胆固醇负载的环糊精对冻融延边黄牛精子鱼精蛋白损失和凋亡的影响
2022-11-17吕艳秋徐海峰曲星霖张雨阳李春宇顾伟玉
吕艳秋,徐海峰,曲星霖,张雨阳,李春宇,顾伟玉,金 一*
(1.延边大学东北寒区肉牛科技创新教育部工程研究中心,吉林延吉 133002;2.长春市第一中等专业学校,吉林长春 130000)
延边黄牛是国家资源保护品种,是中国五大地方精品牛之一,提高延边黄牛的繁殖力对分子育种计划十分重要。精液的冷冻和解冻会对精子质膜的渗透性、顶体和DNA 的完整性有很大影响,这些变化导致mRNA 和蛋白质的降解,从而影响生殖能力[1]。甘油是主要的渗透型冷冻保护剂,保护精子免受冰点以下的热休克,提供 “盐缓冲”的机制,可与金属离子结合,使细胞脱水,降低冷冻压力,并减少凝固过程中精子内部的冰晶体积,从而防止精子损伤[2-4]。解冻后牛精子活力和受精能力受冷冻保护剂中甘油浓度的影响[5]。研究表明牛精子冷冻保存过程中甘油最佳浓度为6%[6],但有报道称甘油会破坏精子的核膜,且浓度高于4%时会影响精子质膜流动性[7],进而导致受精能力下降。
胆固醇是细胞膜的主要结构成分[8]。Mocé 等[9]研究发现通过添加环糊精以提高胆固醇:磷脂的比例,和甘油有着相似的作用,即使精子细胞保护免受冰点以下的热休克。胆固醇负载的环糊精(CLC)为非渗透型冷冻保护剂,已被广泛用于公猪[10]、公牛[11]、种马[12]等动物精子保存,以提高精子活力。本研究在冷冻保护剂中用CLC 替代部分甘油,探究在降低甘油毒性后,不同浓度CLC 对冻融延边黄牛精子质量的影响,为进一步完善牛精子冻融损伤机理和完善冷冻保护液配方提供理论依据。
1 材料与方法
1.1 实验材料
1.1.1 主要试剂 微量总RNA、cDNA 和荧光定量PCR提取试剂盒购置美国Thermo Fisher 公司。胆固醇、甲基-β-环糊精、Tris、柠檬酸钠、葡萄糖、青霉素、硫酸链霉素、甘油、CMA3、七水合磷酸氢二钠、氯化镁等试剂购置美国Sigma 公司。
1.1.2 CLC的制备采用Purdy和Graham[13]法制备CLC。简单地说,将200mg 胆固醇溶解于1 mL 氯仿中,混合后,吸取0.45 mL 加入1 g 甲基-β-环糊精溶解的2 mL 甲醇溶液中,在电击仪中完全融合。然后倒入干净的无菌玻璃血中,放入干燥箱中,37℃烘干。干燥后,取出载有胆固醇的环糊精(CLC)晶体放入干净的玻璃瓶中,22℃保存。使用前,将50 mg CLC 加入1 mL TALP[14]溶液中,混匀后使用。
1.2 实验方法
1.2.1 精液的收集与冷冻解冻程序 选择5 头3~5 岁、健康无疾病的延边黄牛混合精液,以消除个体差异,使用假阴道法每周采集2 次,仅收集活力≥85%精液进行试验。为评价不同浓度的CLC 对牛精子的影响,先配制不含甘油的Tris 稀释液(Tris 2.42 g、柠檬酸钠 1.35 g、葡萄糖 1.1 g、青霉素 0.08 g、硫酸链霉素 0.1 g 和卵黄20 mL,双蒸水定容至100 mL),之后将含有甘油和不同浓度CLC 的Tris 稀释液将精子稀释至1.2×108个/mL,鲜精组使用PBS 进行稀释。具体试验分组为鲜精组、6%甘油对照组(无CLC,甘油6 mL,Tris 稀释液94 mL)、3%甘油组(无CLC,甘油3mL,Tris稀释液97mL)和3%甘油+不同浓度CLC组(0.75、1.5、3.0 mg/mL CLC,甘油3 mL,Tris 稀释液97 mL)。为了使CLC最佳地掺入到精子膜中,用CLC 处理过的样品在22℃孵育15 min,在精子冷冻前600 r/min 离心8 min 取出CLC,之后再用只含有3%甘油的Tris 稀释液重悬(不含CLC),为了与处理组一致,对照组使用含有6%甘油的Tris 稀释液进行相同的处理,使各组精子最终浓度为 80×106个/mL(pH=7.0,300 mOsm)。然后,在室温22~25℃,将精子样本灌注到0.25 mL 法式细管,平衡在4℃,3.5 h。在液氮上方5 cm 静液氮蒸汽中熏蒸8~10 min,然后放入液氮中保存。解冻时将冷冻的细管在37℃的水浴中解冻30 s,用于研究精液的不同参数。
1.2.2 精子活力的评估 将精子样品置于已预热(37℃)的载玻片上,通过计算机辅助精子运动分析技术(CASA,中国,同方)对精子进行评估。简而言之,对于每个样品,将5 μL 精液放在分析仪的腔室中,该腔室在分析过程中保持在37℃。随机选择5 个视野,至少计数200 个精子,检测精子活力。
1.2.3 精子顶体完整性的评估 顶体完整性通过考马斯亮蓝染色进行检测,取100 μL 稀释后的精液涂于载玻片上,再取1 mL 考马斯亮蓝染色液将精子铺盖,避光下染色30 min,随后镜检(400×),至少计数200 个精子,检测精子顶体完整率。其中精子头部受损被认为是顶体不完整的精子,精子头部完整被认为是顶体完整的精子。
1.2.4 鱼精蛋白损失的评估 精液在不含Ca2+和Mg2+的磷酸盐缓冲盐水(PBS)中洗涤2 次,并在4℃条件下于固定液(甲醇: 冰醋酸=3:1)中固定5 min。涂片,将每张载玻片用100 μL CMA3 染色工作液(在McIlvaine 缓冲液中加入0.25 mg/mL(0.1 mol/L 柠檬酸7 mL +0.12 mol/L 七水合磷酸氢二钠32.9 mL,pH 7.0,其中包含氯化镁10 mmol/L)避光染色处理20 min。然后将载玻片在McIlvain 的缓冲液中冲洗并风干。随后镜检(400×),至少计数200 个精子,检测精子鱼精蛋白损失率。其中精子头部亮绿色荧光染色质结构异常被认为鱼精蛋白损失,精子头部暗绿色荧光染色质结构正常被认为鱼精蛋白未损失。
1.2.5 基因表达量的评估 实时定量聚合酶链反应(RTqPCR)用于使用寡核苷酸引物序列评估转录本丰度,所用引物由生工生物工程(上海)股份有限公司合成,所有序列均列于表1。简而言之,根据制造商推荐方案使用DynabeadsTMmRNA DIRECT Kit 将用PBS 清洗3次后的精子细胞(1×107个/mL)提取总RNA,并用分光光度计测量其浓度;使用RevertAidTM First Strand cDNA Synthesis Kit 合成互补DNA,置于 PCR 仪中,设置程序为50℃,30 min,85℃,5 min;最后使用Fast SYBRTMGreen Master Mix 制备反应体系,每20μL反应体系中含cDNA(1 500 ng/μL)2 μL,2×SYBR Green(1X)10 μL,上下游引物各1(10 pmol/L)μL,ddH2O 6 μL。采用PCRmax Eco 48 实时PCR 系统进行RT-qPCR 分析,PCR 扩增步骤为:95 ℃10 min,然后95℃变性15 s,54℃退火15 s,72℃20 s,40 个循环,每个反应体系进行3 次独立生物学重复。并使用公式2-ΔΔCT法计算基因的表达量。

表1 RT-qPCR 引物序列
1.3 统计分析 所有试验均重复五次,使用统计学软件包SPSS 25.0 进行单因素ANOVN 检验。结果以平均值±标准差表示,当P<0.05 时,差异为显著。
2 结果
2.1 用CLC 替代部分甘油对冻融精子活力、顶体膜完整性和鱼精蛋白损失率的影响 如图1 所示,精子通过色霉素A3(CMA3)和考马斯亮蓝染色。由表2 可以看出,与对照组相比,3%甘油组精子活力和顶体完整性显著下降,鱼精蛋白损失率显著升高,3%甘油+CLC(0.75、1.5、3 mg/mL)处理组精子活力显著升高,鱼精蛋白损失率显著下降,且CLC 浓度为1.5 mg/mL 时效果最佳。因此后续试验将使用浓度为1.5 mg/mL 的CLC 进行研究。

表2 CLC 替代部分甘油对冻融精子活力、顶体完整性和鱼精蛋白损失率的影响
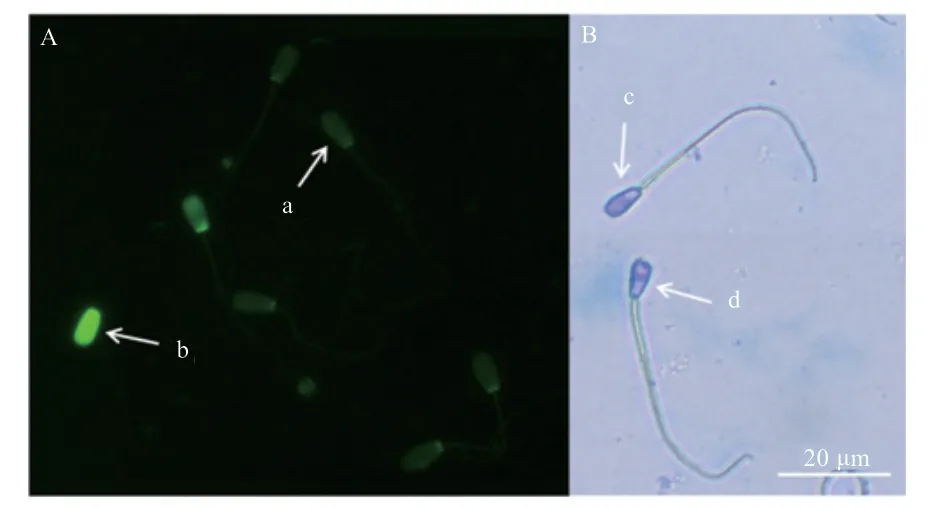
图1 冻融前后精子鱼精蛋白损失和顶体完整性的镜检
2.2 用CLC 替代部分甘油对冻融精子鱼精蛋白基因的影响 由图2 可以看出,经过冻融处理的精子鱼精蛋白基因PRM2和PRM3表达量显著下降,同时,与对照组相比(1.04%),3% 甘油组PRM2(0.72%)和PRM3(0.56%)表达量显著下降,3%甘油+1.5 mg/mL CLC组显著升高(1.92%、1.89%)。

图2 冻融前后精子PRM2、PRM3 相对表达量
2.3 用CLC 替代部分甘油对冻融精子顶体完整性和氧化应激相关基因的影响 由图3 可以看出,经过冻融处理的精子顶体完整性相关基因SPACA3表达量显著降低,氧化应激相关基因ROMO1表达量显著升高。与对照组相比(1.02%,1.07%),3%甘油组SPACA3(0.94%)表达量显著下),ROMO1(1.32%)表达量显著升高,3%甘油+1.5 mg/mL CLC 组SPACA3(1.26%)表达量显著升高,ROMO1(0.92%)表达量显著下降。
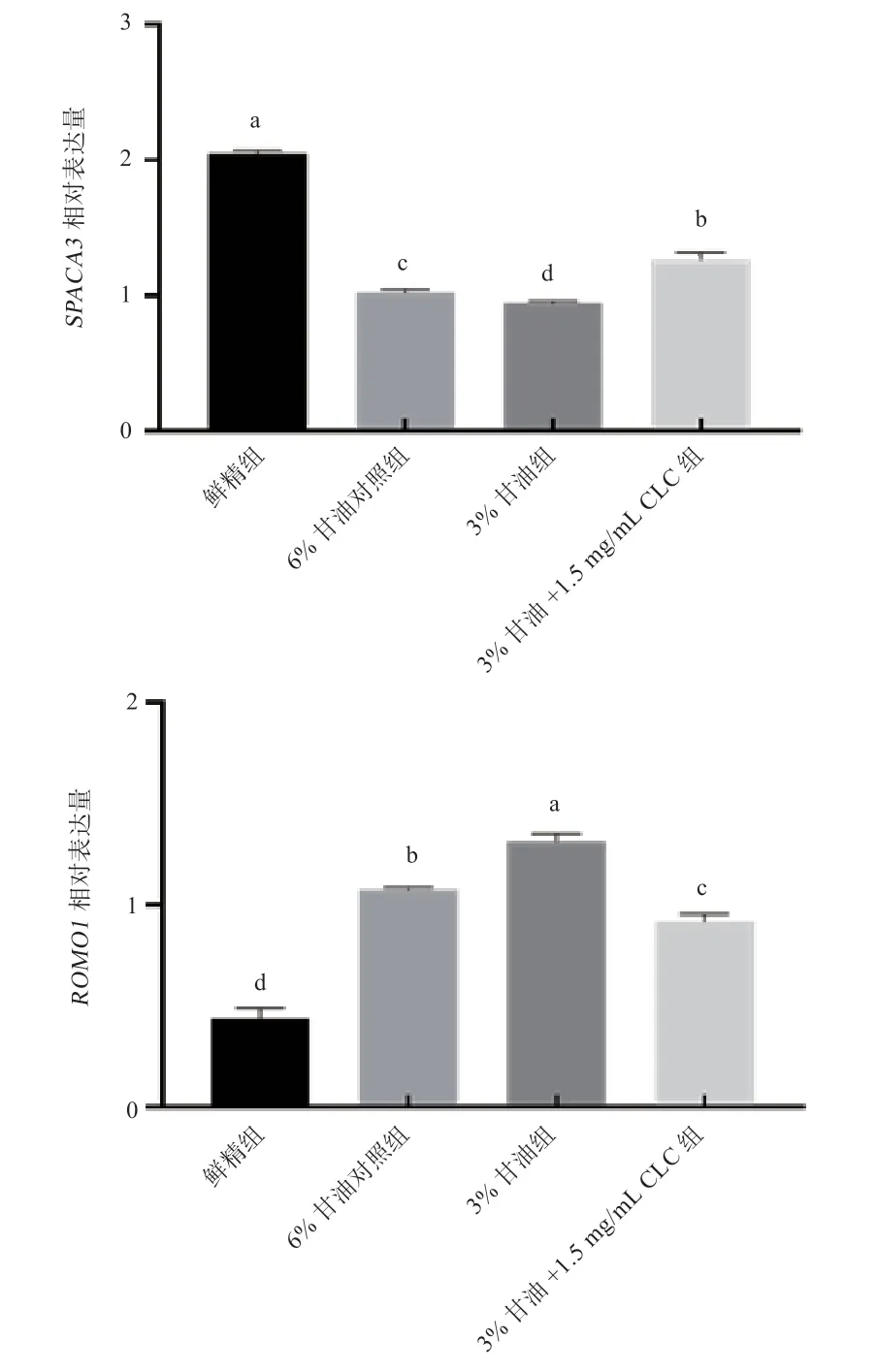
图3 冻融前后精子ROMO1、SPACA3 相对表达量
2.4 用CLC 替代部分甘油对冻融精子凋亡相关基因的影响 由图4 可以看出,经过冻融处理的凋亡相关基因BCL2和BCL2/BAX表达量显著降低,BAX表达量显著升高。与对照组相比(1.06%,1.04%,1.02%),3%甘油组BCL2和BCL2/BAX(0.76%,0.62%)表达量显著下降,BAX(1.22%)表达量显著升高;3% 甘油+1.5 mg/mL CLC 组BCL2和BCL2/BAX(1.61%,3.07%)表达量显著升高,BAX(0.51%)表达量显著下降。

图4 冻融前后精子BCL2、BAX、BCL2/BAX 相对表达量
3 讨 论
3.1 用CLC 替代部分甘油对冻融精子活力、顶体膜完整性的影响 即使随着冷冻保存技术的进步,冷冻保存仍然会对精子造成损害,低温保存通过对精子膜、细胞骨架、运动器官和细胞代谢等产生影响,降低精子的生育能力[15]。精子的活力、顶体完整性和抗氧化状态等各种参数与生育能力有关,Shiva 等[16]研究表明所有这些精子属性都受到冻融过程的影响。Lodhi 等[17]报道解冻后正常形态精子的活力和膜完整性之间有显著相关性。本实验也得到相同结果,即经过冻融处理的精子活力显著下降,在3% 甘油中添加1.5 mg/mL 的CLC 后精子的活力和顶体完整性得到显著改善,且精子活力和顶体完整性均优于对照组,结果说明甘油浓度降低至3%后添加CLC 处理可改善精子活力和顶体功能,同时精子活力提高可能是由于在冷冻过程中精子受到CLC 对精子膜的保护作用。
3.2 用CLC 替代部分甘油对冻融精子鱼精蛋白损失率的影响 鱼精蛋白(PRM)是精子核中的主要蛋白质,在其正常功能中起着重要作用,包括 DNA 的结合过程[18]。PRM 是在精子发生阶段形成的[19],在此期间,精子核中发生蛋白质替代过程,在最初支配精子核的组蛋白,通过复杂的过程被 PRM 取代,例如甲基化、磷酸化和泛素化[18]。PRM 将优化包装精子 DNA 以增加染色质凝聚,这将保护父本基因组的遗传完整性,免受核酸酶、诱变剂和其他可能损害 DNA 的因素的影响。本实验中,与对照组相比,在3%甘油中添加1.5 mg/mL CLC 的精子鱼精蛋白损失率显著降低。Miikeda 等[20]研究发现,牛精子中PRM1、PRM2和PRM3的鱼精蛋白表达对精子的正常功能发挥重要作用,所以3%甘油+1.5 mg/mL CLC 处理组鱼精蛋白水平的改善可能是由于鱼精蛋白相关基因起到了绝对的作用,为证实这一研究假设,本实验又对鱼精蛋白PRM2和PRM3基因进行了检测。
3.3 用CLC 替代部分甘油对冻融精子鱼精蛋白、凋亡、顶体完整性和氧化应激基因的影响 精子是一种特殊细胞,其功能取决于蛋白质的激活[21],受精前蛋白质的翻译后修饰在获能受精过程中起关键作用[22]。多年来,人们一直认为成熟的精子细胞中不会发生蛋白质翻译,第一,精子发生过程中组蛋白被鱼精蛋白取代而导致的紧凑的精子核使转录机制无法进入基因组;第二,大多数细胞器的损失,包括内质网和核糖体,限制或阻止了翻译活动。尽管有这些观察结果,但在人类精子中已经鉴定出几种类型的编码和非编码 RNA[23]。研究发现,哺乳动物的精子不仅具有合成线粒体编码RNA 的能力,而且还含有核编码的mRNA 和蛋白质,在获能过程中,精子可以替代降解的蛋白质或合成新的蛋白质,这些蛋白质对受精至关重要[24]。在冻融犬精液中发现鱼精蛋白PRM2和PRM3基因会发生变化[25-26],同样在本研究中发现牛精液PRM2和PRM3基因也发生了显著变化,在3%甘油+1.5 mg/mL CLC 组PRM2和PRM3表达量显著增加,验证了我们的假设,与CMA3 染色结果一致,说明添加CLC 替代部分甘油在冷冻保存期间减少精子鱼精蛋白的损失,是由PRM2和PRM3大量聚集而阻止了精子鱼精蛋白的损失。同时值得注意的是,在冷冻的种马精子中PRM2基因表达量与对照组相比显著下降[27],这可能是不同物种的原因。细胞凋亡,也称为选择性细胞死亡,是一种影响细胞活力的现象,该过程已在人类精子中被记录,并与辅助生殖技术后的受精失败有关。如今,精子的冷冻和解冻通常与宫内受精和体外受精等辅助生殖技术结合使用,但冷冻和解冻过程会增加凋亡精子的数量,从而降低辅助生殖技术的成功率。因此,应仔细评估当前精子的冷冻和解冻方法,以确保最大限度地减少细胞凋亡诱导的损伤,对判断冻融后的精子能够保持其受精能力是十分重要的[28]。据报道[29],BCL2超家族的成员是线粒体凋亡的关键调控因子,该超家族可分为促凋亡成员和抗凋亡成员,促凋亡BCL2成员,如BAX和BAK 主要位于细胞质中,可以根据需要而插入到线粒体外膜(OMM)中;抗凋亡成员,如BCL2本身和BCL-XL 位于 OMM 中,可通过阻止与促凋亡BCL2成员相关的电压依赖性阴离子通道(VDAC)的开放,也可通过抑制促凋亡BCL2成员形成的超分子开口的组装来抑制细胞凋亡[30]。精子尾中段大量的线粒体为细胞内细胞凋亡应激提供可能性[31]。在本实验中,3% 甘油+1.5 mg/mL CLC 组与对照组相比,抗凋亡基因BCL2的转录表达增加和促凋亡基因BAX的转录表达降低,BCL2/BAX的值显著升高,这与精子活力的检测是一致的,说明冷冻保护液中添加CLC 替代部分甘油在冷冻保存期间减少了细胞凋亡,保护精子发生。ROMO1是产生线粒体ROS 的关键基因[32],线粒体呼吸链中产生大量内源性ROS,可引起基因随机突变并导致程序性细胞死亡[33]。在本实验中,与对照组相比,3% 甘油+1.5 mg/mL CLC 组ROMO1基因转录表达降低,说明冷冻保护液中添加CLC 替代部分甘油在冻融过程中可保护线粒体,并减少ROS 的产生。SPACA3基因存在于质膜周围细胞外基质中的N-乙酰氨基葡糖寡糖的结合位点,可编码SPRASA 蛋白进而参与精卵细胞的识别和结合,并在生育力中起绝对作用[34-35]。研究结果显示与对照组相比,3%甘油+1.5mg/mL CLC 组的SPACA3基因转录表达增加,说明冷冻保护液中添加CLC 替代部分甘油在冷冻保存期间可使精子保持其正常的生理机能。这也有可能是细胞凋亡减少和线粒体保护增加的原因。
4 结 论
本研究结果显示,减少3%甘油浓度后添加1.5 mg/mL CLC 可显著改善冷冻保存精子质量。
