玉米ZmGly3基因的生物信息学与表达特性分析
2022-11-17于秋鸿曾黎宋希云李军
于秋鸿,曾黎,宋希云,李军
(青岛农业大学农学院,山东 青岛 266109)
干旱和盐渍是制约作物产量、品质和农业可持续发展的重要影响因素[1]。研究表明,在遭受干旱、盐胁迫等逆境时,植物细胞会积累过量的甲基乙二醛(methylglyoxal,MG)[2,3]。MG在低浓度下可作为一种信号分子,参与种子萌发、根发育、茎形态发生、细胞凋亡和胁迫耐受等多种生物学过程[4-8],但在高浓度下对细胞内的多种成分(如蛋白质、DNA、RNA和脂质)具有高度反应活性,导致它们不可逆地修饰或形成交联复合物,引发细胞毒害[9]。
乙二醛酶系统(Gly1、Gly2和Gly3)在MG的降解清除中发挥着极为重要的作用。Gly1和Gly2需要还原型谷胱甘肽(GSH)作辅助因子将MG降解为无毒的乳酸,而Gly3可以不依赖GSH的协助完成对MG的直接降解[10]。目前普遍认为Gly1-Gly2途径在MG降解过程中是最有效的,如过表达Gly1和Gly2的转基因烟草、水稻和拟南芥均表现出较强的耐盐和抗旱能力[11-14]。但Gly1-Gly2途径需要GSH和金属离子作为其活性的辅助因子,因此当GSH不足时,Gly3才能在MG降解中发挥作用[2]。Gly3广泛存在于大肠杆菌[15]、裂殖酵母[16]和真核生物[17,18]中。已有证据表明,Gly3也可参与植物对生物和非生物逆境的应答[10,18-20]。
玉米是一种重要的粮饲兼用作物,生长过程中经常遭受干旱胁迫,而且近年来因全球气候变暖导致的热旱害日趋严重[21]。土壤盐渍化也在一定程度上限制了玉米的种植和推广。为此,我们从NCBI网站(https://preview.ncbi.nlm.nih.gov)检索到了玉米Gly3蛋白的编码基因(命名为ZmGly3),利用生物信息学和分子遗传学方法对ZmGly3的编码蛋白理化性质、空间结构和表达时空特异性等进行解析,以期找出该基因在玉米应答干旱和盐胁迫中的作用,为玉米抗旱耐盐育种提供可用的基因资源。
1 材料与方法
1.1 试验材料及处理方法
供试玉米品种为郑单958。将种子播于含有营养土和蛭石(体积比为3∶1)的花盆中,每盆1粒,置于青岛农业大学智能温室内(25℃)培养;待幼苗长至三叶期时分别用20% PEG600和125 mmol/L NaCl溶液进行模拟干旱和盐胁迫处理,每处理5株幼苗,重复3次。分别于处理0、12、24、48 h后选取玉米幼苗的根、茎和叶,快速置于液氮中,于-80℃保存备用。
1.2 试剂
RNA提取试剂盒购自TaKaRa公司;反转录试剂盒和荧光定量试剂盒购自南京诺唯赞生物科技有限公司;引物由北京擎科生物科技有限公司合成(表1)。

表1 qRT-PCR引物
1.3 生物信息学分析
从NCBI网站(https://preview.ncbi.nlm.nih.gov)检索到ZmGly3(登录号NM_001153455.1);用ProtParam tool(https://web.expasy.org/protparam)软件分析ZmGly3编码蛋白的理化性质;用TMHMM Server 2.0软件进行蛋白跨膜结构预测;用SOPMA(https://npsa-prabi.ibcp.fr)和SWISS-MODEL(https://swissmodel.expasy.org/)进行蛋白质高级结构分析;用MEGA 7.0软件对ZmGly3进行系统进化树分析。
1.4 总RNA提取、cDNA合成和表达模式分析
按照RNA提取试剂盒操作说明提取总RNA;用反转录试剂盒合成cDNA;用qRT-PCR分析表达模式,以Actin作为内参基因。
2 结果与分析
2.1 ZmGly3的理化性质分析
用ProtParam tool软件分析发现,ZmGly3全长cDNA序列为1164 bp,编码387个氨基酸,理论分子量为41.2 kD,等电点为5.37。对氨基酸组成分析发现,带负电荷的氨基酸(Asp+Glu)残基总数为43,带正电荷的氨基酸(Arg+Lys)残基总数为31;Ala含量最高,占11.9%;Trp含量最少,占0.9%。蛋白不稳定系数较低,为31.70,属于稳定蛋白。用ProtScale分析发现,此蛋白整体为亲水性蛋白,在第127个氨基酸残基处的数值为2.38,疏水性最高,在第251个氨基酸残基处的数值为-2.344,亲水性最高(图1a)。TMHMM Server分析发现,该蛋白不含跨膜结构域(图1b),为细胞质蛋白。用CD-Search分析发现,ZmGly3含有两个DJ-1/Pfp1结构域,属于谷氨酰胺氨基转移酶(GAT-1)超家族(图1c)。

图1 ZmGly3蛋白的亲、疏水性(a)、跨膜结构域(b)和保守区(c)分析
2.2 ZmGly3的结构分析
用SOPMA软件分析发现,ZmGly3蛋白的二级结构主要有α-螺旋、延伸链、β-折叠和无规则卷曲。其中,α-螺旋有124个氨基酸,占32.04%;延伸链有83个氨基酸,占21.45%;β-折叠有39个氨基酸,占10.08%;无规则卷曲有141个氨基酸,占36.43%(图2a)。用SWISS-MODEL软件分析发现,ZmGly3蛋白由三条链组成,无金属结合位点(图2b)。

图2 ZmGly3蛋白的二级结构(a)和空间结构(b)
2.3 系统进化树分析
用BLAST程序从NCBI数据库中检索了与ZmGly3蛋白同源的其他植物的氨基酸序列,选取氨基酸序列同源性≥60%的10种植物进行进化树分析。结果表明,ZmGly3蛋白与马蹄芒MlGly3同源性为95.09%,亲缘关系最近;与茶树CsGly3同源性为79.53%,亲缘关系最远(图3)。

图3 ZmGly3同源序列比对和系统进化树分析
2.4 ZmGly3组织特异性表达
在正常生长条件下,ZmGly3在玉米根、茎和叶中均有表达,但在茎中最高,叶中次之,根中最低(图4)。茎中ZmGly3的相对表达量分别是根和叶中表达量的7.2倍和2.9倍。

图4 ZmGly3在玉米不同器官中的相对表达量
2.5 ZmGly3在干旱和盐胁迫下的表达
在干旱胁迫下,ZmGly3在玉米幼苗根中的相对表达量随处理时间的延长呈现先升高后降低的趋势,处理24 h相对表达量达到最高值,是对照(0 h)的3.6倍;干旱胁迫降低ZmGly3在茎中的相对表达量,处理48 h的相对表达量仅为对照的70%;在叶中的相对表达量总体呈现升高趋势,处理48 h的相对表达量最高,是对照的2.2倍(图5a)。盐胁迫能促进ZmGly3在玉米幼苗根、茎、叶中的相对表达量,且均随处理时间的延长呈现先升高后降低的趋势,均在处理24 h达到最高值,分别是对照的2.0、1.3、1.8倍(图5b)。
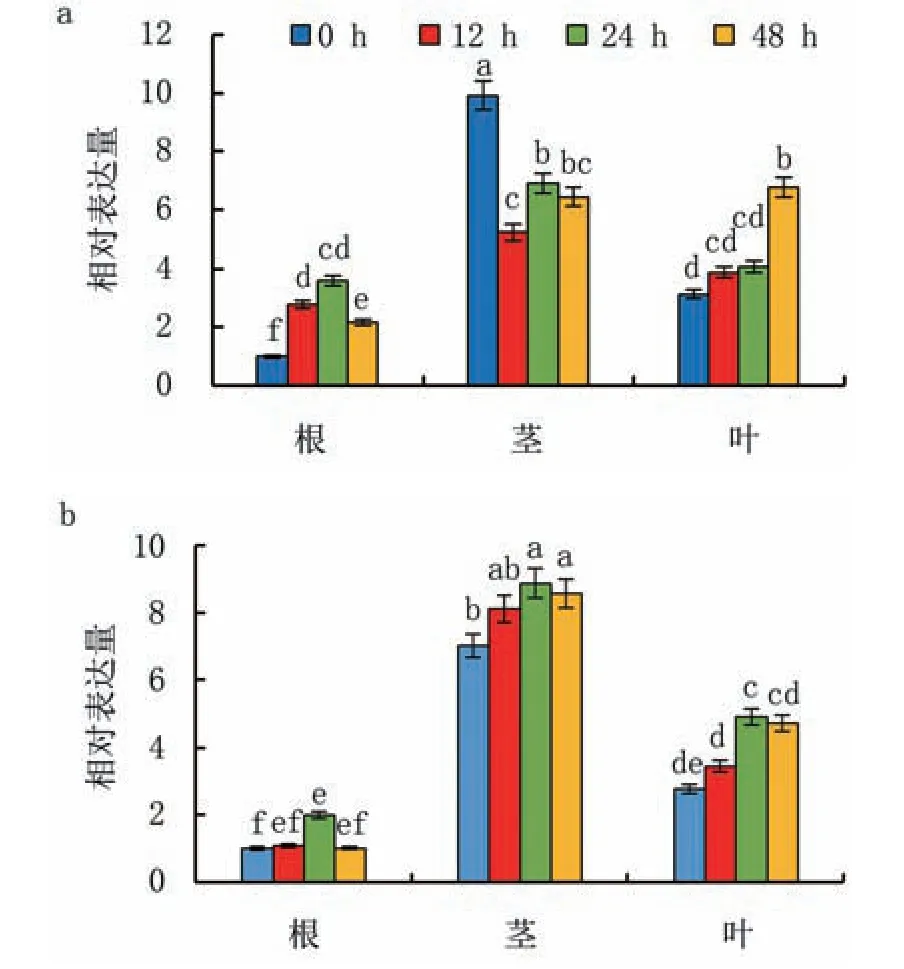
图5 ZmGly3在干旱(a)和盐(b)胁迫下的相对表达量
3 讨论与结论
Gly3参与的MG降解途径可以被认为是细胞内GSH不足或匮乏时清除过量MG的替代途径。Gly3的标志性特征是具有DJ-1/PfpI结构域,该结构域中保守的His-Cys-Glu/Asp是酶的活性位点,有MG降解活性但缺乏对GSH的结合亲和力;当Cys119定点突变为丙氨酸时,Gly3活性显著降低[2,17]。Gly3普遍存在于从低等(苔藓、石松)到高等(裸子、被子)的植物中[17]。另外,Gly3蛋白主要定位于细胞核、细胞质和叶绿体中[3,22]。我们利用生物信息学方法分析了玉米Gly3(ZmGly3)蛋白的理化性质和结构特点,结果表明,ZmGly3含有387个氨基酸,为稳定的亲水性蛋白;含有两个DJ-1/Pfp1结构域,是GAT-1超家族的成员;与Gly1和Gly2不同,ZmGly3没有金属结合位点;ZmGly3与马蹄芒MlGly3同源性最高,亲缘关系最近。
已有证据表明,Gly3参与了植物对逆境胁迫的应答。Ghosh等[17]对多种胁迫条件下的水稻DJ-1转录本研究发现,OsDJ-1(即OsGly3)受外源MG的强烈诱导,其中OsDJ-1C在不依赖于GSH的情况下将MG直接降解;OsDJ-1C在水稻的所有发育阶段和组织中表现出较高水平的组成型表达。Manoj等[10]发现在干旱和盐胁迫条件下抗性较强的野生近缘种甘蔗Gly3比商品种具有更高的表达水平。Meera等[19]研究发现盐胁迫下红树RmGly1和RmGly2的相对表达量明显高于RmGly3。Mohanan等[20]发现过表达EaGly3的甘蔗抗旱性增强。本研究结果表明,正常条件下,ZmGly3在玉米根、茎和叶中均有表达,但在茎中最高;在干旱和盐胁迫条件下,ZmGly3在根和叶片中的表达量增加,但在茎中的表达受干旱和盐胁迫影响不同,在干旱处理下降低而在盐处理下增加。
可见,ZmGly3参与了玉米对干旱和盐胁迫的应答,但在不同器官中对不同胁迫条件的响应并不完全一致。今后还需对其具体的生物学功能进行进一步解析。
