不同果锈等级的阳光玫瑰葡萄果皮结构及相关物质变化
2022-11-25娄玉穗尚泓泉樊红杰吕中伟张晓锋吴文莹
娄玉穗,尚泓泉,樊红杰,李 政,张 柯,吕中伟,张晓锋,吴文莹
(1. 河南省农业科学院 园艺研究所,河南 郑州 450002;2. 河南中远葡萄研究所有限公司,河南 郑州 450045)
阳光玫瑰是目前我国最受欢迎的葡萄品种之一,然而该品种在成熟期会出现果锈症状,严重影响其外观品质和商品性[1⁃2]。果锈是一种生理性病害,表现为在果实表面呈现条状或不规则状黄褐色锈斑,严重时连成片[3⁃4]。果锈常发生在梨、苹果、枇杷和黄绿色的鲜食葡萄品种上[5⁃7]。
组织学上观察发现,发生果锈的苹果和梨果实表皮细胞角质层破裂,木栓化程度加深,表皮细胞甚至皮层细胞死亡,且果锈形成的微观结构变化早于外观变化[8⁃10]。与抗锈苹果品种相比,感锈品种的果实表皮细胞排列松散、无序、细胞壁薄,且发生果锈部位的表皮角质层和蜡质层薄,没有规则,易破裂[8]。SUEHIRO 等[3]在显微镜下观察发现,阳光玫瑰葡萄果锈的褐变部位位于表皮细胞组织内,褐变物质位于果皮的细胞间隙和细胞壁上,而关于阳光玫瑰葡萄果锈产生过程的组织结构变化尚未见报道。王慧等[4]在翠冠梨上的观察发现,果锈主要是破碎的角质膜、木质化加厚的表皮和亚表皮细胞以及果点上较易剥落的物质。张鹏飞[11]的研究表明,与无锈梨相比,有锈梨果皮木质素含量升高,角质含量减少,木栓质含量增加。还有研究表明,赤霉素和氯吡脲处理降低阳光玫瑰葡萄果锈发生程度与果皮中木质素和总酚含量的降低有关[12]。另外,在苹果和梨的研究中还发现,果锈发生伴随着苯丙烷代谢、酚类物质代谢等次生代谢的发生[13⁃14],且苯丙氨酸解氨酶与过氧化物酶是梨果皮木质素合成关键酶[15]。HOU 等[16]研究表明,赤霉素和氯吡脲处理会降低阳光玫瑰葡萄苯丙氨酸解氨酶活性,从而降低果锈的发生。因此,研究不同果锈等级的阳光玫瑰葡萄果皮组织结构、细胞膜透性、果皮结构物质及相关酶活性变化,旨在探明阳光玫瑰葡萄果锈形成的生理机制,为果锈防治技术开发奠定基础。
1 材料和方法
1.1 试验材料
试验于2019 年在河南现代农业研究开发基地(35.0°N、113.7°E)进行。以8年生阳光玫瑰葡萄植株为试验材料,贝达砧,南北行向,株行距1.5 m×3.0 m,高宽垂架式,简易避雨栽培。花果管理和肥水管理参考娄玉穗等[17]的方法进行,于见花时修整花穗,留穗尖6 cm,盛花后1~3 d(2019 年5 月11—13 日)分批次用25 mg/L GA3(赤霉酸)+2 mg/L CPPU(氯吡脲)进行保花保果处理,14 d 后用25 mg/L GA3进行膨大处理。在果粒黄豆粒大小时疏果至60粒左右,控制单穗质量700 g左右。
1.2 果锈发生率的调查
从果实进入软化期后(8月7日,盛花后90 d)开始调查阳光玫瑰葡萄果锈发生率,每10 d 调查一次,直至10 月16 日(盛花后160 d),每次调查15 串果穗。调查方法:调查并记录每串果穗上有果锈的果粒数量和果穗上总果粒数量,计算果粒果锈发生率,果锈发生率=有果锈果粒数量/总果粒数量×100%。
1.3 果皮组织结构的观察
果实进入软化期后(8月15日、9月20日),采集有果锈和无果锈的果粒样品,带回实验室。按照果锈发生程度将果粒分成轻度果锈(果锈面积≤5%)、中度果锈(果锈面积6%~25%)和重度果锈(果锈面积>25%)3 个等级。利用体视显微镜(M205A,Leica)观察不同等级的果粒果皮组织结构。光学显微镜观察:取不同果锈程度的果粒,用刀片在果皮上切1 cm2左右,然后用镊子轻轻夹起果皮,放到载玻片上,滴上一滴蒸馏水,盖上盖玻片,放到显微镜(ECLIPSE E100,Nikon)下观察果皮组织结构特征[18]。
1.4 果皮细胞膜相对渗透率、细胞壁成分与果锈物质含量及相关酶活性的测定
在果实成熟期(9 月16 日),采集有果锈和无果锈的果粒样品,参考冯娇等[19]的方法对果粒进行等级划分,即0级:没有果锈;1级:果锈面积≤5%;2级:果锈面积6%~15%;3 级:果锈面积16%~25%;4 级:果锈面积>25%。
1.4.1 果皮细胞膜相对渗透率 采用电导仪法测定果皮细胞膜相对渗透率。首先用去离子水清洗果粒,用吸水纸吸干,然后用直径为8 mm 的打孔器打取不同果锈等级的果皮,用镊子夹取果皮放到吸水纸上吸取果皮内侧果汁,每个等级的果皮样品打取20 个小圆片,接着放入试管中,加入15 mL 去离子水,室温下摇床振荡浸泡1 h,测定电导率;之后在100 ℃沸水浴中煮沸10 min,冷却至常温后测定电导率;计算果皮细胞膜相对渗透率,细胞膜相对渗透率=煮沸前的电导率/煮沸后的电导率×100%。
1.4.2 果皮细胞壁成分与果锈物质含量及相关酶活性 用镊子分别夹取5 个果锈等级的果皮样品,然后用液氮进行快速冷冻,之后用冷冻研磨仪(A11 basic,德国IKA/艾卡)进行研磨粉碎,粉碎样品用于以下指标的测定。纤维素含量采用蒽酮比色法测定,半纤维素含量采用DNS 比色法测定,原果胶含量采用咔唑比色法测定,木质素含量采用紫外分光光度法测定,总酚含量采用Folin-Ciocalteu 试剂法测定[19]。纤维素酶活性采用蒽酮比色法测定,酸性磷酸酶活性采用对硝基苯磷酸二钠水解法测定,果胶酶、碱性磷酸酶和苯丙氨酸解氨酶活性采用吸光法测定,过氧化物酶活性采用愈创木酚法测定,多酚氧化酶活性采用邻苯二酚法测定[20]。
1.3 数据处理
试验数据用Excel 2010 软件进行整理和绘图,用SPSS 20软件进行统计分析。
2 结果与分析
2.1 不同挂树时间对阳光玫瑰葡萄果锈发生率的影响
如图1所示,随着果实的成熟,果锈发生率呈现先增加后趋于稳定的变化趋势。在所调查的时间中,盛花后90 d 的阳光玫瑰葡萄果锈发生率最低,为3.3%;盛花后100 d和110 d的果锈发生率之间差异不显著,且盛花后100 d 的果锈发生率显著低于盛花后120 d 及之后时间调查的果锈发生率;盛花后130 d 的果锈发生率达到最大值,为20.0%,其与盛花后120 d 及之后时间的果锈发生率之间差异均不显著。

图1 不同挂树时间对阳光玫瑰葡萄果锈发生率的影响Fig.1 Effect of days after full bloom on berry russet rate of Shine Muscat grape
2.2 阳光玫瑰葡萄果锈组织结构特征
2.2.1 体式显微镜的观察结果 从图2 可以看出,轻度果锈仅在果皮表面出现褐色不规则斑块,细胞壁褐化,使得细胞结构凸显(图2A、2B);中度果锈的果皮上褐色斑块增大,果锈位置呈现凹凸不平的状态(图2C、2D);重度果锈的果皮褐化程度加深,甚至呈现黑褐色,果皮上凹凸不平的组织连成片,部分细胞全部褐化坏死(图2E、2F)。

图2 体式显微镜下阳光玫瑰葡萄果皮果锈特征Fig.2 The characteristics of berry russet of Shine Muscat grape under stereomicroscope
2.2.2 光学显微镜的观察结果 从图3 可以看出,没有发生果锈的果皮细胞排列整齐,细胞颜色均匀,细胞结构完整(图3A)。在果锈发生初期(软化期),轻度果锈的果皮细胞壁和细胞间隙呈褐色,细胞内部颜色不变(图3B);中度果锈的果皮褐化范围增大(图3C)。在果锈发生后期(成熟期),轻度果锈的果皮褐色物质向细胞内部蔓延且凹陷,细胞间隙增大,褐化范围增加(图3D);中度和重度果锈的果皮上褐化细胞数量增多,且部分细胞内部全部褐化,细胞结构遭到破坏(图3E、3F)。

图3 光学显微镜下阳光玫瑰葡萄果皮果锈特征Fig.3 The characteristics of berry russet of Shine Muscat grape under optical microscope
2.3 不同果锈等级对阳光玫瑰葡萄果皮细胞膜相对渗透率的影响
如图4所示,随着果锈等级的增加,果皮细胞膜相对渗透率增大,说明细胞膜受到破坏的程度增大。果锈等级0、1 和2 的果皮细胞膜相对渗透率分别为67.9%、69.8%、70.8%,三者之间差异不显著,且均显著小于果锈等级3 和4 的果皮细胞膜相对渗透率;果锈等级3 和4 的果皮细胞膜相对渗透率分别为77.1%和80.4%,两者之间差异不显著,说明果锈等级3及以上的果皮细胞膜受到较严重破坏。

图4 阳光玫瑰葡萄果锈等级与果皮细胞膜相对渗透率之间的关系Fig.4 Relationship of berry russet level and relative permeability of cell membrane in Shine Muscat grape peel
2.4 不同果锈等级对阳光玫瑰葡萄果皮细胞壁和果锈相关物质含量的影响
从表1可知,随着果锈等级的增加,阳光玫瑰葡萄果皮的纤维素、半纤维素、原果胶含量呈现逐渐降低的变化趋势,而木质素和总酚含量呈现逐渐升高的变化趋势。其中,没有发生果锈的果皮纤维素含量为330.7 mg/g,其显著高于所有发生果锈的果皮纤维素含量;果锈等级1 和2 的果皮纤维素含量分别为315.8 mg/g 和309.3 mg/g,两者之间差异不显著,且果锈等级1 的果皮纤维素含量显著高于果锈等级3 和4 的果皮纤维素含量;果锈等级3 和4 的果皮纤维素含量之间差异显著。在果皮半纤维的含量上,果锈等级0 和1 的半纤维素含量之间差异不显著,且两者均显著高于果锈等级2、3 和4 的果皮半纤维素含量;果锈等级2 和3 的果皮半纤维素含量分别为64.3 mg/g 和61.3 mg/g,两者之间差异不显著,且均显著高于果锈等级4的果皮半纤维素含量。果锈等级0 的果皮原果胶含量为74.9 mg/g,其显著高于果锈等级3 和4 的果皮原果胶含量;果锈等级1、2 和3 的果皮原果胶含量之间差异均不显著,且三者均显著高于果锈等级4 的果皮原果胶含量;果锈等级4的果皮原果胶含量为62.9 mg/g。
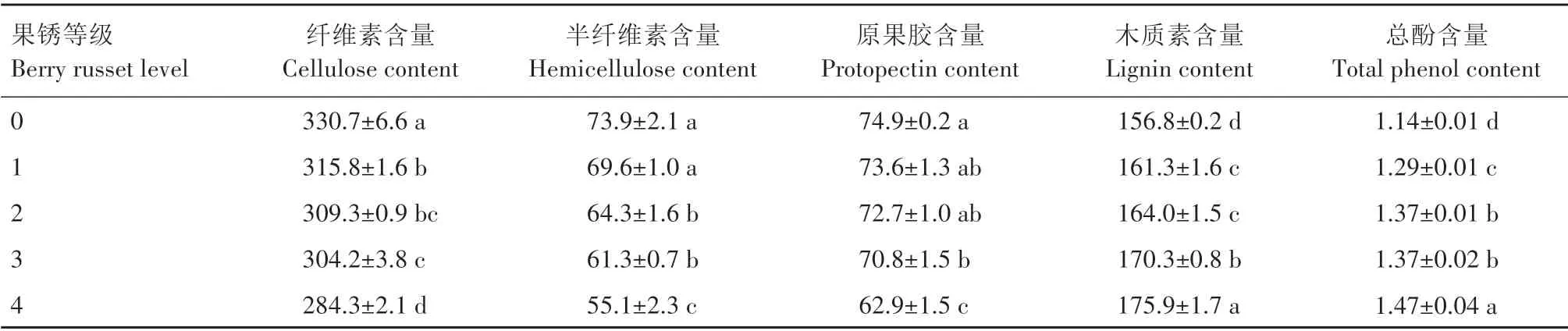
表1 不同果锈等级的阳光玫瑰葡萄果皮细胞壁及果锈相关物质含量的变化Tab.1 Changes in related substances of cell wall and berry russet of different berry russet levels in Shine Muscat grape peel mg/g
没有发生果锈的果皮木质素含量为156.8 mg/g,其显著低于所有发生果锈的果皮木质素含量;果锈等级1 和2 的果皮木质素含量分别为161.3 mg/g 和164.0 mg/g,两者均显著低于果锈等级3 和4 的果皮木质素含量;果锈等级3 和4 的果皮木质素含量分别为170.3 mg/g 和175.9 mg/g,两者之间差异显著。果锈等级4 的果皮总酚含量为1.47 mg/g,其显著高于其他所有等级的果皮总酚含量;果锈等级2 和3的果皮总酚含量均为1.37 mg/g,两者均显著高于果锈等级0和1的果皮总酚含量;果锈等级1的果皮总酚含量显著高于果锈等级0的果皮总酚含量。
2.5 不同果锈等级对阳光玫瑰葡萄果皮相关酶活性的影响
在与细胞壁降解相关酶中(表2),随着果锈等级的增加,阳光玫瑰葡萄果皮的纤维素酶、酸性磷酸酶活性呈现先升高后降低的变化趋势,碱性磷酸酶活性呈现先降低后升高的变化趋势,果胶酶活性整体上呈现逐渐升高的变化趋势。5个果锈等级的果皮纤维素酶活性之间差异均显著,其中,果锈等级3的果皮纤维素酶活性最高,为1 665.63µg/(min·g),果锈等级4 的果皮纤维素酶活性介于果锈等级0 与1 之间,果锈等级0 的果皮纤维素酶活性最低。果锈等级0的果皮果胶酶活性最低,为1.11 mg/(h·g),其显著低于其他所有等级的果皮果胶酶活性;果锈等级4的果皮果胶酶活性最高,为7.93 mg/(h·g),其显著高于其他所有等级的果皮果胶酶活性;果锈等级1 和3 的果胶酶活性之间差异不显著,且均显著低于果锈等级2 的果皮果胶酶活性。果锈等级2 的果皮酸性磷酸酶活性最高,为1.215µmol/(min·g),其与果锈等级3的果皮酸性磷酸酶活性之间差异不显著,但显著高于其他等级的果皮酸性磷酸酶活性;果锈等级1 与3 的果皮酸性磷酸酶活性之间差异不显著,且均显著高于果锈等级0 和4 的果皮酸性磷酸酶活性;果锈等级4 的果皮酸性磷酸酶活性最低。果锈等级2 的果皮碱性磷酸酶活性最低,为10.180µmol/(min·g),其显著低于果锈等级0、3和4的果皮碱性磷酸酶活性;果锈等级4 的果皮碱性磷酸酶活性最高,为12.696 µmol/(min·g),其显著高于其他所有等级的果皮碱性磷酸酶活性;果锈等级3与0的果皮碱性磷酸酶活性之间差异不显著。

表2 不同果锈等级的阳光玫瑰葡萄果皮相关酶活性的变化Tab.2 Changes in related enzyme activities of different berry russet levels in Shine Muscat grape peel
在果锈物质合成相关酶中,随着果锈等级的增加,阳光玫瑰葡萄果皮苯丙氨酸解氨酶活性呈现逐渐升高的变化趋势,过氧化物酶活性呈现先降低后升高的变化趋势,多酚氧化酶活性呈现逐渐降低的变化趋势。5 个果锈等级的果皮苯丙氨酸解氨酶活性之间差异均显著,其变化范围为8.205~11.354 U/g。果锈等级0、1 和2 的果皮过氧化物酶活性之间差异均不显著,三者均显著低于果锈等级3 和4 的果皮过氧化物酶活性;果锈等级3 和4 的果皮过氧化物酶活性分别为60.553 U/g和68.060 U/g,两者之间差异显著。5个果锈等级的果皮多酚氧化酶活性之间差异均显著,其变化范围为4.787~12.880 U/g。
3 结论与讨论
果锈是果实发育过程中的一种生理紊乱现象,在无核白鸡心、维多利亚、阳光玫瑰等黄绿色葡萄品种上经常发生[1,7],成为降低其商品性的主要因素之一。本研究调查发现,阳光玫瑰葡萄在生长前期没有果锈发生,进入软化期后才会有果锈发生,且随着果实的成熟,果锈发生率越来越高,这与之前的研究结果相似[1,11,19]。果锈的发生主要是次生代谢的结果[13⁃14],果实的次生代谢主要发生在果实发育后期[21],这也是造成果锈主要在近果实成熟期发生的主要原因。
袁高鹏[22]在金冠苹果果锈形成过程的观察发现,在果锈发生初期,果皮开始形成微裂隙,表皮细胞开始破裂,之后微裂隙进一步加深,形成V 形裂痕,果锈成分开始积累,最后V 形裂缝变得更长、更深,果锈成分大量沉积。阳光玫瑰葡萄果锈的发生过程与之不同,日本岛根农技研报[23]的报道表明,阳光玫瑰葡萄果锈的褐变部位位于表皮细胞组织内,以细胞间隙为中心,或是在亚表皮细胞层中被发现,且褐变部位正上方的表皮细胞层变薄,果皮表面呈现出凹凸不平的状态。SUEHIRO等[3]的研究也表明,阳光玫瑰葡萄果锈的褐变部位位于表皮细胞组织内,褐变物质位于果皮的细胞间隙。本研究结果与之相似,阳光玫瑰葡萄果锈首先发生在细胞间隙和细胞壁上,细胞结构凸显;随着发生程度的增加,果锈物质逐渐向细胞内部蔓延,褐化范围和细胞间隙增大,果皮表面呈现凹凸不平的状态;果锈严重时褐化物质渗透整个细胞,使细胞坏死。
植物细胞膜是调节细胞内外物质交流的屏障,同时对维持细胞的微环境和正常代谢起着重要作用。当植物受到逆境胁迫时,细胞膜遭到破坏,膜透性增大,从而使细胞内的电解质外渗,以致植物细胞浸提液的电导率增大[24⁃25]。本研究中,随着果锈发生程度的增加,阳光玫瑰葡萄果皮细胞膜的相对渗透率逐渐增加,且果锈等级3 和4 的果皮细胞膜的相对渗透率显著升高,说明果锈等级3 及以上等级的果皮细胞膜遭到严重破坏,细胞膜透性增大的程度与果锈发生程度密切相关。
纤维素是细胞壁的主要结构成分,对于维持细胞结构的稳定性具有重要作用;半纤维素是构成初生壁的主要成分,其结合在纤维素微纤维的表面,构成坚硬的细胞相互连接的网络[26]。果胶是构成细胞初生壁和中胶层的主要成分,对维持植物的结构和硬度起着重要作用[27]。张勃[28]在黄冠梨上的研究表明,果皮中果胶物质含量的增高及纤维素含量的降低可以提高果皮韧性和延展性,增大果皮破裂应力,从而抑制果实锈斑的发生。本研究中,随着果锈等级的增加,阳光玫瑰葡萄果皮纤维素、半纤维素和原果胶含量逐渐降低,且三者含量开始显著降低时的果锈等级分别为1、2 和3,说明阳光玫瑰葡萄果锈的发生首先是细胞壁的骨架—纤维素的快速降解,然后是纤维素微纤维间的网络—半纤维素的快速降解,最后是原果胶的快速降解,这种变化与纤维素酶和果胶酶的活性变化相一致。随着果锈等级的增加,阳光玫瑰葡萄果皮的纤维素酶活性呈现先升高后降低的变化趋势,果胶酶活性呈现逐渐升高的变化趋势,其中,果锈等级4的果皮纤维素酶活性的降低说明此时果皮纤维素的降解减慢,也间接说明果锈等级1~3 的果皮纤维素降解较快,从而造成果实一开始发生果锈便出现纤维素含量显著下降的变化;而果胶酶活性的持续升高说明原果胶是在果胶酶活性达到一定水平后才显著降解,从而造成果锈发生后期果皮的原果胶含量显著降低。
果锈是一层褐色的木栓化次生保护组织,木栓层的形成与木质素、酚类物质含量有关[8,19]。本研究结果表明,随着阳光玫瑰葡萄果锈等级的增加,果皮木质素、总酚含量逐渐增加,这与果皮组织结构观察的果皮褐化程度的增加相一致,也与冯娇[12]的研究结果相似,说明阳光玫瑰葡萄果锈的发生伴随着果皮中木质素和总酚含量的增加。木质素是苯丙烷类代谢途径的主要终产物,是木栓层形成、果实木质化的标志[29]。木质素是在多个酶促反应的作用下合成的,苯丙氨酸解氨酶、过氧化物酶、多酚氧化酶等均在木质素的合成过程中起着重要作用[13,15],其中,过氧化物酶和苯丙氨酸解氨酶是调控苹果、梨果实锈斑产生的关键酶[13,28,30],且WANG等[31]的研究表明,苹果苯丙氨酸解氨酶、过氧化物酶、多酚氧化酶活性的降低与表皮细胞木质素合成的减少密切相关,曹爱娟等[32]的研究表明,当苯丙氨酸解氨酶、过氧化物酶活性达到一个相对高的水平后,桃果皮木质素才开始快速积累。本研究中,随着果锈等级的增加,阳光玫瑰葡萄果皮的苯丙氨酸解氨酶活性逐渐升高,说明苯丙氨酸解氨酶活性与木质素的合成呈正相关,这与NAZARI 等[33]的研究结果一致,而与ASSIS 等[34]的研究结果相反,其研究中苯丙氨酸解氨酶活性的增加不能促进毛叶番荔枝果实木质素的合成。之前还有研究表明,枇杷皮的木质素合成与过氧化物酶活性密切相关[35],而本研究中,过氧化物酶活性随果锈等级的增加表现出先降低后升高的变化趋势,而多酚氧化酶活性表现出逐渐降低的变化趋势,这与HOU 等[16]的研究结果相似,其研究中阳光玫瑰葡萄果皮木质素的合成与过氧化物酶、多酚氧化酶活性的增加不一致,说明这2种酶不是调控阳光玫瑰葡萄果皮木质素合成的关键酶。
