果实分类系统的修订和教学
——植物学教材质疑(十)
2022-11-16黎维平
黎维平
(湖南师范大学生命科学学院,中国湖南 长沙 410081)
果实是被子植物的特有结构,具有保护种子、帮助种子传播的作用,有些果实还能控制种子萌发[1]。果实以多样化的形态结构适应这些功能,其形态结构的复杂性给果实分类带来了一定困难。果实的植物学研究被称为果实学(carpology)和比较果实学(comparative carpology),甚至还有果实学家(carpologists)一词。目前,中外植物学教材[2~19]采用的果实分类系统较为一致。但是,其划分存在诸多不合理之处,有些果实类型的定义也不够严谨。此外,各教材对果实的介绍偏于简单枯燥,缺乏生物学理论背景。本文对传统的果实分类系统进行了一些修订,并分析了果实一些性状的相关性、演化及其生物学意义,旨在改进植物学教材的编写,提高果实教学的理论水平。
1 果实分类
中外教材[2~19]普遍从3个角度来划分果实类型。首先,从果实的来源,将果实分为单果(1花:1子房:1果)、聚合果(1花:多子房:多小果)和聚花果(1花序:多花:多果);其次,根据果实与子房的关系,将果实分为真果(果实完全来自子房)和假果(子房以外的结构如花托参与果实的组成);最后,根据果皮质地等特征,将果实分为肉果和干果及进一步多级划分。其中,根据果皮质地划分果实类型的分类问题尤其多,有必要加以改进。
1.1 聚合果的定义及其小果名称
离生雌蕊(apocarpous pistil)的每个子房发育成小果(fruitlet),小果组成聚合果(aggregate fruit)。聚合核果(drupetum或nuculanium)的小果被有的教材[6]称为小核果(drupelet)。聚合果中的小果多种多样,只对聚合核果中的小果命名,此“厚此薄彼”的做法有失“公允”。因此,其他聚合果,如聚合蓇葖果(follicletum或follicetum)、聚合瘦果(achenetum)、聚合坚果(aggregate nuts)和聚合浆果(baccacetum),其各类小果也有必要冠以相应名称,可分别称为蓇葖小果(folliclelet)、小瘦果(achenelet)、小坚果(nutlet)、小浆果(berrylet)。有的教材将胡桃科枫杨属(Pterocarya Kunth)和莎草科的由复雌蕊发育而成的坚果称为小坚果[20],或nutlet[21]。仅仅因为坚果较小而称之为小坚果有些不妥,这些所谓的小坚果与本文定义的小果不相干。蒲公英的下位瘦果也不宜称为“dandelion fruitlets[13]”。将牻牛儿苗科植物的分果爿(mericarp)称作“fruitlet[13]”同样是不合适的。还有文章[22]将发育不良而早落的苹果(Malus pumila Mill.)果实称为“apple fruitlets”,全然不顾术语的规范性,不应接受这类名称。
有些类群的合生雌蕊具次生性离生子房(secondary distinct ovary或secondary apo-ovary),其每个子房发育成1个小果,一朵花的所有小果是否也称为聚合果?这是一个未曾被提出,更未见讨论的问题。次生性离生子房见于唇形科、紫草科和萝藦亚科(夹竹桃科)。它们的花柱是合生的,所以不能说其雌蕊为离生雌蕊,只能称合生雌蕊有离生子房(apo-ovary)。当前,植物学教材[3,5~8]都将唇形科和紫草科的四分子房发育成的四小坚果归入分果类。但是,分果是单个子房在发育成果实后适应果实传播(避免同胞竞争)发生果瓣分离的现象,而四分子房的子房间部分分离至完全分离,果实成熟时无分裂或无明显分裂。此外,萝藦亚科植物的果实若被归入分果就更不合理了,因为分果的每个分果爿通常是不开裂以利用果皮保护种子、帮助种子传播,而萝藦亚科植物1个雌蕊的两个子房分别发育成开裂的蓇葖果。可是,Morris等[23]就将萝藦亚科的这种果实归入分果,称为“schizocarpic follicle”(分果类蓇葖果)。由次生性离生子房发育成的果实是放在分果类中,还是置于聚合果内,尚难有定论,有待进一步探讨。
1.2 假果
非子房组织(nonovarian tissue,noncarpellary tissue或accessory tissue)参与果实的形成,此果实就是假果(false fruit,accessory fruit或pseudocarp)[2~4,9]。单果和聚合果中既有真果(true fruit),也有假果;而聚花果(复果)均是假果,因为后者不可避免地有子房以外的部分参与果实的形成。苹果和草莓(Fragaria×ananassa Duch.)是植物学教材最常列举的假果例子[2~4,9,13]。在解释二者果实是假果时,各教材总要提到其供食用的肉质部分不是来自子房壁,这使得假果的定义出现两个误区。第一,有教材有意无意地将假果定义转变为子房壁以外的部分肉质化的果实,结果误导出如下推断:胎座肉质化的番茄(Lycopersicon esculentum Mill.)和猕猴桃(Actinidia chinensis Planch.)的果实是假果;具肉质假种皮的荔枝(Litchi chinensis Sonn.)和龙眼(Dimocarpus longan Lour.)的果实,也是假果[10]。事实上,番茄、猕猴桃、荔枝和龙眼都是真果。子房内任何部分肉质化都不是其所结果实被划归假果的依据。第二,在介绍假果时往往忽略了子房以外的部分参与果实形成而未肉质化的这类果实。所以,在讲解菊科果实时,学生较难接受其为假果的观点;蓼科酸模属(Rumex L.)植物内轮3萼片果期增大,包被果实,具有帮助果实风播的作用,此果应是假果,但是因其附属部分非肉质化,从未见将其称为假果。茄科植物花萼果期膨大,包被果实基部甚至整体,也未见将其归入假果的说法。在提到西瓜(Citrullus lanatus Matsum.et Nakai)的果实是假果时,必须强调其“假”不在于食用部分是胎座,而是其外果皮含有来源于花托的结构。
果实学作为一门学问,严谨性自然是必不可少的。如果说真果完全来自子房,那么花柱宿存于果实顶端时,该果实是否为真果?例如:毛茛科白头翁属(Pulsatilla Adans.)和蔷薇科仙女木属(Dryas L.)植物瘦果小,宿存花柱强烈增长成为羽毛状,帮助果实风播;牻牛儿苗科牻牛儿苗属(Erodium L′Hér.ex Aiton)分果瓣顶具发达的宿存花柱,其功能是帮助分果瓣自播并在其后钻入泥土中。发达的宿存花柱是子房以外的结构,参与了果实的形成,这种情况就不符合真果的定义。是否将真果定义扩大为雌蕊发育而成的果实?茄科植物花后花萼显著增大,宿存于果实基部,甚至包裹果实;稻属(Oryza L.)稃片包被颖果,当然不能把真果定义扩大到包括花被、稃片参与果实形成。严格说起来,这几种情况都应归为假果。这类问题还有待植物学同行共同探讨。
1.3 肉果的分类
各教材根据果皮质地,分出肉果和干果两类;肉果进一步被分成核果、浆果和梨果。
1.3.1 核果类
核果有时被定义为“通常由单雌蕊发展而成[3,8,16~17,19]”,这种将目光局限于蔷薇科梅亚科所导致的定义过于偏狭,因为除了提及椰子(Cocos nucifera L.)外,核果例子几乎全来自梅亚科[3,5,11,16]。实际上,核果在许多科独立地演化出来,是趋同进化的现象。大部分科的核果都是复雌蕊,如樟科、桑科、鼠李科、山矾科、桃金娘科、木樨科、茜草科、忍冬科、棕榈科等科植物具复雌蕊,其全科或部分植物的子房发育成核果。
有教材认为,枣(Ziziphus jujuba Mill.)的果实“有花盘参与发育,因此不是真正的核果,属于‘拟核果’[17]”。这是对核果定义的误解,核果的本质是中果皮肉质和内果皮骨质。荚蒾属(Viburnum L.)植物(原属于忍冬科,现移入五福花科)子房下位,其果皮的形成有花托参与,属于假果,但仍是核果。根据内果皮骨质这一特征,部分教材将椰子的果实归入核果[2~4]。矛盾的是,核果属于肉果,而椰子的果皮无肉质部分。所以,椰子的果实不应归入肉果中的核果,应列入干果中的闭果之内,可称为核果状坚果(drupelike nut)(表1)。
聚合核果中的小核果是核果类的另一亚型(表1)。教材普遍将草莓聚合果中的小果称为瘦果[2,5,7,13~14,16,18~19],马炜梁[6]纠正了这一错误,认定其为小核果。
核果类中有一类特殊果实为浆果状核果(表1),其特征为,果实成熟时内果皮分成几个单种子分核(1-seeded pyrenes),而中果皮肉质。这类果实见于五加科、冬青科、唇形科紫珠属(Callicarpa L.)、茜草科虎刺属(Damnacanthus Gaertn.f.)、五福花科接骨木属(Sambucus L.;原属忍冬科)以及蔷薇科山楂属(Crataegus L.)和火棘属(Pyracantha M.Roem.)等。将这类果实名称中加“浆果状”一词,是基于其有两个方面和浆果相同:1)中果皮多汁(吸引动物传播种子);2)整个果实内种子2至数枚(各种子有机会在传播过程中相互分离)。浆果状核果是核果的亚类之一(表1)。Gu等[24]称,火棘属的果实为“Pome……;pyrenes(nutlets)5”[梨果……;果瓣(小坚果)5],此处有两点值得商榷:1)火棘属的果实不是梨果,而是浆果状核果;2)将分核称小坚果不恰当。
一些应用性较强的杂志上发表的文章,常将中果皮多汁的核果称为浆果状核果[25]。中果皮多汁在核果中并不少见,为了避免混乱,建议不要将中果皮多汁的核果称为浆果状核果,而将该名称限于具分核者。
1.3.2 浆果类
1.3.2.1 浆果的定义
浆果被定义为“由一至多室的复子房形成,柔嫩、肉质而多汁,内含多数种子[6]”。这个定义下得有些武断,至少有两点值得商榷:1)浆果不止限于复子房,单子房也可发育成浆果,如五味子属(Schisandra Michx.)植物的聚合浆果;2)葡萄属(Vitis L.)植物的浆果具2~4粒种子,佛手瓜(Sechium edule Swartz)的浆果含1粒种子,均不适合称“种子多数”。所以,浆果可定义为:通常多为复雌蕊、稀单雌蕊发育成的肉质多汁、种子多数至1枚的果实。肉质化部分常为果皮,有时胎座(如西瓜;将胎座肉质化当成瓠果的特征[7,16,18]是以偏概全)或胎座及假隔膜(番茄等)也肉质化,但柑果肉质化部分为内果皮上的多细胞囊状毛(汁胞)。
一些教材[2,5,7,9~12,14~18]把柑果、瓠果与浆果并列,这样处理使果实分类过于散乱,三者合并成浆果为宜[3,6,8,13,19],而将普通浆果称真浆果(true berry)[13]。
1.3.2.2 建议将梨果并入浆果类
蔷薇科苹果亚科(已并入桃亚科Amygdaloideae)的特征性果实类型之一是梨果(pome)。梨果的特征为:1)果皮肉质部分来自被丝托(hypanthium;也称花筒floral tube);2)心皮发育成纸质或软骨质的内果皮,或描述为“薄薄的内果皮木质化[8]”;3)种子数枚。
一些教材[3~4,6,8,13,19]将梨果与核果、浆果并列成3类肉果。但是,本文建议将梨果并入浆果类。
梨果的肉质部分主要来自被丝托,来自心皮的外果皮、中果皮也肉质化,薄薄的内果皮木质化[8]或纸质至革质[13],这是各教材将梨果“自立门户”而与浆果并列的理由[9]。若因其为假果(肉质部分主要来自被丝托)就将其排除在浆果之外,那么葫芦科的瓠果也是假果,也该从浆果中分出。如果因为内果皮纸质、革质或木质而将梨果排除出浆果,那么,外果皮革质且为假果的瓠果和果皮非肉质化的柑果也不应归属于浆果内了。鉴定果实为浆果的两个要点是:1)果皮肉质多汁;2)种子常多于1枚。按照此标准,梨果就是浆果。所以,合适的处理是,考虑梨果的独特性(肉质部分主要来自被丝托,薄薄的内果皮木质化),将梨果当作浆果的亚类型之一,与真浆果、瓠果和柑果并列(表 1)。
在定义梨果时,一些教材[10,14,17~19]将山楂属植物作为实例之一。另有一些教材[5~8,11~12,15~16]将梨果作为苹果亚科的一个特征,并因山楂属和火棘属属于苹果亚科而默认其果实为梨果。山楂属和火棘属植物心皮5枚,熟时为骨质内果皮,5小核,每核具1种子(每室2胚珠,常仅1枚发育),得到骨质内果皮保护的种子仅具膜质种皮。此类果实中,中果皮肉质(来自被丝托);内果皮(来自子房壁)骨质组成分核。所以,山楂属和火棘属植物并不具梨果,而具浆果状核果(见上文)。
1.4 干果的分类
根据果皮开裂与否,干果被分为闭果和裂果[4]。这样处理使分果的地位有些“尴尬”,因为其既不像闭果,又不同于裂果。此外,各教材在对闭果和裂果进一步划分中也出现了一些不合理的处理。
1.4.1 分果应与闭果和裂果并列
分果被归入闭果,理由是其子房室成熟后不开裂(分果中的蓇葖果是例外)。实际上,分果应是介于闭果和裂果之间的中间类型,其兼具二者的优点:它们既像闭果那样,利用果皮保护和传播种子;又像裂果一样让果内各种子散布,即分果爿“各奔前程”,以免将一个果实内的几枚种子“捆绑”在一起而抵达一处,防止同胞竞争(sib competition)。所以,建议将分果与闭果、裂果并列,使这三者成为干果的一级划分。
蜀葵属(Althaea L.)果实呈盘状,分果爿有30枚至更多,成熟时与中轴分离,为典型分果。牻牛儿苗属果实也是分果。植物学教材中,无患子科的荔枝和龙眼的种子在介绍假种皮时总是被提及,而果实分类中荔枝和龙眼的果实则往往被忽略。该果实的分类难度较大:1)荔枝和龙眼的果实是非典型分果,一方面,其来自合生心皮发育而成果实的一个分果瓣,故它们的果实属于分果;另一方面,几个果瓣通常只有1个发育,故又与一般的分果不同;2)其果皮较薄,果实内发达的肉质化假种皮与浆果有点相似,但果皮的革质和种子1枚等与浆果显著不同,所以称之为浆果状分果(berrylike schizocarp)不太合适。根据果皮革质且较薄,称之为瘦果较为恰当。
一些教材通常将双悬果、双翅果和四小坚果包括在分果之内。为避免与聚合果中的小坚果一词重合,四小坚果一词可改称四分果爿(four mericarps)。蜀葵属植物的分果没有特殊名称,只好直呼其为分果(狭义的)。有的教材将双悬果和分果并列[7,10]而不说明二概念间的包含关系,不够合理。另一些教材,只提双悬果,不提分果[3,6,8],或将分果等同于双悬果[5,11~12,16,19],这也过于简单了。
如前文所说,四小坚果和分果类蓇葖果来自次生性离生子房,应属于聚合果还是分果类,有待进一步研究;本文暂时将其归在分果类名下(表1)。
豆科的节荚(loment)通常被置于荚果之中[3,6,8,10,12,16~19]。节荚与其他分果一样,传播时,种子仍被包被在果皮内,但果实横向断裂为含1粒种子的节片。其他分果行纵裂而节荚横列,二者无本质区别,在种子传播中具有相同的生物学意义。节荚加入分果后,分果的定义中就不能再认为其是由2至多心皮的子房发育而成,而应将心皮数的下限改为1。果实成熟时经纵裂或横列成数个具单种子的果爿,果爿是种子传播单元,这种果实就是分果。
十字花科某些植物的果实大部分是裂果中的角果。但是,高河菜属(Megacarpaea DC.)和臭荠属(Coronopus J.G.Zinn)植物的短角果成熟时果实纵裂成2果爿,每果爿内包裹1种子,此为典型的分果,可称为分角果。仿照分果的英文单词schizocarp,分角果的英文可为schizosilicle,即由前缀 schizo(断裂)和silicle(短角果)组成。
十字花科离子芥属(Chorispora R.Br.ex DC.)、四棱荠属(Goldbachia DC.)和隐子芥属(Cryptospora Kar.et Kir.)的果实也可像节荚那样分节横裂,每个节片内含1种子,可称之为节裂角果,英文为segmented silique,也可称节荚状长角果(lomentose silique)。像节荚一样,节荚状长角果也属于分果。
总之,分果是与闭果和裂果并列的3种干果之一。分果除了普遍归入其内的双悬果、双翅果、四小坚果之外,还应包括豆科的节荚、十字花科的分角果和节裂角果。教材关于十字花科果实只提长、短角果,其他变异从不提及,今后应加以改进。
1.4.2 裂果的分类
裂果常分为蓇葖果、荚果、角果和蒴果。蒴果是复雌蕊发育而成的裂果[6]。此定义包括两个标准:1)发育成蒴果的雌蕊为复雌蕊,以排除单雌蕊发育而成的蓇葖果和荚果等裂果;2)其为裂果。据此定义,角果理应置于蒴果之内,因为:1)角果来自2心皮的复雌蕊;2)角果是裂果。从进化上看,十字花科的姊妹科白花菜科有2心皮的蒴果,而角果除了增加了假隔膜外,其他方面都保留了白花菜科植物蒴果的特征。假隔膜的出现并未改变果实的性质,所以角果可置于蒴果内成为其亚类型之一(表1)。
1.4.3 闭果的分类
闭果之下的类型较为多样,有教材[3,8]将其分为瘦果、翅果、坚果、颖果、胞果和双悬果,另有教材还提及下位瘦果和小坚果[6]。如此一盘散沙似的罗列,损失了一些有价值的信息,不利于体现果实分类系统的逻辑性,更不利于教学。
根据果皮的厚薄、硬度等性质,闭果可分为瘦果和坚果两类。果皮较厚而坚硬的为坚果,其中有具翅坚果、核果状坚果;基部或整体被壳包被的坚果被称为槲果;此外,还可以包括归入分果、聚合果的小坚果(表1)。胡桃属(Juglans L.)植物果实的肉质外果皮由苞片、小苞片和花被片发育而成,子房发育成骨质果皮包被的“核”,一般称之为“核果[15,18]”“核果状坚果[6,10,20]”。笔者最初对此具肉质果皮的果实归入坚果而不是核果表示过不解。后来,笔者接受了胡桃属植物果实为坚果[12]的说法,因为肉质果皮在果实传播时已枯萎,该果实由以种子而不是果皮为食的啮齿动物传播[26]。其肉质果皮在果实传播中没有发挥作用,将胡桃属植物的果实归入坚果而称之为核果状坚果是恰当的。豆科花生(Arachis hypogaea L.)的子房钻入地下发育成果实,自然无法开裂,不属于裂果中的荚果;其果皮较厚较坚硬,种皮膜质,应属于坚果,英文称之为peanut是有道理的;唯一困扰笔者的是其果内种子常为2~3粒,不符合坚果只有单粒种子的一般特征。关于花生果实在果实分类系统中的地位有待进一步研究。
瘦果型果实将闭果中坚果之外的果型囊括其中。将瘦果局限于单心皮、上位子房发育而成的果实[2]是不合理的。龙眼和荔枝的果实被当作假果[10]当然不对,但不明确指出其属于何种果实也不恰当。笔者建议,根据其果皮较薄,将龙眼和荔枝的果实归入瘦果而不是坚果[2]。豆科植物中一些果实果皮不开裂、只含1粒种子,应置于瘦果之列。例如:两型豆属(Amphicarpaea Elliott ex Nutt.)闭锁花伸入地下结的果实、鸡眼草属(Kummerowia Schindl.)的果实,均已不具备荚果沿背缝线和腹缝线同时开裂的特征,应纳入瘦果范畴。十字花科单种属舟果荠属(Tauscheria Fisch.ex DC.)的“短角果”具翅,不开裂,种子1粒,显然此“短角果”实为瘦果。
菊科的果实因其发育自下位子房而单列为瘦果的一种特殊类型,称为连萼瘦果或下位瘦果[6](cypsela)[2,4]。颖果的果皮和种皮愈合,为禾本科特有,是瘦果的一种特殊类型。原藜科现属于苋科的一些植物具有胞果(囊果),有薄薄的果皮;因其来自复子房,果皮与种子极易分离,可仍保留胞果的名称,列为瘦果的亚型之一。
坚果和瘦果依据果皮的厚薄和坚硬程度加以区别。以此标准,有些果实易于归类,壳斗科、莲科的果实归入坚果,菊科和毛茛属(Ranunculus L.)的果实分别为下位瘦果和聚合瘦果,分类没有争议。但同一套教材,上册介绍瘦果时,所举例子提到蓼科的荞麦[3](Fagopyrum esculentum Moench),而下册认为蓼科植物具坚果[20];在介绍蓼科特征时,多数教材[5,7,11,14~15]记载为瘦果,另有教材[6,18]描述为坚果。这让学生不知所措。蓼科植物既有瘦果,也有坚果,如此表述更为恰当。蔷薇属(Rosa L.)的果实(蔷薇果,rosehip)为“多数瘦果集于肉质的被丝托内[6]”,但其小果的果皮坚硬(hard pericarp[27]),为骨质、革质或木质(bony,leathery or woody[27]),应属小坚果。
1.5 新的果实分类系统
基于以上的分析,笔者对传统的果实分类进行修订,提出了一个新的果实分类系统(表1)。该系统与传统的果实分类的不同之处在于:1)将果实分类区分为3~4个等级;2)在二级划分上,把肉果由过去的一分为三(核果、浆果和梨果)改为一分为二,即把梨果并入浆果;相反,将干果由传统的一分为二改为一分为三,把分果从闭果中分出而与之并列;3)在三级划分上,首先,将长、短角果并入蒴果;其次,将闭果内的众多果型归入坚果型和瘦果型;再次,分果型中加入了节荚、分角果、节裂角果;4)在裂果和闭果下划分出第四等级,翅果不再被单列为一类而分成具翅坚果和具翅瘦果(双翅果在三级划分时就已归入分果类),分属于坚果类和瘦果类,而不是笼统地认为其“与瘦果类似[8,17]”或“具瘦果性质[6,12]”或为“具翅的瘦果[18~19]”,更不必将其限于“上位子房发育而成[18]”。此外,翅果“果皮的一部分向外扩延成翼翅,如枫杨(Pterocarya stenoptera DC.)[15,17,19]”,这种说法通常是对的[16],但其举例恰巧是例外,枫杨的果翅来自小苞片而不是果皮的延伸结构。考虑到翅果多为瘦果,所以将samara一词给予了具翅瘦果[4];把颖果和胞果置于瘦果型内(表1)。
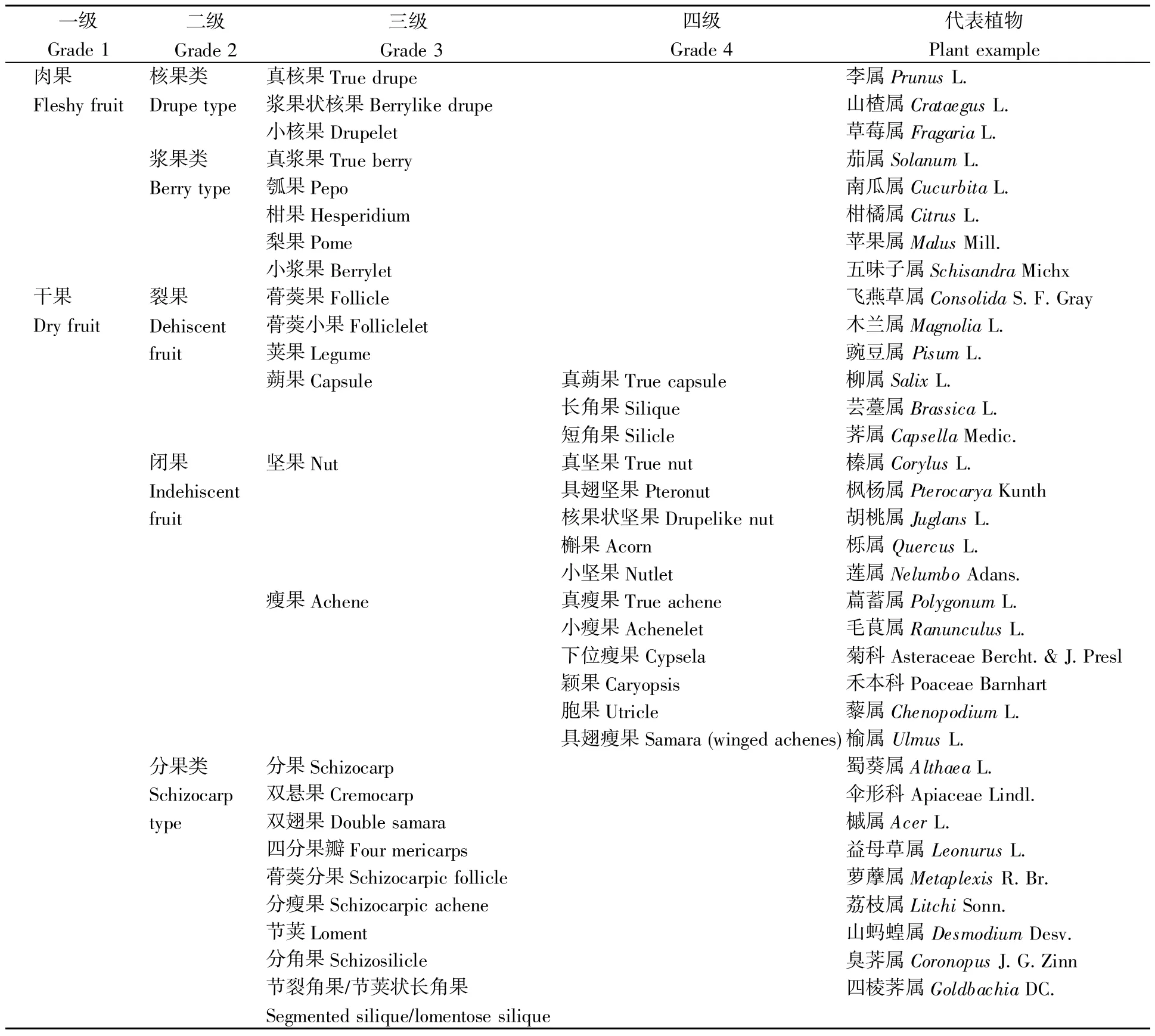
表1 基于果皮特征的果实类型分类Table 1 Classification of fruit types based on characteristics of pericarp
2 基于探究的果实分类教学
果实分类的教学若像开中药铺似地列举果实名称和特征,必定枯燥无味;若联系日常生活,固然生动活泼,但又稍嫌浅显,缺乏理论深度。结合果皮的功能分析果实各结构间的相关性,理解结构和功能间的关系,学习根据一种结构推测另一结构,这对于培养学生的学习兴趣和思维能力都会有不错的效果。
2.1 果实概念:从民间到植物学
植物果实是人类赖以生存的最重要的物质基础之一,与人们的日常生活密切相关,以至于每个学生从幼年开始就不断积累民间关于果实分类的知识。这些知识在脑海中根深蒂固,对学习果实的植物学知识有一些帮助,但也带来了不少负面影响。
银杏(Ginkgo biloba L.)的种子俗称白果(white nut),解释其为何是种子而不是果实,这是植物学教材和课堂教学必须解决的问题。解释虽不难,但不要忽略“白果”种皮结构与杏(Prunus armeniaca L.)果皮及种皮结构的相似性所包含的生物学意义。银杏种子的3层种皮分别为肉质、骨质和膜质,发达的胚乳含“生化武器”氰苷,与杏核果的中果皮肉质、内果皮骨质和种皮膜质以及发达的子叶含氰苷有精确的对应关系,可用趋同进化加以解释。
中小学语文课本上提到播种时,都将农民播撒的水稻(Oryza sativa L.)、玉米(Zea mays L.)的颖果说成种子。中学生物教材受多方面因素的限制,没能将果实与种子加以明确区分,如把水稻、玉米的颖果称作籽粒,将苍耳(Xanthium strumarium L.)的聚花果叫种子,这是对世俗的迁就。绝大多数人终身不学习植物学,这些错误将伴随其一生;如果学习植物学,这些扎根于脑中的知识也给学习果实的科学分类带来困难。况且,植物学教材在叙述果实传播时总要提到苍耳的果实,但并不明确说明其为聚花果;在解释聚花果时,也未见提到苍耳果。
食品中的市售坚果(commercial nuts)与植物学中的坚果(botanical nuts)相差很远。有科普网站(中国农业信息网:http://www.agri.cn/V20/SHXX/jksp/202012/t20201202_7569434.htm;2022-09-21)宣称,“坚果一般分为两类:一是籽坚果,包括花生、南瓜子、葵花籽、西瓜子;另一类是树坚果,包括核桃、杏仁、开心果、板栗、榛子、松子、腰果、碧根果、白果等”。事实上,白果、松子、南瓜子和西瓜子都是种子,葵花籽是下位瘦果,杏仁和开心果是核果的核,腰果是槭树科腰果(Anacardium occidentale L.)花托肉质膨大而成、托于核果下的棒状或梨形结构(并非子房的部分),真正的坚果只有核桃、花生、碧根果(美国山核桃Carya illinoensis K.Koch的果实)和榛子(Corylus heterophylla Fisch.ex Trautv.的果实)。
严格讲,西王从高速手里接掌山东男篮的时机并不好。由于是国企资产转让给民企,转让过程和变更手续耗费时日和精力过多,使西王无暇把工作重心及时转向球队备战新赛季,而东家交替给原管理层特别是教练班子带来的人心浮动,则极大地干扰了球队的正常训练。据知情人士透露,球队夏训形同虚设,体能储备严重不足,技战术磨合无从谈起。
英文中,民间果实名称与植物学果实分类的矛盾也十分常见,如 mulberry(桑葚)、strawberry(草莓)、raspberry(覆盆子)和 blackberry(黑莓)均与berry(浆果)无关[13]。
“叫板”民间植物知识是植物学教学不容回避的任务。果实的民间分类和植物学分类的差异,可列为学生的课后作业。
2.2 各层果皮间的分化及其生物学意义
果皮通常分外、中和内3层。分析各层果皮间的关系,有利于理解果实的结构和功能。
核果的果皮分为膜质的外果皮、肉质的中果皮和骨质的内果皮。中果皮肉质,富含水分,所以需要膜质的外果皮保护,减少水分散失。来自外果皮或中果皮的鲜艳色彩、中果皮的肉质结构能吸引动物捕食;为防止动物取食过程中及消化过程中对种子的伤害,内果皮骨质(通常由石细胞组成)以保护种子[9]。三层果皮相互配合,共同服务于种子的保护与传播。
干果中的裂果在开裂前,中果皮由薄壁细胞组成,内果皮为不连续的厚壁组织(李扬汉[5]指出其内果皮为厚壁组织,但未提及厚壁组织不连续这一关键性状)。裂果开裂的机制为果皮不均匀加厚导致不均匀收缩,进而在厚壁组织间断处开裂。换句话说,果实的开裂过程为:1)中果皮的薄壁组织失水,导致中果皮收缩;2)内果皮的厚壁组织由坚硬的死细胞组成,不会像薄壁组织那样失水收缩,但会因为中果皮的收缩承受向外翻卷的拉力;3)厚壁组织受到外侧拉力后,间断处的薄壁细胞破裂,果皮开裂。果实是沿背缝线开裂还是沿腹缝线开裂,取决于内果皮的机械组织在何处间断。夹竹桃科9属植物蓇葖果的果皮结构:外果皮和中果皮为表皮组织和薄壁组织,内果皮为多层厚壁组织;果实裂口(dehiscing zone)为内果皮厚壁组织的间断处[28]。该裂口可类比于真蕨类植物孢子囊和被子植物花药药室内壁的唇细胞,内果皮的厚壁组织在功能上等同于孢子囊环带和花药纤维层的加厚的壁,其外果皮和中果皮失水收缩扣动开裂的“扳机”,此功能相当于环带和纤维层外切向壁失水收缩导致孢子囊和花药开裂。果实、孢子囊和花粉囊分享类似的开裂机理。
分果和裂果开裂具有类似的机理。但是分果的种子开裂后并不暴露,这是因为在纵裂类型中,其内果皮的机械组织(厚壁组织)在心皮内连续,而在心皮间由薄壁组织隔开,这些薄壁组织为裂口;在分果的横列类型中,内果皮的机械组织纵向间断,即在纵向排列的种子间,内果皮机械组织间断而为薄壁组织填充,此处为果实断裂处。此结论来自笔者对合萌(Aeschynomene indica L.)等节荚果皮纵切面的观察(黎维平,未发表)。
闭果则在结构上无裂口,不发生不均匀收缩。例如,紫菀族植物瘦果果皮均为薄壁细胞[29],成熟过程中均匀枯萎收缩,形成干燥闭合果皮。
2.3 果皮特征和种子传播方式的相关性
本系列论文的第7篇曾指出,果实和种子不是生殖器官,而是传播体[30]。肉果及分果的一个果爿、裂果的一粒种子和一个闭果都是一个种子传播单元(seed-dispersing unit)。果实存在的价值在于其为种子提供保护和帮助种子传播[9]。实现这些功能的途径是多样化的,果皮的特征也是多种多样的。
果实和种子肉质化的结构通常是用于吸引动物以帮助果实、种子传播。肉质的果皮(肉果中的浆果、核果)、花托(假果)、花序轴(聚花果)都可对动物有吸引力。而种子具肉质假种皮[如卫矛属(Euonymus L.)蒴果]、种皮脂质体[4][见堇菜科堇菜属(Viola L.)和罂粟科紫堇属(Corydalis DC.)]和肉质外种皮(如木兰属Magnolia L.)时,其果实通常开裂,暴露出种子上的肉质结构以吸引动物帮助传播种子。例外见于无患子科的龙眼属(Dimocarpus Lour.)和荔枝属(Litchi Sonn.),这些植物的果实通过假种皮吸引动物[31],但果皮并不开裂,也许是以往的经验或芳香气味帮助传播动物识别这些果实。
动物被动传播果实(闭果、分果)和种子(裂果)时,果皮或种皮具刺毛、黏液等,以附着动物身体而随之扩散。
依靠风力传播的果实是具翅坚果、具翅瘦果、双翅果、具宿存羽状花柱的瘦果。而借助风力传播的种子来自裂果,种子具翅、被毛。像风媒花的花粉一样,风播的种子通常小而轻[9,13],以兰科植物种子最为突出。
种子自播的传播力来自果皮(如凤仙花属Impatiens L.、大豆属 Glycine Willd.植物)或果实内部的压力(如葫芦科喷瓜Ecballium elaterium A.Rich.)。
2.4 果皮特征与种皮质地的相关性:保护功能的此消彼长
果皮特征对种皮特征有直接或间接的决定作用,其中,二者的保护功能具有此消彼长的相关性。肉果中的浆果和干果中的裂果,其内的种子都有厚而硬的种皮,以利于动物咀嚼、消化时以及扩散后对种子的保护[9]。裂果果实开裂后,其内的种子直接接触外界环境(包括生物环境和非生物环境),种皮厚而硬,也是起着保护种子的作用[9]。
相反,核果和闭果、分果的种子始终受到果皮的保护,种皮则退化为膜质。苏铁(Cycas revoluta Thunb.)和银杏的外种皮肉质、中种皮骨质而与核果相似,所以其内种皮退化为膜质,也可作同样的解释。苍耳聚花果中,2连萼瘦果被厚革质的总苞包裹,其总苞相当于果皮,而真正的果皮以及其内的种皮因保护功能大大弱化而退化成膜质。
毫不意外的是,总会出现果皮和种皮的关系与上述不相符合的例子:1)龙眼属、荔枝属植物的果实是闭果(瘦果),但因其具假种皮而被动物啃食,所以有坚硬的种皮保护胚;2)兰科植物具蒴果,可果内种子的种皮却为单层细胞的膜质结构,并不坚硬。这种反常可解释为种皮若坚硬则会阻止真菌的侵染,其种子无胚(具一团胚性细胞)无胚乳,种子萌发有赖于和特定真菌共生。
梨果果皮和种皮此消彼长现象也十分显著。苹果的内果皮是软骨质,保护功能不如骨质结构,所以其种皮也是软骨质,比膜质结构的保护功能强。
2.5 果皮特征与种子数量的相关性
桃(Prunus persica L.)、李(P.salicina Lindl.)等植物每个子房中有两个胚珠,其每个果实只有1粒种子,这是因为此果实是核果。1核果、1闭果和1分果爿都是1传播子,其趋同性状不仅是上文提到过的种皮膜质,还包括都只含1枚种子。栗属(Castanea Mill.)具坚果,其子房有9或6室,每室2胚珠,1子房共有18或12胚珠,但通常仅1胚珠发育成种子。核果、闭果和分果瓣果皮的保护使种皮失去存在的价值而退化为膜质结构,果皮对其包被种子的束缚则是无论子房含多少胚珠都只有1粒发育成种子。假如这些果实或果瓣含不止1枚种子,1个传播子内的所有种子必将被传播到一处,种子萌发后会展开激烈的同胞竞争。所以,核果内不止1胚珠时,要么进行胚胎选择,只让1胚珠发育成种子;要么像冬青属(Ilex L.)形成分核,将每个种子分装入不同的核内而不是困于1核中,使其可如浆果中的多个种子那样不一定传播到同一地点,各种子有分离的机会。可见,植物界不仅处处有种内、种间竞争,植物也会采取种种策略避免不必要的竞争。当然,核果和闭果只含1粒种子的结论也有例外,如落花生属(Arachis L.)植物果皮不开裂,虽然有些野生种只含1种子,但也有不少野生种果内有几枚种子。为什么落花生属这些野生种能忍受同胞竞争?这值得深入研究。
笔者一直对苍耳聚花果内含2瘦果倍感困惑。该聚花果如同闭果,借助总苞上的钩刺被动物传播,束缚在其内的2果被带到同一地点,势必发生激烈的同胞竞争。经过对数百粒苍耳聚花果萌发实验的观察(黎维平,未发表),发现其总是1枚种子先萌发,子叶出土,含有另一枚种子的总苞附着在子叶上,等待二次传播。于是,苍耳聚花果内二果实种内竞争的问题化解于果实的二次传播。
2.6 果实类型的分类学价值
一些果实为某一分类群特有,称为该类群的独征,也是该类群内各成员的同源共有衍征,是鉴定该类群的检索表特征。例如,瓠果、槲果、荚果、角果、颖果、梨果、蔷薇果、双悬果、柑果等分别为葫芦科、山毛榉科、豆科、十字花科、禾本科、苹果亚科(已归入桃亚科)、蔷薇属、伞形科和芸香科植物特有。当然,这些特有性状也可能有复杂变异,如豆科的荚果可演化出节荚(属分果)、瘦果等。
2.7 果实的演化
和花一样,果实是被子植物的特有性状。被子植物起源后,各类群爆发式增长,果实类型也发生多样化的演化。换句话说,果实的多样化也推动着被子植物各类群的演化[4]。所以,介绍果实系统分类,必然涉及果实多样性形成的过程,即果实的演化。
2.7.1 原始的果实类型
根据真花学说,木兰目的主要果实类型聚合蓇葖果,尤其是小果螺旋状排列的聚合蓇葖果,被认为是最原始的果实类型[6]。蓇葖果是裂果,但APGⅣ (被子植物分类系统第4版)中的被子植物基部类群却不是裂果。无油樟(Amborella trichopoda Baill.)是被子植物最基部类群无油樟目唯一现存的物种,其果实属于聚合核果或类似于聚合核果[32]。睡莲目是被子植物的第二个基部类群,其果实为坚果或浆果。被子植物的第三个基部类群是木兰藤目,包括木兰藤科、苞被木科和五味子科。木兰藤科植物的果实是聚合浆果,苞被木科的果实为浆果或浆果状,五味子科的果实为聚合浆果或小果轮生的聚合蓇葖果。所以,被子植物大多不含聚合蓇葖果,均不含小果螺旋状排列的聚合蓇葖果。但是,化石子植物辽宁古果(Archaefructus liaoningensis Sun,Dilcher,Zheng et Zhou)却有小果螺旋状排列的聚合蓇葖果[6]。不过,辽宁古果不是最古老的被子植物。聚合蓇葖果为被子植物原始果实类型的观点有待证实。
2.7.2 果实演化的几个例子
蔷薇科是植物学系统分类部分的重点内容。各教材[5~8,10~12,14~20]都按绣线菊亚科、蔷薇亚科、苹果亚科和梅亚科顺序编排,并强调4亚科在果实特征上分别对应于聚合蓇葖果、聚合瘦果和聚合核果、梨果以及核果。近年,分子系统学研究建议,蔷薇科分成3个亚科,大体上为过去的蔷薇亚科拆分为仙女木亚科和蔷薇亚科,另3个旧的亚科(绣线菊亚科、苹果亚科和梅亚科)合并成桃亚科[33]。在果实进化上,主要进化路线[33]有:1)在仙女木亚科和蔷薇亚科,聚合瘦果向聚合核果方向演化;2)在桃亚科,聚合蓇葖果分别向3个方向演化,一是心皮合生,果实演化成蒴果(如白鹃梅属Exochorda Lindl.);二是5离生心皮减少到1枚,果实发育成核果;三是5离生心皮与被丝托愈合,子房上位演化成子房半下位并进一步演化至下位,果实发育成梨果,稀为浆果状核果(如山楂属和火棘属)。至于蔷薇科的原始果实类型是聚合瘦果还是聚合蓇葖果,尚未得到可靠答案。
十字花科的姊妹科是白花菜科,白花菜科由2侧膜胎座的雌蕊发育成圆柱形蒴果;十字花科和白花菜科形成的分支的姊妹分支是山柑科,后者的果实为浆果或蒴果。所以,可以对十字花科果实的起源和演化作出推论:1)2心皮雌蕊发育成的柱状蒴果通过形成假隔膜演化成长角果;2)长角果通过减少种子、短缩果实进一步演化出短角果;3)长角果演化出横列的分果(如离子芥属的节裂角果);4)短角果演化出纵裂的分果(如臭荠属植物的果实);5)短角果演化出含1粒种子而不开裂的瘦果(见于沙芥属Pugionium Gaertn.、海滨芥属Cakil Mill.和两节荠属Crambe L.)。在果实演化上,豆科与十字花科有些相似,荚果演化出坚果、瘦果、节荚果等。
菊类植物(Asterids)与蔷薇类植物(Rosids)是APG系统真双子叶植物的主体。菊类植物的一大分支是桔梗类植物,包括冬青目、菊目、伞形目和川续断目等7个目。有研究[32]表明,桔梗类的原始果型是蒴果。冬青目、忍冬科、五加科和五福花科的肉果起源于蒴果;桔梗类的主要果实类型是含单粒种子的闭果,也是来自蒴果。除这些演化的主线外,还有多种多样的果实类型间的转化,例如:菊科的下位瘦果适应鸟播,发展出肉质果皮,转变成核果,见核果菊属(Chrysanthemoides Tourn.ex Medik.)[34]。
一些研究[35]还发现,干果中的蒴果和瘦果通常有赖于风的传播,所以干果适应于草本植物占优势的生境、温带环境或季节性干旱环境;而活跃于森林中的哺乳动物、鸟类是肉果的传播者,所以肉果被认为对热带木本被子植物多样性有重大贡献。另一些研究[35]则指出,物种多样性与种子传播综合征(dispersal syndrome)之间没有关系,推动被子植物多样性的主要力量可能是传粉和生活史多样性。
果实的演化受到多重因素的影响,且普遍存在趋同进化[4]、平行进化,果实与其传播者之间还可能存在协同进化[9],厘清多样化的果实演化还任重道远。