大花序桉半同胞家系苗期测定及家系选择
2022-11-11曹昆彬杨勇智郭洪英吴玉丹李佳蔓黄振陈炙
曹昆彬, 杨勇智, 郭洪英, 吴玉丹, 李佳蔓, 黄振, 陈炙*
1. 四川省林业科学研究院,森林和湿地生态恢复与保育四川重点实验室,四川 成都 610081;2. 四川省草原科学研究院,四川 成都 611731
大花序桉(Eucalyptus cloezianaF. Muell.)又名昆士兰桉,为桉树属昆士兰桉亚属(Idiogenes)唯一的种,高大乔木,最高可达55m[1]。大花序桉干形通直,密度高,应力变化小,是优良的硬木材,是制作高档家具材的优良材料,具有广阔的开发前景[3]。自上世纪80年代以来,南方各地开展了各类大花序桉引种试验[4]。叶露等在广西的试验结果显示,种源的树高生长以及家系的树高、胸径、单株材积生长与地点的交互作用显著,部分种源在钦州试验点表现较好,但在玉林试验点表现极差[5]。薛正华等在粤中地区的试验结果表明,5年生大花序桉的材积增长率最为显著,作为大径级工业用材培育极具潜力,但抗风能力差,适合低山丘陵少风地区种植[6]。吴坤明等在海南西部的引种试验发现大花序桉无法忍受长期干旱,试验林3年保存率仅有10%[7]。陈云峰等对桂北桉树生长与材性的研究结果表明,大花序桉适合于桂北地区发展[8]。徐耀昌在福建诏安的试验结果表明大花序桉不同家系试验林的保存率差异显著,部分家系保存率超过90%[9]。但张荣标在漳州的试验结果表明10个大花序桉家系造林平均成活率仅为74.6%[10]。杨汉波等在四川的试验结果表明大花序桉较小帽桉在生长表现上有较强的优势[11]。
开展苗期测定,在林木苗期对目标性状进行选择,是提高选育效率、加快选育过程的有效途径之一[12]。在桉树研究领域,早期选择的研究屡见不鲜。尚秀华等对1年生赤桉家系的生长性状和风害情况进行测定和遗传分析,筛选出3个具有较强抗风性的赤桉家系[13]。熊涛等分析了2.5年生赤桉第二代家系试验林的生长性状和遗传变异,筛选出5个优良家系和65株优良单株[14]。吴世军等对滇南亚高山4.5年生巨桉11种源173家系的生长性状进行分析,根据其遗传增益选择出17个优良家系[15]。玉首杰等对141个巨桉第二代家系的生长特性和遗传变异进行分析,筛选出16个优良家系和65株优良单株[16]。徐耀昌分析了闽南山地3年生大花序桉的早期生长适应性,并初步筛选出适合闽南山地造林推广的2个家系[9]。迄今为止,大花序桉种源/家系的早期选择主要对象均为大花序桉幼林[17,18],对于大花序桉家系苗期特性的研究鲜有报道[19]。因此,本试验本以47个大花序桉家系为试验材料,对其苗期生长性状进行研究,探究其遗传变异规律,并开展早期选择,选择出表现良好的家系补充进四川大花序桉基因库,丰富四川大花序桉的选育基础。
1 材料和方法
1.1 材料来源
本试验参试的47个家系均来自澳大利亚北昆士兰州,属热带气候,年平均降水量可达4 500 mm,年均最高气温45℃,最低气温-5℃。夏季炎热多雨,温度在20℃至30℃之间;冬季气候温和晴朗,最高温度在10℃左右。参试家系种子由澳大利亚CSIRO种子中心提供。各种源、家系基本情况详见表1。

表 1 大花序桉种源、家系基本情况Tab. 1 Basic information on provenance and family of Eucalyptus cloeziana
1.2 试验地点
育苗地点设置在成都市郫都区横山村。属亚热带季风性湿润气候,夏无酷暑,冬无严寒,雨量充沛。年平均气温16℃,一月平均气温5℃,八月平均气温26℃左右。年度极端最高气温35.3℃、极端最低气温-4.0℃,降水量979.4 mm,日照1 014.0 h。
1.3 试验方法
整地要求深翻20 cm以上,整平耙细;畦宽1 m、长15~20 m。 播种前对大花序桉种子消毒,消毒液为1 000倍高锰酸钾溶液,直接把带有大花序桉种子的网袋完全浸泡在消毒液中,浸泡时间为30 min,浸泡后用自来水冲洗至无色为止,并晾干。把种子袋重新并置于恒温恒湿的环境中催芽至种子露白。
试验采用完全随机区组设计,3次重复。2020年3月底,以株行距为20 cm×20 cm的规格进行点播,播种后覆盖0~4 mm厚泥炭土,使肉眼不见种子即可。播种后,用细蓬头浇透苗床,并搭建小拱棚。待首批幼苗出土后,掀开拱棚两侧的薄膜,开始通风,棚内温度不高于25℃。待苗高5 cm时,着重注意防范猝倒病,可用1 000倍喹啉酮溶液喷洒,每隔15 d喷洒1次至苗高10 cm以上、茎段开始木质化。
待幼苗长至高5~10 cm,可用小铲子从苗床下10 cm处水平挖入,小心端出幼苗,注意不能伤到根部。预选准备直径4.5 cm、高8 cm的基质段,基质段成分为0~10 mm泥炭土:椰糠=1∶1,用筷子或直镊在基质段中间捅一深洞后,弄镊子夹住根系前段,把幼苗整体送入基质段。
移栽后第3 d、10 d、17 d、24 d施用3‰复合肥(氯化铵型复合肥N15-P5-K5);后期追肥指移栽一个月后的追肥,肥料使用氯化铵型复合肥(N15-P5-K5)。追肥均采用复合肥溶液淋洒,配制成相应浓度后,单次施肥量按每840株施用10 L溶液计算。
在每个家系中随机选取10株苗木,重复3次,用钢尺(1 m)和游标卡尺分别测量苗木的苗高和地径(距地面大约1 cm处的苗木直径)。
1.4 数据分析
采用SPSS 20.0和DPS 16.05高级版进行数理统计分析。
遗传参数采用以下公式计算:

其中:为单株遗传力;为家系遗传力;分别为家系、环境的方差分量。n1和n2分别表示重复数和小区调和株数。CVG和CVP分别表示遗传变异系数和表型变异系数;表示表型方差。ΔG表示遗传增益,X表示性状总平均值,x表示入选家系性状均值。
2 结果与分析
2.1 大花序桉家系苗期生长比较
大花序桉家系苗期苗高和地径方差分析结果见表2,结果表明,2个性状的生长差异在种源和家系间均达到极显著差异水平,这表明大花序桉幼苗在种源和家系间存在丰富的变异,具有较大的家系选择潜力,这为苗期初步选择优良家系提供了依据。
苗高和地径多重比较结果见表3,47个大花序桉家系平均苗高35.03 cm,苗高超过均值的家系有23个,2号家系苗高最高,为56.68 cm。47个家系苗高整体变异系数为35.66%,家系苗高变异系数高于整体苗高变异系数的家系分别为10号、17号、20号和22号家系,说明这4个家系内的苗高变异较大,在育种实践过程中存在提供较高的种质变异和创新贡献率的可能。47个家系根据苗高可分为19个子集,表现最好的子集记为a,仅由2号家系组成,苗高变异系数为19.92%,远低于整体苗高变异系数,说明2号家系苗高性状较为稳定。
47个大花序桉家系平均地径为5.68 mm,地径超过均值的家系有24个,15号家系的地径最大,为8.82 mm。47个家系地径整体变异系数为47.39%,表明地径这个性状存在丰富的变异;家系地径变异系数超过整体地径变异系数的家系分别为6号、17号、20号和30号家系,其中6号家系的地径变异系数达到98.47%,说明该家系地径性状的表型极其不稳定。47个家系根据地径可分为18个子集,表现最好的子集(标记a)由6个家系组成,分别为15号、6号、2号、5号、1号和7号家系,除6号家系外,其余5个家系地径变异系数介于27.40%~35.98%之间,低于地径整体变异系数,说明这5个家系的地径性状较为稳定,但也能够提供较高的种质变异。
2.2 大花序桉家系苗高和地径的相关分析
对大花序桉47个半同胞家系的苗高和地径性状相关性分析结果表明(见表4),大花序桉苗期苗高和地径性状呈极显著正相关,相关系数为0.354。因此,在苗期进行大范围选优时,可以将过程简化为根据苗高进行筛选。
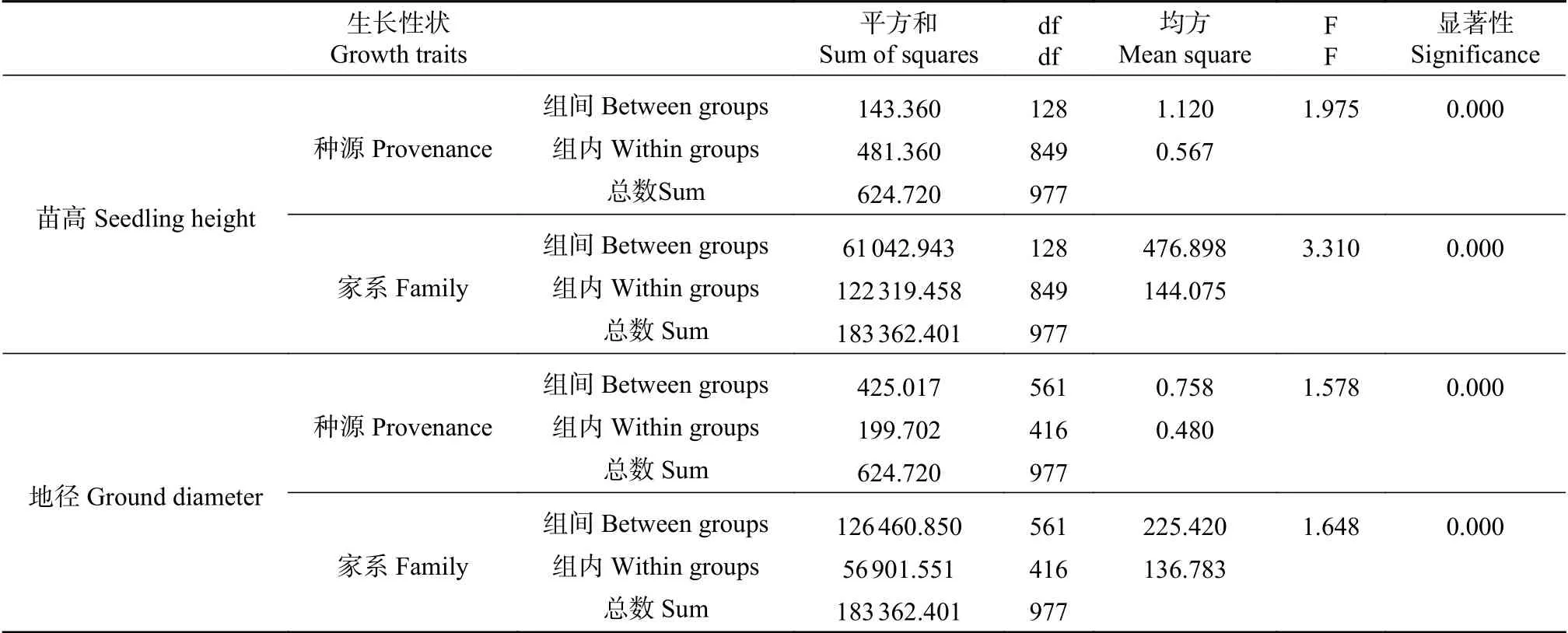
表 2 大花序桉半同胞家系苗期生长性状方差分析Tab. 2 Variance analysis of seedling growth traits for Eucalyptus grandiflorum half-sib families
2.3 大花序桉家系苗高和地径的遗传变异
大花序桉家系苗高和地径的遗传参数见表5,结果表明,大花序桉家系苗高和地径的家系遗传力分别为0.97、0.95,属于强度遗传范围,这表明苗高和地径这两个性状生长受遗传因素影响大。能够保障入选家系优良性状的高度遗传,确保选育结果能获得更好的遗传增益。苗高和地径遗传变异系数为28.61%、28.18%,处于较高水平,表明这47个大花序桉半同胞家系在苗期存在丰富的遗传变异,选育基础良好,遗传改良潜力较大,可以为大花序桉的苗期选育提供丰富的选育材料。
表型变异系数是表征环境方差占总均方的比例,大花序桉家系苗高和地径的表型变异系数分别为3.17%、4.57%,均不足5%,说明家系苗的生长受环境的影响较小。参试幼苗苗龄仅有15个月,且整个生长过程均由人为管控、未受到逆境影响,因此环境对苗高和地径这两个性状的影响较小。
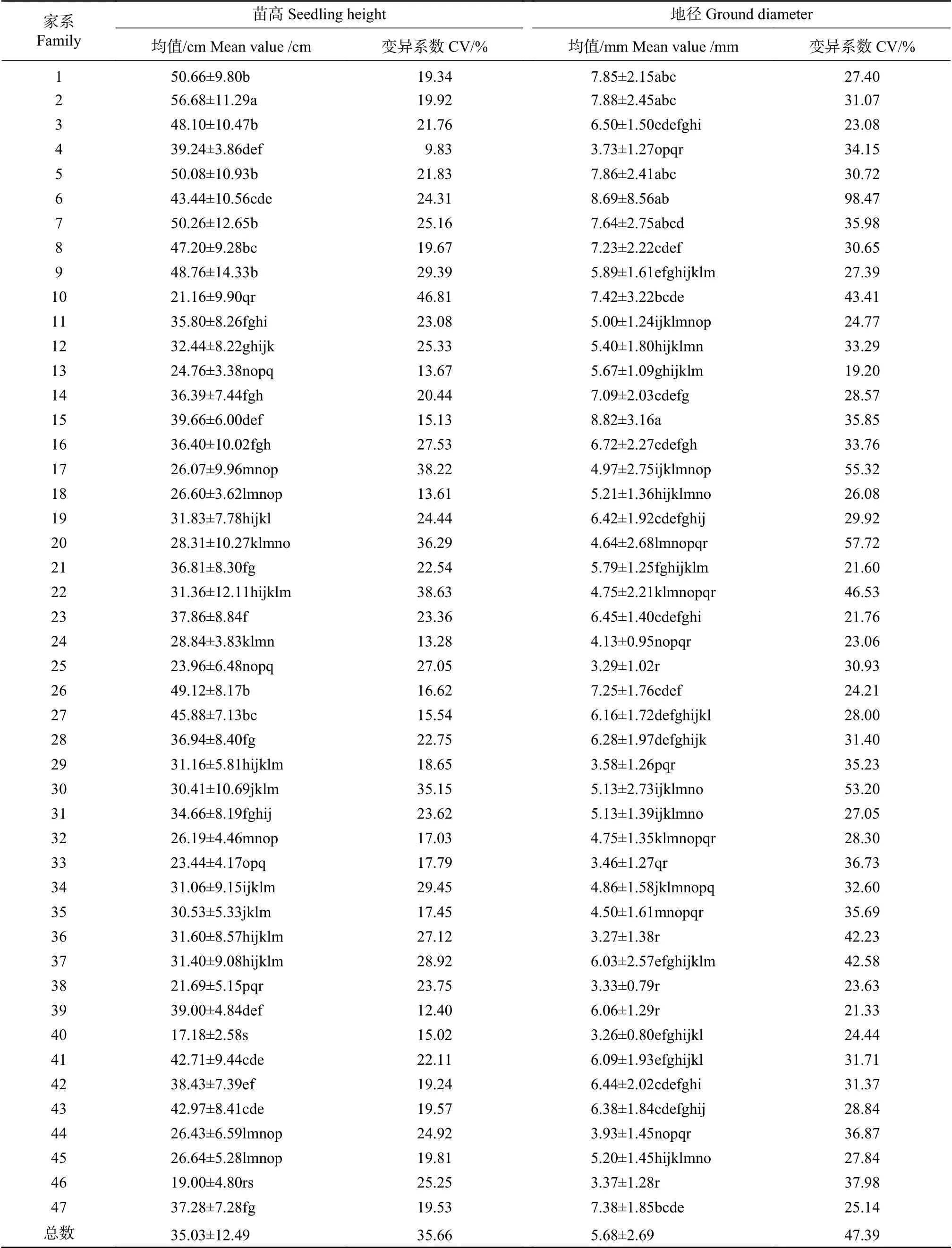
表 3 大花序桉家系苗高和地径生长比较Tab. 3 Comparison of seedling height and ground diameter taits of Eucalyptus cloeziana families

表 4 大花序桉家系苗高和地径相关性分析Tab. 4 Correlation analysis of seedling height and ground diameter characters of Eucalyptus cloeziana half-sib families

表 5 大花序桉家系苗高和地径遗传参数Tab. 5 Genetic parameter of seedling height and ground diameter taits of Eucalyptus cloeziana half-sib families
2.4 优良大花序桉家系选择
比较各家系性状生长与苗期总体平均值,超出苗高总体平均值的家系共有23个家系,超出地径总体平均值的家系有24个。将超出性状苗期总体平均值30%的家系作为优良家系的候选家系,苗高超出总体均值30%的家系有9个,分别为2号、1号、7号、5号、27号、9号、3号、8号和28号家系,2号家系苗高超出总体均值61.81%;地径超过总体平均值30%的家系有7个,15号、6号、2号、5号、1号、7号和10号家系,15号和6号家系,其中15号和6号家系地径分别超出总体平均值55.28%和53.12%。
计算上述各候选优良家系性状的遗传增益(见表6)。结果表明,2号家系苗高遗传增益最高,达到59.95%,15号家系地径遗传增益最好,达到53.63%。苗高和地径的遗传增益超过30%的家系有4个,分别为1号、2号、5号和7号家系,苗高和地径表现在所有候选优良家系中均较好,可入选优良家系。余下的候选优良家系均存在在某个性状的遗传增益表现较好,但另一个性状的遗传增益表现一般的情况,如15号和6号家系,地径遗传增益达到53.63%和51.52%,但其苗高遗传增益仅为12.82%和23.29%,综合表现一般,因此需要在后续的培育中进一步观察这些家系的表现。
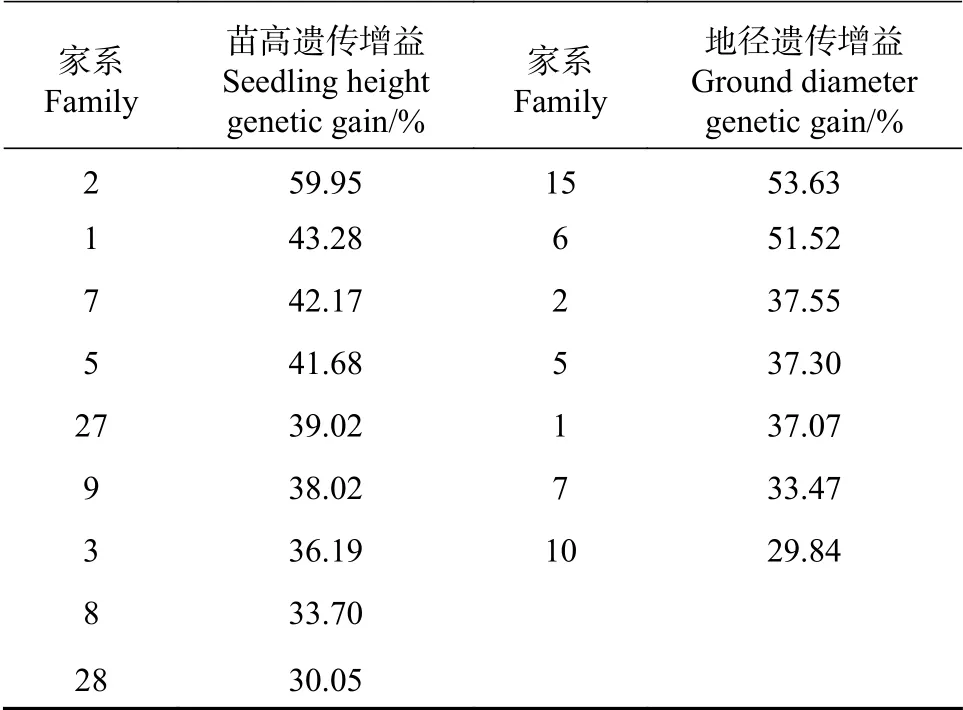
表 6 大花序桉候选优良家系遗传增益Tab. 6 Genetic gain of candidate superior Eucalyptus cloeziana families
3 结论与讨论
遗传变异是选择的基础,对变异来源、特点和规律的研究是进行树种改良并获得遗传增益的理论前提[20]。本试验结果表明,大花序桉苗高和地径在家系间差异极显著,说明这2个性状在家系水平上具有较大的遗传改良潜力,这与陈小中等的研究结果一致[19]。苗高和地径的变异系数分别为35.66%和47.39%,低于陈小中等对引种大花序桉优树子代研究结果[19],表明引种驯化后的大花序桉子代存在更大的变异。
遗传力是估算遗传增益的重要参数,是亲本将其遗传特性传递给子代能力大小的体现[21]。本试验中,苗高和地径的家系遗传力均高于不同树龄的大花序桉种源/家系[22]、1年生赤桉家系[13]、6年生大花序桉种源[11]、4.5年生滇南亚高山巨桉种源/家系[15],两个性状的家系遗传力均高于0.9,受到高等强度遗传控制,说明所选家系的苗高和地径能够稳定遗传。
遗传变异系数表征遗传因素导致性状观测值相对变异程度,其数值大小可体现该性状遗传改良潜力的大小[23]。本试验苗高的遗传变异系数高于1年生赤桉家系[13]、滇南亚高山巨桉种源/家系[15]的研究结果,地径的变异系数高于1年生赤桉家系[13],较4.5年生滇南亚高山巨桉种源/家系[15]略低,表明大花序桉苗高和地径具有较大的选择潜力。表型变异系数体现该性状表型受环境影响的大小,本试验中两个性状的表型变异系数均小于滇南亚高山巨桉种源/家系[15],这与参试苗木的年龄有关,本试验参试苗龄仅为15个月,因此受到环境影响的加性效应较小。
大花序桉是极具培育价值的珍贵用材树种,本试验初步筛选出2号、1号、5号和7号4个优良家系,苗高遗传增益均达到40%以上,地径遗传增益均达到30%以上,高于桢楠、红松和杉木等用材树种[24-26],说明初选的优良家系具有较高的遗传增益,通过苗期选择的方式进行遗传改良将会取得较好的效果。但家系选择势必要淘汰部分家系,导致群体遗传基础变窄,在多世代育种过程中会加速近交率的增加,这些情况广泛存在于杉木、辐射松、马尾松等传统用材林树种育种进程中[27-29]。本试验引种的47个家系苗高和地径在种源和家系间存在丰富的变异,且受高度遗传控制,具有良好的遗传改良潜力。以此补充四川大花序桉基因库,能够增加四川大花序桉的遗传多样性,丰富四川大花序桉的选育基础,降低近交率的增加速度。
大花序桉与四川传统乡土珍贵用材树种桢楠一样,常作为大径材培育。本试验初步筛选出2号1号、5号和7号为优良家系,仅为大花序桉优良家系早期选育提供参考,但优良家系的适应性和稳定性仍需通过对后续区域造林试验的跟踪观测来进行分析验证。此外,也可参考桢楠等乡土树种开展林下种植[30]等相关方向研究,为大花序桉的开发利用提供参考。
