是菌核病、列当等抗性基因的原始来源。因此,加强对现有种质资源系统性的多年多点的表型鉴定,尤其是野生资源及农家品种的利用;同时,加强国内外不同育种家之间材料交换,将不同种质的优点聚合起来,提高育种基础群体的异质性,将是未来选育突破性杂交种,或联合开发适于更广泛地域的品种,推动食葵产业持续发展的核心和保障。

A.多样性地方品种;B.野生向日葵
4 生物育种技术融合及引领
近20年来中国食葵育种取得了巨大成就,引领着食葵产业的发展方向。但就育种方法来讲,早期主要借鉴前苏联的半分法(half division method)、集团选择法(mass selection)、或轮回选择法(recurrent selection),用于基础育种群体的改良,对常规品种选育非常有效;其他常用方法如系谱选择法(pedigree selection)主要用于亲本自交系的选育[4,16,176]。从育种技术上,主要以常规育种(表型选择)为主[37],分子育种技术远未普遍应用。尤其在优异基因鉴定,如抗病性筛选仍主要依赖表型鉴定,相关的分子标记开发等方面比较薄弱,仅有少数研究报道,比如:利用回交或RIL群体鉴定抗旱QTL[177-178];采用抗感极端池全基因组测序(BAS-Seq)方法鉴定耐盐基因[179];或利用SLAF-seq技术开发油葵SNP(single nucleotide polymorphism)标记[180]。随着转录组学兴起,大量研究集中于特定功能基因的组织表达分析[181-188]。
目前国际上公开发表的SNP标记主要是基于油用向日葵开发的,食葵分子育种体系欠缺。三瑞农科与华智生物合作,开发了一套适于食葵的覆盖全基因组的1500余个高质量的KASP (kompetitive allele specific PCR)标记。该标记可用于回交转育中背景选择、功能标记开发、核心品种指纹图谱构建、品种保护及种子生产中的纯度鉴定等(未发表)。其次,应加快开发和利用与重要性状相关的SSR/SNP分子标记,包括油葵中已成功应用的,如抗列当、抗黄萎病、抗锈病等功能基因标记,使分子标记辅助育种成为常规手段,并借助幼胚培养技术,加速现有骨干品种的改良升级,实现分子育种与常规育种的有机整合。
5 结 论
中国作为食葵生产大国,在高产、优质育种方面取得了巨大成就,总体上实现了4次品种更新:从自留自种农家种到规模化种植常规种,过渡到美葵为主的国外杂交种垄断市场,最终实现自主选育优质杂交种,完成了食葵品种的国产化。随着育种目标从注重高产到追求商品品质,食葵育种总体进入了第5次品种更新,优质营养、多抗、宜机收的新品种将得到市场青睐。在现阶段育种工作中,应充分挖掘和利用外来种质或原始优良种质资源,提高育种基础群体的异质性,为突破性品种的选育奠定基础。以食葵为目标的SNP标记开发,极大地有助于重要性状基因转育和聚合,促进常规育种与分子育种的有机结合,加快核心品种更新换代的进度。随着品种审定登记制度进一步完善,以及DNA指纹数据库建立和真实性鉴定技术的实施,对于仿冒套牌等现象的市场监管更加严格和规范,中国食葵产业进入良性有序发展阶段。
国内外食葵市场前景广阔,但国内食葵种植面积基本稳定。因此,加强科企合作,加快品种改良和技术更新,提高单产潜力对产业发展至关重要。未来,根据地域和市场细分由单一品种发展到多元化,包括生货和炒货、高端和通货、以及专用特用品种。这一强大的消费市场效应,促使育种、生产、加工等各个环节进一步规范化,商品以质论价,长粒、黑底白边、色泽亮、口感香脆品种,成为高端消费市场的风向标。尤其是依托国家对油料作物政策上的大力扶持,使中国向日葵产业实现了育种研发、生产繁育、订单种植、炒货加工、出口贸易等全产业链的融合发展新模式。真正使向日葵产业从单纯种子研发和销售,过渡到高附加值的产品开发,实现葵花籽就地转化升值,带动企业和农户增收,促进整个食葵产业的高质量发展。
致谢:感谢吉林省白城市农业科学院向日葵研究所牛庆杰研究员在成稿过程中无私交流及宝贵建议!感谢三瑞农科科学院同事大力支持!
