黄芪甲苷通过JAK/STAT3信号通路抑制肺癌细胞增殖和迁移的作用机制研究
2022-11-21张志宏王春梅孙靖辉张成义陈建光
张志宏,王春梅,李 贺,孙靖辉,张成义,陈建光
(北华大学药学院,吉林 吉林 132013)
据2022年美国癌症学会最新数据[1]显示:肺癌是目前发病率最高的恶性肿瘤,同时是男性死亡率第一位(22%)、女性死亡率第二位的恶性肿瘤(17%)(仅次于乳腺癌死亡率).肺癌患者临床症状不明显,发现时常常已是中晚期.对于失去手术机会的患者,靶向治疗是延长生存期并改善生活质量的重要治疗方式.研究[2]表明:肺肿瘤局部增殖和远处迁移的生物学特性是导致肺癌发病率和死亡率居高不下的主要原因.因此,寻找有效影响肺癌增殖和迁移分子靶点的天然小分子化合物是当前肺癌研究中亟须解决的问题之一.
Janus激酶/信号转导和转录激活因子3(JAK/STAT3)信号通路与许多癌症的发生和进展有关[3].研究[4]表明:肿瘤细胞中JAK-STAT信号途径异常活跃可使活化的STAT3与受体分离形成二聚体,从细胞质转运到细胞核并利用其DNA结合域与核内特定DNA序列结合,行使基因转录调控的功能,促进与肿瘤细胞增殖和迁移的基因表达,进而促进癌细胞增殖和迁移.除了直接影响肿瘤细胞外,JAK/STAT3激活后,可通过上调EMT相关转录因子而增强其转移[5].总之,靶向JAK/STAT3信号通路已被证明对治疗多种癌症有益,因此,在抑制肿瘤生长和恢复抗肿瘤免疫方面具有相当大的应用前景.
黄芪是一种在临床治疗中常用的中药,现代药理学表明其具有提高免疫、脏器保护、抗肿瘤等疗效.黄芪甲苷为黄芪制剂质量鉴定的指标成分,在中国药典中被作为黄芪质量检测的标志物.黄芪甲苷是中药黄芪的主要活性成分,具有多种药理作用,如抗炎、抗氧化和免疫调节作用[6].此外,黄芪甲苷已表明具有抗肿瘤的药理活性[7].然而,黄芪甲苷是否会通过JAK/STAT3途径抑制肺癌尚不明确.因此,在本研究旨在探究黄芪甲苷是否以JAK/STAT3途径干预肺癌细胞增殖和迁移,以期为开发肺癌靶向治疗策略提供依据.
1 材料与方法
1.1 细胞与试剂材料
人肺癌细胞A549细胞(北华大学五味子开发及产业化工程研究中心);胎牛血清(FBS,天津市灏洋生物制品科技有限责任公司);胰蛋白酶(含EDTA)、DMEM培养基(赛默飞科技有限公司);p-JAK1、p-STAT3、GAPDH、HRP链接的二抗(兔抗、鼠抗,武汉爱博泰克生物科技有限公司);BCA蛋白浓度测定试剂盒(北京鼎国昌盛生物技术有限公司);结晶紫(默克公司,德国);荧光素酶检测试剂盒(普洛麦格公司,美国);黄芪甲苷(AS-IV,C41H68O14,纯度≥98%,成都普菲德生物技术有限公司);细胞实验时,将黄芪甲苷溶于二甲基亚砜(DMSO)中,备用.
1.2 实验方法
1.2.1 细胞培养
A549培养于DMEM培养液中(含10%胎牛血清、100 U/mL青霉素-链霉素),培养条件为37 ℃、5% CO2、湿度95%,细胞生长对数期进行传代、备用.
1.2.2 MTT实验
MTT实验用于检测细胞活性,A549细胞以1×104个/孔接种于96孔板中,培养6 h.AS-IV以0、5、10、20、30、40 μmol/L 6个浓度给药,继续培养24 h.加入MTT溶液(5 mg/mL,10 μL),培养4 h,弃去上清液;溶解DMSO,测定490 nm处各孔内吸光度值,将每个孔的吸光度与AS-IV 0 μmol/L处理组进行比较.
1.2.3 荧光素酶报告基因实验
应用荧光素酶报告基因实验检测STAT3的转录活性,脂质体2000将pGL3-STAT3和pRL-CMV荧光素酶质粒共转染入A549细胞.孵育24 h,细胞(1×105个细胞/孔)被接种在96孔板中,并用黄芪甲苷孵育12 h.通过荧光素酶检测试剂盒测量荧光素酶活性,进而判断STAT3的转录活性.
1.2.4 蛋白免疫印迹实验
应用蛋白免疫印迹实验检测细胞p-JAK2、p-STAT3蛋白表达水平,A549细胞以1×105个/孔接种于6孔板中,继续培养细胞12 h后,AS-IV以0、5、10、20 μmol/L 4个浓度给药,继续培养12 h.应用细胞裂解液进行蛋白提取,按照BCA试剂盒方法进行蛋白定量,以20 μg/孔蛋白量上样,进行凝胶电泳分离、转膜、孵育一抗(p-JAK1、p-STAT3、GAPDH抗体),4 ℃孵育过夜,次日加入二抗室温孵育1 h,洗膜,ECL显影液进行显色处理.
1.2.5 集落形成实验
应用集落形成实验检测各组细胞克隆形成能力,将细胞接种于6孔板中,用AS-IV对细胞进行处理.10 d后应用甲醇固定,0.1%结晶紫染色液进行染色,冲洗、干燥后拍照.
1.2.6 划痕实验检测
应用划痕实验检测细胞迁移能力,将细胞接种于24孔板中,待细胞长至90%密度时用200 μL Tip制造同等伤口,并且进行分组及给药处理.48 h后,再次观察、拍照记录伤口大小,计算细胞相对迁移率.
1.3 统计学分析

2 结 果
2.1 AS-IV对A549细胞活性的影响
AS-IV为羊毛脂醇型四环三萜皂苷,其化学结构见图1 a.我们通过MTT测定,评估AS-IV对A549细胞毒性的影响.结果表明,AS-IV在浓度低20 μmol/L时没有明显细胞毒性.因此,接下来实验选择浓度低于20 μmol/L的AS-IV进行进一步实验.见图1 b.
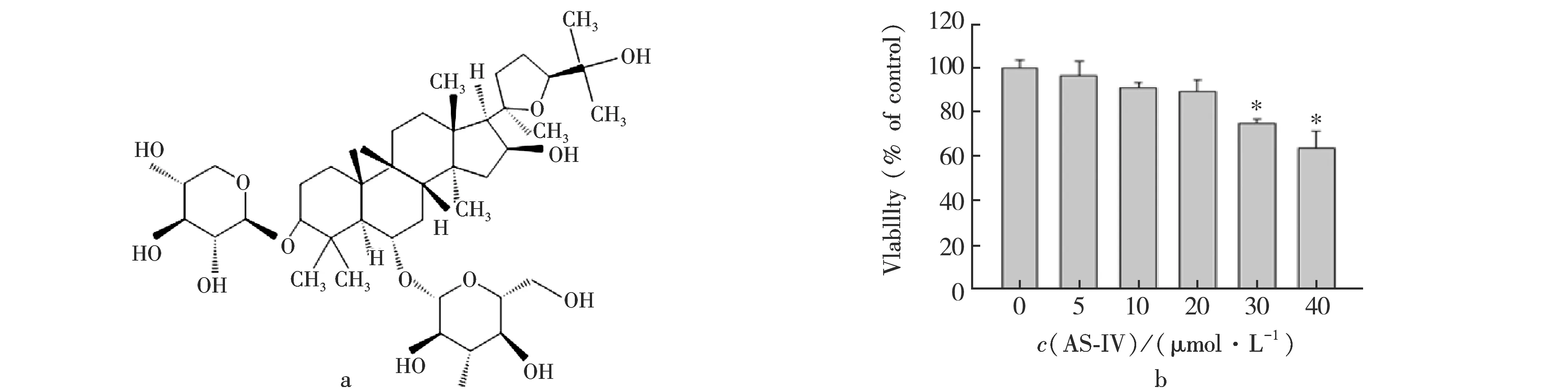
*.与对照组比较P<0.05.图1黄芪甲苷结构式(a)与黄芪甲苷对A549细胞毒性的影响(b)Fig.1Effect of chemical structure of AS-IV (a) and Astragaloside IV on cytotoxicity of A549 (b)
2.2 AS-IV对A549细胞JAK/STAT3信号途径的影响
JAK/STAT3通路具有介导肿瘤形成和进展的关键特性,如增殖、转移和细胞存活等,因此,在本研究中,我们试图确定AS-IV是否对JAK/STAT3信号通路有影响.首先,我们进行了STAT3基因分析,探讨AS-IV是否调节STAT3转录活性.结果表明:AS-IV以剂量依赖性方式显著抑制STAT3转录活性(见图2 a).由于STAT3蛋白是潜在的细胞质转录因子,其上游JAK1磷酸化可促进STAT3磷酸化,促使STAT3激活,因此,我们研究了AS-IV对p-JAK1和p-STAT3的影响,结果发现,在A549细胞,AS-IV以剂量依赖性方式有效抑制p-JAK1和p-STAT3蛋白表达(见图2 b).以上结果表明,AS-IV可抑制A549细胞JAK/STAT3信号途径.

与对照组比较,*.P<0.05,**.P<0.01,***.P<0.001.图2黄芪甲苷抑制STAT3转录活性(a)及JAK/STAT3信号途径(b)Fig.2Astragaloside IV inhibits STAT3 transcriptional activity (a) and JAK/STAT3 signaling pathway (b)
2.3 AS-IV对A549细胞增殖和迁移的影响
异常的STAT3激活是通过多种机制促进肿瘤细胞增殖和迁移,为此,我们通过细胞集落实验和划痕实验探究五味子乙素对A549细胞长期增殖和迁移的影响.结果显示:五味子乙素可抑制A549细胞长期的增殖(见图3 a).划痕实验结果表明,五味子乙素可抑制A549细胞的迁移(见图3 b).

**.与对照组比较P<0.01.图3黄芪甲苷抑制肺癌A549细胞增殖(a)和迁移(b)Fig.3Astragaloside IV inhibits proliferation (a) and migration (b) of lung cancer A549 cells
3 讨 论
肺癌是最常见的癌症,也是全球癌症相关死亡的主要原因[8].一般肺癌患者Ⅰ期或Ⅱ期手术切除术具有良好的预后,5 a生存率高达70%.然而,大多数(75%以上)患者在诊断时就已经为晚期,5 a生存率低于25%[9-12].在过去10 a中,晚期肺癌的治疗方式已从传统的化疗(如基于细胞毒性铂类的药物)发展为针对特定分子亚型的更有效方案,如显示表皮生长因子受体(EGFR)突变或间变性淋巴瘤激酶(ALK)融合癌基因的方案[13-14].尽管靶向治疗已在临床获益,但大多数患者由于新突变(例如EGFR T790M和C797S突变)或基因拷贝数增加(即ALK癌基因复制)而最终导致疾病复发[15-17].此外,对于没有分子定义的肺癌患者靶向治疗无效,这些没有分子定义的肺癌患者占肺癌病例的大多数[13-14].最近,针对程序性细胞死亡-1(PD1)、程序性细胞死亡配体-1(PDL1)和细胞毒性T细胞淋巴细胞抗原-4(CTLA4)的免疫检查点抑制剂应用到一线肺癌治疗方案中,提高了患者的生存率和生活质量;但只有不到20%的患者获得长期获益,大多数患者在治疗期间最终会死于进展性疾病[18].因此,迫切需要额外的治疗靶点和相应的天然小分子化合物以改善肺癌治疗结果.
信号转导和转录激活因子3(STAT3)属于STAT蛋白家族,它们既是信号换能器又是转录因子,目前已经确定了7个STAT家族成员,包括STAT1、STAT2、STAT3、STAT4、STAT5A、STAT5B和STAT6[14].STAT3的结构是STAT家族的一个代表特征,包括N端结构域、线圈结构域、DNA结合结构域、连接子、SH2结构域和C端反激活结构域[19].DNA结合结构域使STAT3和DNA之间能够形成复合物,SH2结构域参与两个磷酸化STAT单体的二聚化.同时,C末端结构域包含一个酪氨酸(Tyr705)残基和一个丝氨酸(Ser727)残基,其中酪氨酸(Tyr705)残基对于SH2结构域介导的二聚化和STAT3的激活至关重要.丝氨酸(Ser727)残基可促使STAT3的转录活性最大化,STAT3在超过50%的非小细胞肺癌患者中持续激活[20],其表达增加与肿瘤分化不良、晚期临床分期、淋巴结转移和耐药性相关[21].目前研究[21]显示:多种细胞外蛋白可与细胞膜表面的特异受体结合,通过酪氨酸激酶(常为JAK家族成员)使受体酪氨酸磷酸化而活化,进而激活胞质中的STAT3分子,这些STAT单体形成同源或异源二聚体,其作为转录因子进入细胞核调控靶基因转录,促进肺癌增殖和迁移.本研究通过筛选发现:黄芪甲苷可抑制A549细胞中JAK/STAT3信号通路,且该作用在黄芪甲苷5、10及20 μmol/L 3个剂量间具有量效关系,同时黄芪甲苷可抑制A549细胞的增殖和迁移.
黄芪甲苷对肝癌SK-Hep1、Hep3B细胞的增殖具有抑制作用,研究[22]表明:200 μmol/L黄芪甲苷能够使肝癌细胞G1周期阻滞,进而诱导细胞凋亡,并抑制其迁移.本研究结果显示:2~20 μmol/L的黄芪甲苷对A549细胞具有抑制增殖作用,提示黄芪甲苷对不同肿瘤细胞的增殖影响可能具有选择性和特异性.在后续工作中,我们将选用多种肿瘤细胞及正常细胞,进一步对黄芪甲苷抑制增殖的剂量进行观察,以期明确是否对肺癌细胞具有特异性.
综上所述,黄芪甲苷通过JAK/STAT3信号通路抑制了肺癌细胞增殖和迁移,本研究可为黄芪资源的合理应用及抗肺癌中药的开发提供理论依据.
