肉牛种业科技创新发展现状与趋势分析
2022-11-22张天留葛菲朱波高会江李俊雅高雪
张天留 葛菲 朱波 高会江 李俊雅 高雪*
(1.中国农业科学院北京畜牧兽医研究所,北京 100193;2.河南农业大学动物科技学院,河南郑州 450046)
我国是牛肉生产和消费大国,据国家统计局和中国海关统计,2021 年我国肉牛存栏9817 万头,出栏4707 万头,牛肉产量约698 万吨,消费量达930 万吨,位居世界第三大生产国和第二大消费国。随着消费和生产缺口的拉开,2021 年我国牛肉进口量达到233万吨,几乎是美国进口量的2 倍,一跃成为世界牛肉第一大进口国。据国家肉牛牦牛产业技术体系研究表明,2030 年我国牛肉消费量将达到1200 万吨以上,如果没有明显的技术进步,我国牛肉生产远远不能满足需求增长。
良种对肉牛业发展的贡献率达40%,为牛肉产品稳产保供提供了关键支撑。自2011 年《全国肉牛遗传改良计划(2011—2025 年)》 实施以来,我国肉牛种业取得显著成效。但对标国际肉牛种业强国,我们仍存在许多方面的“卡脖子” 问题,一是自主品种市场占有率不高,“顶天立地” 的品种少;二是肉牛育种数据库基础性建设薄弱;三是精准高效育种技术的研发与应用仍处于起步阶段;四是地方种质资源挖掘与创新利用不足;五是肉牛联合育种机制仍需要探索。面对这些“卡脖子” 问题,科技创新是引领肉牛种业高质量发展的基础和关键。为深入贯彻落实习近平总书记关于打好种业翻身仗、推动科技自立自强等重要指示精神,该文立足“十四五” 面向“十五五”,准确把握肉牛种业领域的科技发展现状及趋势,探讨肉牛种业科技创新重大基础理论问题、核心关键技术研发和新品种培育能力提升等方面内容。同时,该文也为加强肉牛种业科技创新及产业战略部署,提升我国肉牛种业科技创新能力,肉牛种业高质量发展奠定基础。
1 肉牛种业基础研究现状
1.1 牛基因组学研究
2009 年,由美国、澳大利亚、新西兰等共计25个国家300 多位科学家组成的国际研究团队,发布了第一个以海福特牛为样品的牛基因组序列,正式拉开了牛基因组学研究的序幕。随后瘤牛、牦牛、水牛等不同牛属不同品种的参考基因组相继组装完成(见表1)。这些牛参考基因组的发布,为研究人员挖掘鉴定与牛生长发育、胴体、肉质和抗病等性状相关的重要功能基因提供了更加完善的牛基因组图谱。正如澳大利亚联邦生物信息研究带头人达尔利姆普勒博士所说:“希望牛基因组测序工作的完成能够有助于培育出肉质更好或产奶量更多的新品种牛”。
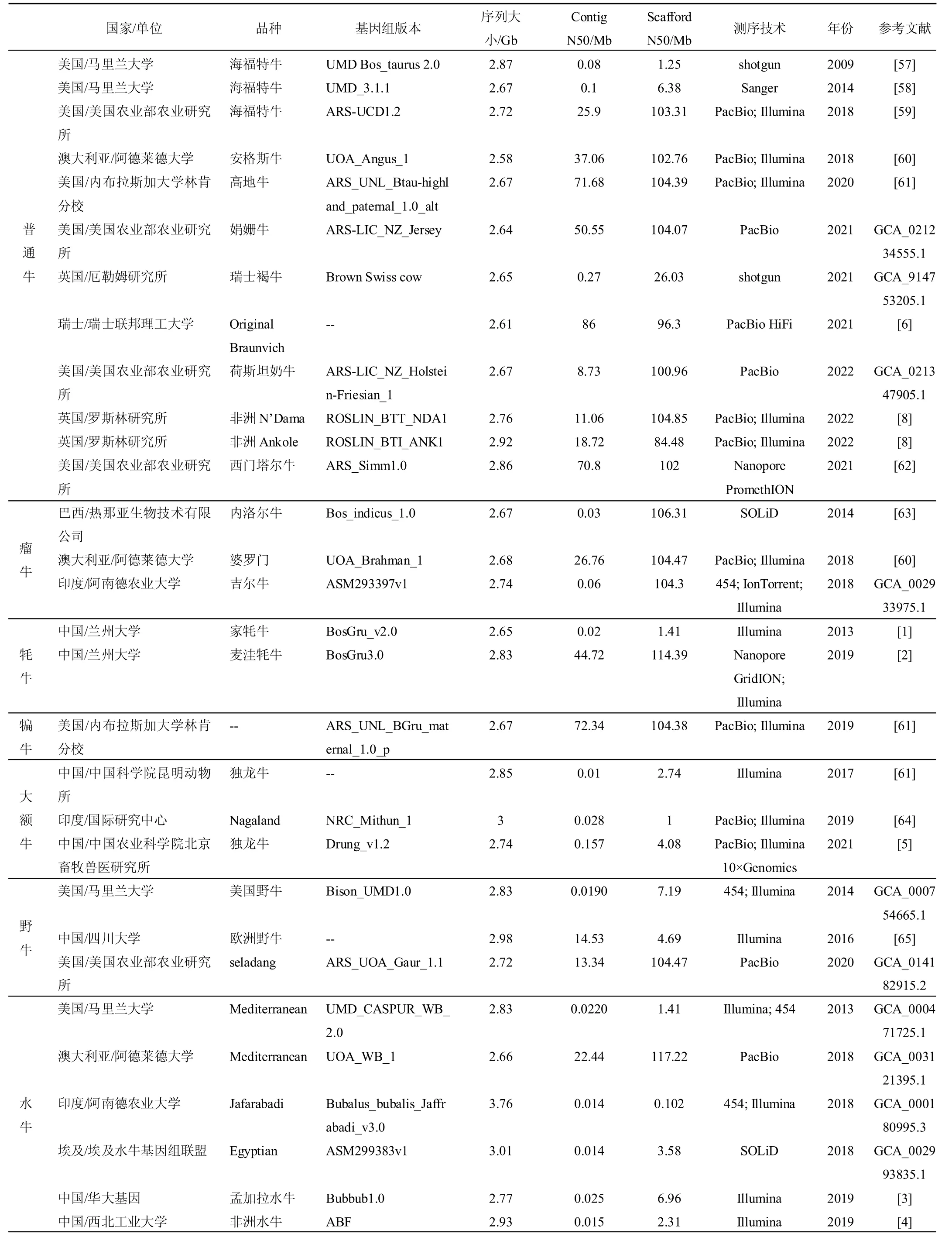
表1 国内外牛基因组组装研究进展
我国在牛基因组方面的研究也紧跟国际前沿,2012 年兰州大学组装了首个家牦牛的基因组[1],2021年又以麦洼牦牛为研究对象,组装了目前为止高质量的染色体水平家牦牛基因组,其Contig N50 长度达到44.72Mb,也是目前最高质量的反刍动物基因组。牦牛高质量基因组的完成,不仅有助于揭示牦牛产奶、产肉等重要经济性状以及其高原适应性的重要遗传机制,也将有助于进一步揭示人类所出现的各种高原不适症,促进对缺氧相关疾病的认识、预防和治疗[2]。此外,2016 年四川大学组装了欧洲野牛基因组[3];2019 年华大基因和西北工业大学分别组装孟加拉水牛、非洲水牛的基因组序列,为深入了解水牛这一物种的起源、驯化过程及其品种选育等奠定重要的遗传学基础[4]。2017 年中国科学院昆明动物所,2022 年中国农业科学院北京畜牧兽医研究所分别组装了独龙牛全基因组组装序列,并通过比较基因组学揭示了独龙牛的起源与系统地位问题,染色体融合的分子机制以及环境适应性的遗传基础,对我国地方牛种质资源的优异种质和基因精准鉴定具有重要意义[5]。
1.2 牛泛基因组学研究
随着三代测序组装的高质量参考基因组的陆续发布,以及大规模重测序和群体遗传学分析的广泛进行,研究人员发现来自单一个体的参考基因组远不能涵盖整个物种的所有遗传序列,因此泛基因组的概念应运而生。2021 年,瑞士苏黎世联邦理工学院发布了Original Braunvich 牛品种的基因组组装,并利用已发表的普通牛(海福特、安格斯、高地牛)、瘤牛(婆罗门牛)和牦牛基因组,构建了牛亚科物种第一个图结构泛基因组。与参考基因组(ARS-UCD1.2)相比,检测到超过7000 万个碱基缺失,包括参与免疫反应和调节的基因[6]。在2022 年5 月,瑞士苏黎世联邦理工学院联合美国农业部动物基因组学和改良实验室以瑞士褐牛的品种内杂交、内洛尔牛和瑞士褐牛的亚种间杂交、印度野牛和皮埃蒙特牛的种间杂交的后代中构建了单倍型分型基因组,结果表明在增加杂合度后,每一个分型基因组在连续性、完整性和准确性方面比目前的牛参考基因组都有了实质性的改善[7]。2022 年,英国爱丁堡大学构建了非洲N'Dama 和Ankole 两个品种公牛个体的高质量基因组(见表1),并结合294 头不同牛品种的重测数据构建了牛图结构泛基因组,发现了116.1Mb(4.2%)的变异在海福特牛基因组中缺失[8]。我国西北农林科技大学也利用已发表的12 个牛属物种基因组构建了牛泛基因组,发现了与免疫、代谢等功能相关的基因,全面挖掘了牛遗传多样性和本土适应机制[9](见表2)。

表2 国内外肉牛泛基因组构建研究进展
1.3 肉牛重要性状功能基因挖掘与遗传机制解析
随着高通量测序技术的飞速发展,肉牛重要经济性状的功能基因挖掘与遗传机制解析策略也从传统的候选基因法、QTL 定位等逐渐发展为各种组学技术的联合分析。近10 年来,国内外学者利用基因组学、转录组学、代谢组学和蛋白质组学等组学技术分析,挖掘鉴定了一批与肉牛生长、胴体、肉质、抗病及环境适应性等性状相关的功能基因和QTLs,为准确和全面揭示肉牛重要经济性状遗传调控机制及肉牛育种奠定了坚实基础。
1.3.1 具有育种价值新基因的挖掘与利用
1.3.1.1 无角性状
在现代化肉牛养殖中,牛角容易造成人员伤亡,而人为去角违背了动物福利且浪费人力物力,因此无角性状引发了高度关注。目前在安格斯、荷斯坦牛、西门塔尔牛等牛种鉴定出无角性状的候选突变PF 和PC。PF 突变是由1 号染色体80128bp(1909352-1989480bp)的重复以及3 个SNP 导致,主要存在黑白花奶牛。PC 突变是由1 号染色体上202bp(1705843-1706045bp)的重复,并替换了原有的10bp(1706051-1706060bp),主要存在于安格斯、西门塔尔牛中[10]。目前美国、德国已利用基因编辑技术将PC突变引入荷斯坦奶牛和荷尔斯坦-弗里西亚牛,培育无角新品种[11-12]。
我国在大通牦牛群体种鉴定出1 号染色体2048750~2196313bp 处长约147kb 的单倍型域内存在无角性状相关的单倍型,包含3 个蛋白编码序列C1H21orf62,GCFC1 和SYNJ1。研究表明,单倍型TGTTTGCCA 与大通牦牛无角性状相关,单倍型CCCGGATTG 与有角性状相关。中国农业科学院兰州畜牧与兽医研究所利用无角性状单倍型TGTTTGCCA选育,于2019 年培育成了“无角阿什旦牦牛”。
1.3.1.2 双肌性状
早在1807 年,英国农场主在短角牛群中发现双肌肉表型,到1997 年美国科学家才确定“牛双肌现象” 是由肌肉抑制素基因(myostatin,MSTN)发生了正常的基因自然突变所致。目前在短角牛、比利时蓝牛、皮尔蒙特牛、夏洛莱和德国黄牛等双肌群体中发现至少有9 个有义突变(S105C、F94L、nt821(del11)、Q204X、D182N、nt419(del7-ins10)、E226X、C313Y、E291X),这些突变均可引起不同程度的双肌现象。因此,各国育种者有意保留群体中的有义突变来选育双肌性状或者利用编辑技术引入双肌基因突变位点。阿斯图里亚纳牛(Asturiana de los Valles)原本是一个乳肉役多用途品种,通过加大双肌性状的选择,选育出MSTN 基因纯合个体,nt821(del11)突变频率在群体中高达93.6%。我国内蒙古大学以蒙古牛、鲁西牛和西门塔尔牛为对象,依据Myostatin 基因突变多态特点及分析自然突变牛的出生、生长与生产性状,选择基本不影响出生重及后期发育性能良好 的g.507del(6)、g.505del(115)、g.415C>A 和g.3942T>G 4 个位点作为基因编辑靶标,利用基因编辑、动物克隆、MOET 与常规繁殖技术成功培育出双肌鲁西牛、双肌蒙古牛和双肌西门塔尔牛等基因编辑肉牛新品系。
1.3.2 功能基因及分子标记研究与应用
1.3.2.1 生长与胴体性状
生长发育和胴体性状是肉牛生产中最重要的经济性状,可占胴体价格影响因素的60%~70%。目前在西门塔尔牛、利木赞、海福特、夏洛莱、安格斯、瑞士褐牛和荷斯坦牛群体中进行Meta-GWAS 和eQTL分析,定位到LCORL/NCAPG 基因SNP 间的LD 很高,并发现LCORL 的一个错义突变与牛的体型显著相关,该基因与人、大鼠和小鼠身高变化也有关[13]。此外,在欧洲牛品种的基因组研究结果中,对37 个牛品种(17 个肉牛品种,14 个奶牛品种,6 个兼用品种)的核心选择信号区域(CSS)进行了报道,鉴定到291 个CSS 与体尺性状相关候选基因[14]。
我国在中国肉用西门塔尔牛群体中基于一步法GWAS、单倍型、拷贝数变异等策略鉴定到与体高、十字部高等体尺性状,初生重、日增重等生长发育性状相关的候选基因,如SOX2、LCORL。以胴体重、屠宰率、净肉率、上脑重、和尚头、金钱腱和骨重等胴体性状为研究对象,鉴定到LCORL、PLAG1、RYR、EML6、HOMER1 和SAMD12 候选基因[15-19]。在中国和牛群体中基于全基因组水平分别检测到18个、5 个和1 个与十字部高、身高和体长相关的SNP,这些SNP 在11 个候选基因内或附近,其中6 个候选基因(PENK、XKR4、IMPAD1、PLAG1、CCND2和SNTG1)已经在多个群体被报道过[20-21]。此外,在地方黄牛的研究中发现,POMC 基因的变异位点与关岭黄牛的体斜长具有关联性[22],还与南阳牛体重相关[23]。SH2B1 基因的多态性显著影响秦川牛的体高、体长、坐骨端宽和胸围[24]。FGF21 基因第297 位和第940 位的多态性与南阳牛18 月龄体重显著相关[25]。
1.3.2.2 饲喂效率性状
在肉牛生产中,采食量和饲料利用效率是极其重要的经济性状,常用饲料转化率(FCR)、料重比(F/G)和剩余采食量(RFI)来衡量饲料利用率。在遗传基础解析上,在巴西内洛尔肉用牛群体中对平均日增重、干物质采食量、FCR、RFI 等性状进行了遗传参数估计和相关性测定,结果表明,RFI 与平均日增重、干物质采食量和FCR 的遗传相关分别为-0.19、0.61 和0.50,表型相关分别为0.03、0.65 和0.35[26]。在肉牛的RFI 及脂肪酸等性状的相关性研究中得到RFI 与n-6/n-3、18:2n-6 等不饱和脂肪酸间的遗传相关系数绝对值在0.4 以上(-0.430.52),说明低RFI 的选育同时也可以改善脂肪酸组成,有利于人体健康[27]。在功能基因挖掘方面,在安格斯、内洛尔等群体中采用GWAS 策略鉴定到与RFI 相关的分子标记,主要与OAS2、STC2、SHOX、XKR4、SGMS1、DCP1B和SUPT20H 候选基因相关[28]。此外,对高低RFI 组的内洛尔牛的骨骼肌和肝脏中表达调控研究(miRNA-mRNA),结果显示共鉴定到310 个牛的miRNA,其中miR-7、miR-199a-3p 和miR-486 等10 个miRNA可能与RFI 性状变异相关。同时,针对差异表达miRNA 的靶基因和miRNA-mRNA 相关模块进行功能富集分析发现,与RFI 性状相关的信号通路涉及胰岛素、脂质、免疫系统、氧化应激和肌肉发育[29]。
1.3.2.3 肉质性状
肉质性状作为复杂的经济性状,其度量指标众多,但它们之间又有相互关联。国际上已在普通牛(日本和牛、韩牛、安格斯、海福特、莫罗黑牛和短角牛),地方热带牛(贝尔蒙特红牛和圣格鲁迪牛)和瘤牛(婆罗门牛)等多个肉牛群体脂肪酸组成的遗传结构分析中发现了多个SNPs 与脂肪酸组分存在显著相关,研究证明,FASN 和SCD 基因对脂肪酸存在较大的影响[30-32]。同时,也鉴定到与肉嫩度、眼肌面积、背膘厚度和大理石纹相关的QTL 区间,基因注释得到与ASAP1、CAPN1、CAPN5[33-34]。此外,在蛋白质和代谢水平中,应用iTRAQ 和HPLC-ESI-MS 技术对不同部位的分割肉块中肌肉颜色和氧化稳定性相关的营养和代谢物质鉴定,确定了酰基肉碱、游离氨基酸、核苷酸、核苷和葡糖醛酸酯等候选物质与肉品质密切相关[35-36]。
在国内研究中,已在华西牛、中国和牛杂交群体中鉴定到与脂肪酸组分相关的候选基因,如ELOVL5、FASN、CASP2、TG[37-38]。同时,在华西牛群体的脂肪颜色、肉色、大理石花纹、眼肌面积和剪切力等5 个肉质形状进行关联分析,找到TMEM236、SORL1、TRDN、S100A10、AP2S1 等一系列候选基因[39]。在地方黄牛中,在郏县红牛、鲁西牛和秦川牛群体中均发现GPR41 基因作为脂肪酸感受器可以感知来自消化腔的短链脂肪酸,能够促进瘦素的分泌、调节糖脂和能量代谢、控制体重等功能[40]。此外,采用转录组和蛋白质策略得到了中国晋江黄牛早期背最长肌和腰大肌分子差异,鉴定到与肉质性状相关基因(MTND2、ND4L、COII、SLC16A7、HOXC6),揭示了酶活性(乳酸脱氢酶、苹果酸脱氢酶和琥珀酸脱氢酶)、丙酮酸含量等代谢物数量发生的变化[41-42]。
1.3.2.4 繁殖性状
繁殖性状作为重要的生产性状,与动物生产成本和经济效益密切相关。Oliver 等发现位于BTA19 的ASIC2 和SPACA3 基因参与氧化应激,影响妊娠[43]。Stegemiller 等通过GWAS 确定了卵泡计数(AFC)位点在2、3 和23 号染色体,生殖道评分(RTS)位点在2、8、10 和11 号染色体,STC1 基因与排卵数减少显著相关,跨膜蛋白260 和胞嘧啶/尿嘧啶单磷酸激酶2 与繁殖性能相关[44]。有趣的是,与嫩度相关的基因会影响牛繁殖性能,CAPN1 等位基因与肉牛产后发情间隔的增加有关,CAST 多态性与牛的生育能力和繁殖寿命有关,DGAT1 多态性也与牛繁殖性状相关,表明使用CAPN1 等标记增加嫩度,可能会延迟母牛地再繁殖[45]。
1.3.2.5 环境适应性
我国有丰富的地方牛品种遗传资源,不同地区牛品种经历了长期的选择和培育后能够生活在差异的环境条件下,其机体已经形成了独特的适应性进化体系。如在寒冷气候适应性研究方面,在延边牛群体中鉴定出3 个候选基因(CORT、FGF5 和CD36),推测出错意突变(c.638A>G)是延边牛耐冷性的候选基因[46]。在大通牦牛和蒙古牛群体中,通过SNP 位点的多态性比例差异分析,鉴定到与热休克蛋白、氧气运输、线粒体DNA 维持、代谢活动、采食量、胴体结构和生殖相关基因[47]。同时,通过比较选取我国南、北方代表型的蒙古牛和闽南牛,鉴定出大量差异性变异,获得1096 和529 个潜在选择基因分别与环境适应性、饲料效率和肉/奶产量等性状相关[48]。此外,高原适应性上,在牦牛[1]、藏牛[49-50]、大额牛[5]群体中逐步挖掘到一系列环境适应性关键候选基因,如心血管系统方面:兰尼碱受体2(RYR2),该基因编码的蛋白是钙通道上Ca2+运输的重要组成因子,当处于缺氧环境中,通常会导致细胞内Ca2+的释放,引发心肌兴奋-收缩耦联过程,刺激细胞收缩,应对低氧反应[51]。低氧应答方面,内皮PAS 蛋白1(EPAS1)、缺氧诱导因子α(HIF-1α)、促红细胞生成素(EPO)、一氧化氮合酶1(NOS1)、血红蛋白β 亚基(HBB)促进血管生成,维持血液中含氧气和二氧化碳浓度变化,参与应急缺氧调节等生物过程[52-56]。
2 肉牛种业核心关键技术研究现状
2.1 常规遗传评估技术
近50 年以来,世界各国肉牛育种工作者经过不断研究与探索,使得肉牛遗传评定方法不断改进和完善。最早是1975 年美国康奈尔大学Henderson 教授使用的公畜模型BLUP 方法。2001 年,Interbull 官方公布指南中指出,对于生产性状而言,动物模型优于公畜模型,多性状模型优于单性状模型;针对阈性状而言,公畜模型和公畜-外祖父模型估计结果更为准确。育种值估计软件常用的有美国1993 年开发的MTDFREML 和1997 年开发的BLUPF90,英国1995 年开发的ASREML 和丹麦2013 年开发的DMU 等。
由于各国肉牛主要品种有一定差异,而不同品种在生产性能方面各有差异,如法国、美国和澳大利亚存栏较多的中大型肉牛品种夏洛莱、利木赞和西门塔尔牛在生长速度上更加突出,美国和澳大利亚存栏较多的中小型肉牛品种安格斯及日本的和牛在肉质性状上更优秀,美国和印度存栏较多的瘤牛品种如婆罗门牛和巴西的内洛尔牛在耐热能力和屠宰率上优势更明显。因此,各国在进行遗传评估时,制定综合选择指数在所选择的性状和权重方面也有一定差异。以西门塔尔牛为例,美国和加拿大评估了生长性状、胴体和肉质性状及繁殖性状共计15 个性状,依据不同的育种目标制定了API 和TI 两个综合选择指数(https://simmental.org/site/index.php);英国评估了生长性状、胴体和肉质性状及繁殖性状共计15 个性状,制定了2个综合选择指数(http://www.britishsimmental.co.uk/);澳大利亚同样评估了生长性状、胴体和肉质性状及繁殖性状共计18 个性状,制定了4 个综合选择指数(https://simmental.com.au/)。
我国肉牛遗传评估工作从2010 年才开始,起步较晚。根据国内肉用种公牛育种数据的实际情况,选择体型外貌评分、6~12 月龄日增重、13~18 月龄日增重和19~24 月龄日增重4 个性状,制定了中国肉牛选择指数CBI 对肉用种牛进行评估。为了对接国外遗传评估体系,2020 年7 月修订了CBI 指数,选择体型外貌评分、出生重、6 月龄重和18 月龄重4 个性状。乳肉兼用牛的遗传评估则使用总性能指数TPI 进行评估。我国目前肉牛生产体系和性能测定体系使得制定的综合选择指数暂时只能利用生长发育性状和母亲产奶量性状。而欧洲、北美和澳洲等肉牛育种发达国家的产业体系、性能测定体系和遗传评估体系较完善,考虑性状较多。
2.2 基因组育种评估技术
进入21 世纪以来,基于基因组高密度标记信息的基因组选择技术(Genomic Selection,简称GS)成为动物育种领域的研究热点。利用该技术,可实现青年公牛早期准确选择,而不必通过后裔测定,从而大幅度缩短世代间隔,加快群体遗传进展,并显著降低育种成本。
2014 年,美国安格斯协会率先开展安格斯青年公牛基因组遗传评估。英国、爱尔兰、德国、法国、丹麦等欧美主要发达国家陆续将基因组选择技术全面应用于不同品种的基因组育种值估计中。以北美为代表的肉牛基因组遗传评估体系开展的时间最早,也是目前最成熟且应用最新计算方法的体系,主要性状的基因组遗传评估可靠性达到70%~80%。以西门塔尔牛品种为例,其制定的API 和TI 指数也是多个国家肉牛育种的主要参考指数,同时美国开发的商业化BLOT 遗传评估系统,该评估系统能综合各品种信息,同时整合系谱和基因组信息进行多品种SSGBLUP 评定,也是目前主流的基因组遗传评估系统之一。欧洲肉牛基因组遗传评估以Interbeef 评估中心为主,大部分欧洲国家加入了该组织,该中心数据库有超200 万头牛基因型数据,以单品种基因组遗传评估为主,目前还未报道相关基因组选择综合选择指数。澳洲肉牛基因组遗传评估系统以BREEDPLAN 为主,主要是澳大利亚和新西兰的各品种肉牛联合会进行评定,其基因型测定个体要少于北美和欧洲。拉丁美洲的基因组遗传评估主要是巴西的内洛尔牛,其他品种无相关报道。
我国基因组选择技术研究从2008 年开始,中国农业科学院北京畜牧兽医研究所自主研发建立了肉用牛基因组选择技术平台,构建了肉用西门塔尔牛与和牛基因组选择参考群体,目前西门塔尔牛参考群体3858 头,和牛参考群体462 头。2019 年制定了基因组中国肉牛基因组选择指数GCBI(China Genomic Beef Index,GCBI),2020 年我国首次在中国肉用及乳肉兼用种公牛遗传评估概要中发布了366 头西门塔尔种公牛的基因组遗传评估结果。从计算方法上,我国基因组评估方法采用的是贝叶斯方法,而国外多采用SSGBLUP 方法,主要由于国外有大量的系谱和表型数据,针对我国表型数据和系谱数据较少的情况下,贝叶斯方法在我国应用更有优势。从参考群测定的表型性状看,我国参考群测定了生长发育、屠宰、胴体、肉质和繁殖性状共计87 个性状,是目前基因组选择测定表型性状最多的国家。
2.3 智能表型测定与收集技术
准确、高效、智能的性状测定技术是肉牛育种的前提和关键基础性工作,育种与科学研究均依赖于表型数据的准确获取。近年来,随着物联网、视频与人工智能技术的结合,肉牛育种性状测定与采集技术手段不断更新与拓展,尤其以信息采集为基础的信息化技术得到发展和应用,如牛的体温、反刍次数、瘤胃pH、运动等智能数据采集技术。美国、英国、澳大利亚等主要发达国家已陆续将信息技术、红外感应、影像捕获、物联网等技术运用到肉牛个体识别和性能测定中,并持续研发性能测定的智能化装备和技术,诞生了一批世界领先的智能化性能测定设备制造企业。而国内大部分肉牛生产性能测定还比较粗放,效率和精确度低下;有少数企业和科研单位通过引进消化吸收和自主创新,开发出肉牛自动称重、CT 扫描等设备,但硬件及配套软件仍需完善和优化升级。
2.4 肉牛繁殖技术
繁殖技术可有效提升优秀肉用种牛遗传物质的推广效率,加速肉牛群体的遗传改良。目前人工授精、母牛发情高效精准鉴定、定时输精、胚胎移植等技术已在国内外规模化肉牛养殖场得到普遍应用。近20年,肉牛冷冻精液生产工艺和人工授精技术无实质性突破,配种受胎率有待提高。另外,随着肉牛活体采卵体外受精(OPU-IVF)技术日益成熟,有望进一步提升肉牛遗传改良效率。在美国、加拿大等肉牛发达国家,OPU-IVF-ET 技术已经应用到高产肉牛核心群快速扩繁生产中,显著提高了群体育种速度和肉牛生产性能。而我国肉牛胚胎移植技术、OPU-IVF 生产技术研发力度不够,总体应用效率低,推广使用规模和范围小,难以满足肉牛业发展对良种牛的实际需求。
2.5 基因修饰编辑育种技术
随着生命科学领域的不断发展进步,基因编辑技术(Gene Editing,GE)逐渐兴起,为肉牛生物育种提供了新思路。与传统转基因等技术不同,GE 可把目标基因引入基因组中的任何特定位置,或在任何位置进行编辑,产生所需的目标性状动物,可大幅提高修改基因组的效率和精确度。目前全球范围内,基因编辑技术已经在水稻、玉米、大豆、小麦和番茄等农作物以及猪、牛、羊等农业动物中广泛应用;糯玉米、高油酸大豆、抗褐变马铃薯、高GABA 番茄、抵抗褐变的蘑菇等基因编辑产品陆续在美国、日本等国家上市推广。
20 世纪90 年代左右,国外牛的转基因育种主要集中在乳腺生物反应器、改善乳品质、抗病能力等方面,而近10 年则在无角牛的基因编辑上取得重大进展。2013 年,美国明尼苏达州大学Tan Wenfang 等通过TALENs 技术编辑无角安格斯牛1 号染色体上Pc突变(即BAT1 1705843~1706045bp 处10bp 缺失的212bp 重复序列),得到编辑的荷斯坦牛胎儿成纤维阳性细胞,通过体细胞核移植技术,成功获得2 头成活的无角荷斯坦奶牛(Spotigy 和Buri),实现了将肉牛Pc 突变成功引入荷斯坦奶牛。2022 年德国科学家Felix Schuster 等利用CRISPR/Cas12a 编辑技术将无角安格斯牛Pc 突变转入荷尔斯坦-弗里西亚公牛,获得2头编辑牛,其中1 头90 日龄流产,经表型和组织学分析具有无角基因型;另外1 头出生后死亡,但具有无角表型。
我国在牛转基因和基因编辑育种研究领域的研究处于国际领先水平。中国农业大学、西北农林科技大学和内蒙古大学分别在转基因牛生物反应器、转基因与基因编辑抗病牛、转基因与基因编辑高产优质肉牛育种研究方面,取得突破性进展(见表3):乳清蛋白、乳铁蛋白转基因牛;抗乳腺炎、抗结核病与抗口蹄疫转基因牛;fat-1 转基因牛、Myostatin 基因编辑蒙古牛、鲁西牛及西门塔尔牛等均进入生产性试验阶段,获得国家生产性试验证书或生产证书,目前正在培育转基因或基因编辑新品系。

表3 2012—2022 年我国肉牛转基因和基因编辑技术研究进展
3 肉牛种业科技创新发展趋势
3.1 基因组与泛基因组学研究仍是未来基础性研究热点
尽管目前已经报道了普通牛、瘤牛、牦牛、水牛、野牛等不同牛属不同品种不同版本的30 个参考基因组,但全球有近1000 多个牛品种,分布范围广、遗传背景复杂、基因组差异大,已发布的基因组并不能饱含所有牛种的基因变异,因此开展肉牛基因组计划,构建高精度、高敏感性的基因组功能注释图谱,全面解析肉牛重要性状形成的遗传机制,提高肉牛基因组育种选择的准确度和敏感性。另外,相对于传统的线性参考基因组,泛基因组在基因组de novo 的基础上,基于图论的组装方法,更有助于挖掘品种特异性结构变异和特征序列。因此选择代表性的牛品种,从头组装各品种基因组,构建全面覆盖牛品种特征,序列的图结构泛基因组,将为肉牛高精度分子育种提供坚实的基因组数据支撑。
3.2 多组学联合分析挖掘重要育种目标性状功能基因
随着不同肉牛品种高质量、高精度基因组序列不断发布,肉牛泛基因组的构建以及转录组、代谢组、蛋白组等组学技术快速发展,利用多组学联合解析重要经济性状遗传机制已成为未来必然趋势。基于此,开展肉牛重要性状遗传机制与生理学基础研究,整合利用多组学技术以及生物信息学、系统生物学策略,挖掘与鉴定影响生长性状、胴体、肉质、抗病、繁殖和长寿等重要性状的功能基因和分子标记,解析分子遗传机制,鉴定具有育种价值的优异基因,为开展分子育种提供必要的基因信息。
3.3 基因组选择技术研究是未来肉牛育种技术主攻方向
随着测序技术的不断发展以及测序成本的下降,获得基因组、转录组、代谢组、蛋白组等海量数据信息,如何整合不同来源的数据到基因组选择技术中是未来肉牛育种技术的主攻方向。优化基因组评估模型和方法,利用深度学习算法整合不同来源的先验数据,包括GWAS 显著SNP 位点、重要通路等多组学信息,并结合高效计算的线性混合模型(GBLUP、SS-GBLUP 等)、预测准确的贝叶斯方法(BayesR、BayesCπ 等),拟合优化基因组评估模型和方法。整合基因功能注释、转录组、调控元件、表观遗传等生物信息的基因组选择技术,基于基因组信息的最优化选配技术,育种目标经济学分析及优化育种方案研究是未来我国乃至世界基因组育种新技术研究的制高点。
3.4 生物育种与数字化育种
3.4.1 生物育种
基因组编辑技术可以从其他品种或物种引入已知的理想等位基因或本品种内创造新的优势等位基因,而不会产生与传统基因渗入相关的连锁阻力,为农业动物优异基因资源的高效聚合及创制提供了新策略。Bastiaansen 等[74]利用数学模型分析,当效率为100%时,基因组编辑可使所需等位基因的固定速度提高4倍。因而,创新基因编辑技术,提高编辑效率,仍是今后的技术攻关难点,重点开发精准高效的基因编辑系统,确保对目标基因具有特异性且不会攻击其他基因,建立并优化精准多位点基因编辑体系。
3.4.2 数字化育种
数字技术引入农业领域代表着第四次农业革命。数字畜牧业技术有自动育种数据收集系统、自动发情识别系统、动物摄食及呼吸心率微控制器、疾病预警系统、虚拟围栏控制系统、面部识别系统、生物传感器,以及大数据分析和机器学习算法等。肉牛业的数字化将实现从二维的物理-社会系统到三维的网络-物理-社会系统的转变,改变未来肉牛种业的发展。
3.5 新兴生物技术的探索与应用
3.5.1 干细胞育种技术
干细胞育种技术是根据育种规划,利用基因组选择技术、干细胞建系与定向分化技术、体外受精与胚胎生产技术,在实验室通过体外实现家畜多世代选种与选配的育种新技术。与传统育种技术体系相比,该方法用胚胎替代个体,完成胚胎育种值估计,育种周期大幅度缩短,有望能革新全球家畜种业格局,实现家畜育种跨越式、颠覆性发展。
3.5.2 胚胎基因编辑育种
家畜胚胎基因编辑育种取得突破,通过直接编辑决定遗传性状的胚胎基因,即可精确改良家畜遗传性状,获得一批传统育种较难培育、肉用性能和抗病力显著提高的牛,彻底颠覆了通过表型性状进行选育的传统育种技术路线,成为肉牛种业革命新引擎。
3.5.3 新型配子与胚胎工程技术
未来,足量配子和胚胎供给是家畜遗传改良与生物育种两个领域的焦点问题。目前,小鼠和人的多能干细胞向配子方向的定向分化已经取得初步成果,但距离健康活体动物的生产尚存在很大差距。而家畜配子的诱导体系尚未取得突破性进展,目前仍没有通过牛多能干细胞诱导获得真正原始生殖细胞(PGCs)的报道。与以往的iPS 干细胞不同,“诱导全能干细胞(iTS 干细胞)” 同时具有发育为胚内和胚外组织的潜能。诱导处理iTS 干细胞向自然受精囊胚的牛“类囊胚样(blastoid-like)” 结构,可逐渐培育出具有繁育能力的人工诱导胚胎。将基因编辑技术和“人造” 配子技术相结合,直接在配子阶段编辑决定性状的基因,可实现经济、准确、高效的生物育种目标。
