生物炭-过氧化钙复合颗粒对酚酸胁迫下番茄生长和根际微生态的调控效应
2022-11-10涂玉婷彭智平许杨贵李珠娴黄继川
涂玉婷,彭智平,许杨贵,李珠娴,黄继川
(广东省农业科学院农业资源与环境研究所 / 广东省养分资源循环利用与耕地保育重点实验室 /农业农村部南方植物营养与肥料重点实验室,广东广州 510640)
近年来,我国农业产业结构调整推动着番茄种植面积大幅增长。为了获得更高的经济效益,番茄周年连茬种植十分普遍[1]。但番茄是不耐连作的作物,这种高强度连续种植的生产模式往往造成土壤自然生态调节功能衰退,植株生长发育受限,并导致番茄产量大幅下降,减产率甚至高达70%[2-3]。解决连作障碍问题对保障番茄产业可持续发展具有重要意义[4]。在连茬种植条件下,土壤中累积的化感物质尤其是酚酸类化感物质,对植物生理的抑制作用及对根际微生态环境恶化的诱导效应是导致连作障碍的主导因素[5-6]。因此,开发高效去除土壤中累积的化感自毒物质并改善土壤微生物群落结构和功能的技术,有望从根源上缓解连作障碍问题。
生物炭是一类以生物质为原料、利用热解工艺制备的多功能材料,具有孔隙丰富、吸附性强等优异特性[7]。近年来,研究发现施用生物炭对连作土壤改良及农作物增产提质均具有一定的效果。Zhang等[8]研究发现,施用1%的棉花秸秆生物炭可通过改善棉花根系生长、增加根系活力、优化土壤养分状况来缓解棉田连作障碍问题。Wu等[9]研究表明,在连续单作土壤中添加稻壳生物炭可以调节黄芪根际微生物群落结构、丰度和代谢,并减少病原体的数量。此外,生物炭对包括酚酸在内的化感物质具有良好的吸附性能,能够有效降低土壤团粒中化感自毒物质的含量[10]。已有研究表明,生物炭对特定有机物的吸附受生物炭原料、热解温度、比表面积、孔隙性质、芳香度、表面官能团、表面碱度等多种因素的影响[11-12]。生物炭对酚酸的吸附量随着热解温度的升高总体上呈增加趋势。对于高温热解制备所得生物炭,其比表面积是影响酚酸吸附量的主导因素[13-14]。因此,本研究选用比表面积丰富的椰壳活性炭用于生物炭基连作土壤改良剂的制备。然而,研究发现富集于生物炭中的高浓度化感物质对植物根系正常生长存在抑制效应,并且在特定条件下会经解吸附作用再次释放回土壤中,对作物和土壤微生物的生长代谢产生潜在威胁[15-16]。
过氧化钙、过氧化镁、过硫酸铵等无机氧化剂具有较高的热稳定性和反应过程可控性,常被用于土壤原位修复、稻田地力提升、食品和医疗等领域[17-19]。这类过氧化物在催化条件下可分解产生羟基自由基、超氧自由基等强氧化性自由基,从而有效去除土壤中难降解有机物,如石油醚、多环芳烃、多氯联苯等[20-21]。黄君霞等[22]研究发现,施用过硫酸铵可一定程度优化连作苹果园土壤环境。据此,本研究提出将生物炭与无机氧化剂相耦合,借助“生物炭吸附-过氧化物氧化”的协同作用,实现对土壤中酚酸类化感物质的有效去除和土壤微生态的正向调控。
本研究选取生产中广泛使用、药源丰富且对环境无污染的过氧化钙为氧化剂,通过造粒技术制备生物炭-过氧化钙复合土壤调理剂。借助番茄盆栽试验,研究酚酸胁迫下番茄植株生长与生理指标、果实产量和品质对生物炭-过氧化钙复合颗粒输入的响应,分析施用复合颗粒对番茄根际土壤理化性质及酚酸残留量、细菌和真菌群落丰度的影响。通过关联分析,揭示复合颗粒对番茄生长和根际土壤微生态调控效应,为连作障碍土壤改良提供理论依据和方法参考。
1 材料与方法
1.1 试验材料
试验所用番茄品种为千禧圣女果,是广东地区主要栽种的小番茄品种之一。供试酚酸为对香豆酸、邻苯二甲酸、苯甲酸、对羟基苯甲酸,均由阿拉丁试剂有限公司生产。盆栽试验用土采自水稻田,为赤红壤,pH为6.15,电导体(EC)值为0.26 mS/cm,有机质含量为2.12%,碱解氮、有效磷和速效钾含量分别为91.3、82.1和56.4 mg/kg。所用生物炭为广州化学试剂厂生产的椰壳活性炭。过氧化钙购自上海麦克林生化科技有限公司。生物炭-过氧化钙复合颗粒参照中国发明专利ZL 202010841842.5[23]所介绍的方法进行制备。生物炭与过氧化钙的质量比为4∶1,矿物粘结剂添加量为4%。制备时先使过氧化钙与矿物粘结剂混合粉末通过造粒机制成直径约2.0 mm的过氧化钙球形内核,进一步使用圆盘造粒技术制备得粒径约为5 mm的生物炭包覆过氧化钙颗粒。所制得的生物炭-过氧化钙复合颗粒比表面积为475.19 m2/g,最大变形力为30.12 N。同时,利用圆盘造粒机按相同配比分别制备生物炭和过氧化钙颗粒,用于对比试验。
1.2 试验方法
番茄盆栽试验于2020年9月—2021年3月在广东省农业科学院农业资源与环境研究所遮阴网室进行。试验共设置5个处理,分别为:1)常规栽培(CK);2)酚酸胁迫 (T1);3)酚酸胁迫+30 g/kg生物炭-过氧化钙复合颗粒处理 (T2);4)酚酸胁迫 + 24 g/kg生物炭颗粒 (T3);5)酚酸胁迫+6 g/kg过氧化钙颗粒 (T4)。其中CK、T1、T3处理通过补充适量硫酸钙,保证各处理中钙元素添加量一致。
研究前期使用HPLC-MS测定连作两茬后番茄根际土中酚酸种类和含量,共鉴定出对香豆酸、苯甲酸、对羟基苯甲酸、邻苯二甲酸、香草酸、阿魏酸、肉桂酸等11种酚酸,其中前4种酚酸占已检出酚酸总量71.5%~76.6%,也是广泛报道的导致连作障碍的酚酸类化感物质[24-25]。前期测定结果还表明,番茄根际土中对香豆酸、苯甲酸、对羟基苯甲酸、邻苯二甲酸的含量比约为3∶1∶0.8∶0.8。且研究表明,向土壤中添加100 μg/g以上的外源酚酸,会对作物的生长和土壤微生物区系产生化感效应[26-27]。综合考虑酚酸在土壤中的吸附与转化,本研究外源酚酸的总施用量为140 μg/g,包含75 μg/g对香豆酸、25 μg/g 邻苯二甲酸、20 μg/g 苯甲酸、20 μg/g 对羟基苯甲酸。所有处理分别将外源添加物、基肥和酚酸溶液与已过2 mm筛的6.5 kg风干土样混匀后,装入高24.0 cm、上口径26.0 cm、底径18.0 cm的塑料盆中。每钵移栽1株高约20 cm的番茄幼苗,每个处理5次重复,每个重复3株苗,共75盆番茄苗,随机排列。试验期间所有处理水肥管理一致。
1.3 测定项目与方法
在移栽后30和120天,每个处理随机选取5株番茄苗,测定地上部及地下部鲜重,采用氯化三苯基四氮唑法测定根系活力。同时,采用抖土法收集移栽后120天 (采果盛期)的番茄植株根际土,一部分土样经风干、过1 mm筛,参照《土壤农业化学分析方法》[28]测定土壤pH、EC值、有机质、碱解氮、有效磷、速效钾含量等土壤理化指标;另一部分土样存于-80℃超低温冰箱中,用于酚酸含量和微生物学测定。土壤中残余酚酸采用李培栋等[29]的方法进行提取,采用福林酚比色法测定土壤中总酚酸的残余量。土壤总DNA使用试剂盒进行提取,本研究使用在生态学研究中被广泛使用的引物对3 3 8 F/806R和ITS1F/ITS2R分别对细菌16S rRNA基因的V3~V4区和真菌ITS1区段进行PCR扩增[30-32],经纯化、定量、均一化处理后建立测序文库,在此基础上,采用基于Illumina HiSeq平台完成测序。对所得的Raw Reads数据进一步进行过滤、拼接、OTUs聚类、物种注释及相对丰度分析等操作。
番茄单株果实产量采用称重累计法计算。各处理随机选择5株番茄的第2、3穗果实用于平均单果重和品质指标的测定。分别采用2,6-二氯酚靛酚滴定法、蒽酮比色法、酸碱滴定法测定番茄果实维生素C、可溶性糖、有机酸含量。
1.4 数据分析
采用Excel 2016、SPSS 19.0进行试验数据处理及统计分析,并通过Duncan’ s多重比较法进行差异显著性检验。微生物多样性数据使用QIIME和mothur软件进行分析。冗余分析 (redundancy analysis,RDA)用Canoco5.0软件完成。使用Excel 2016和R语言绘图。
2 结果与分析
2.1 不同处理对番茄植株生长和根系活力的影响
如表1所示,与CK处理相比,酚酸胁迫处理(T1)导致移栽后30和120天的番茄苗生物量、地下部鲜重和根冠比均显著下降,降幅为12.1%~35.2%,植株根系活力分别下降了38.9%与44.0%。可见,外源酚酸明显抑制了番茄植株的生长势和生理活性。施用复合颗粒处理 (T2)、生物炭颗粒处理(T3)和过氧化钙颗粒处理 (T4)的番茄生物量、地下部鲜重、根系活力均高于同时期的T1处理,显示出一定程度的抗酚酸胁迫效果,其中以T2处理的效果最明显。与T1处理相比,T2处理番茄苗移栽后30和120天的生物量和地下部鲜重增幅为15.9%~39.6%,根冠比分别增加了22.2%和4.7%,根系活力平均提高了47.2%。与T3和T4处理相比,T2处理的地下部鲜重增加了7.0%~25.4%,根系活力提高了14.1%~22.9%。由此可见,生物炭-过氧化钙复合土壤调理剂可以缓解酚酸对番茄苗生长的胁迫。
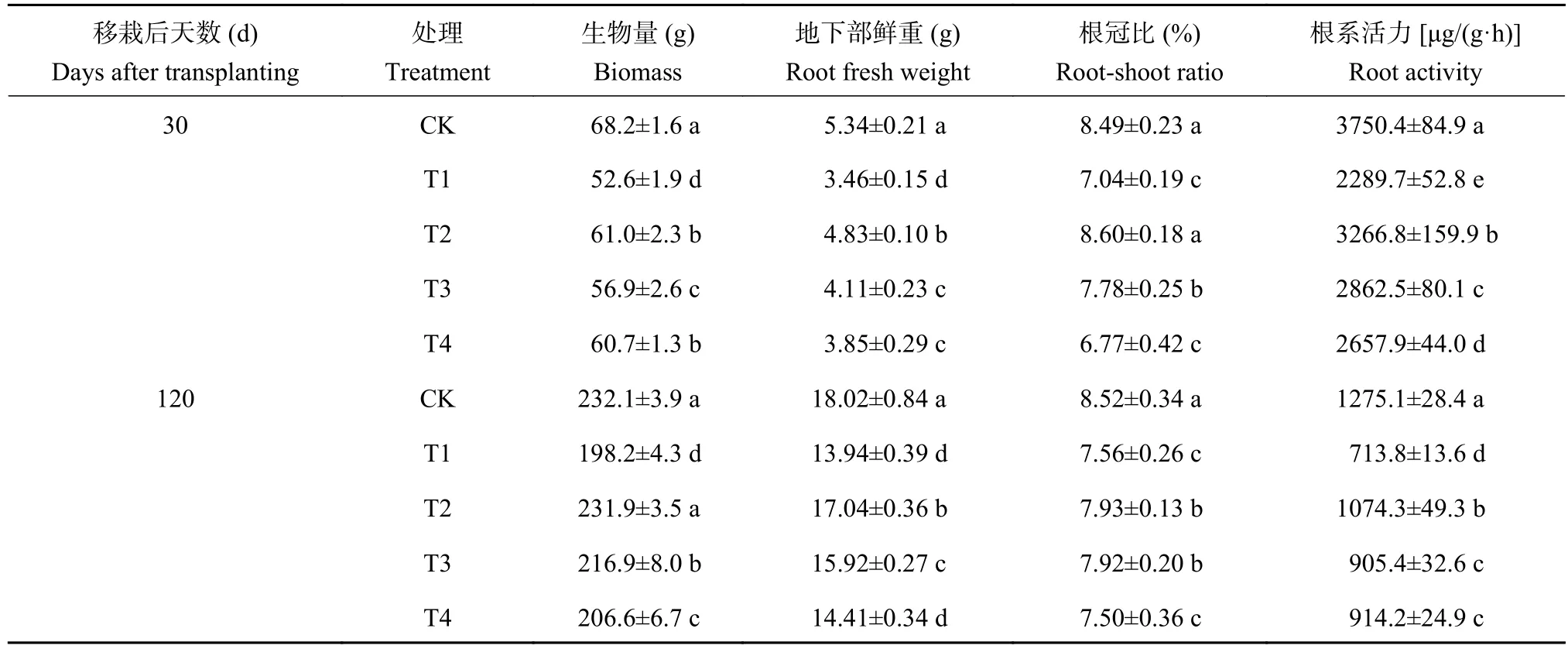
表1 不同处理番茄苗的生长指标和根系活力Table 1 Growth index and root activity of tomato plants under different treatments
2.2 不同处理对番茄果实产量和品质的影响
如表2所示,5个处理番茄果实单株产量由大到小依次为T2、T3、CK、T4、T1。与CK处理相比,T1处理的单株产量、单果重分别下降9.2%与16.8%,有机酸含量提高29.7%,维生素C和可溶性糖含量、糖酸比分别下降了13.9%、10.7%及30.8%。施用复合颗粒 (T2)和生物炭 (T3)均能显著缓解酚酸胁迫对番茄产量形成的负效应。其中,施用复合颗粒的增产效果最明显,单株产量和单果重分别比T1处理提升了13.8%和20.1%,且果实Vc、可溶性糖含量均显著提高,糖酸比增加52.6%。T2处理的番茄果实单果重、Vc和可溶性糖含量与CK处理相比差异不显著,且T2处理的单株产量和糖酸比显著高于CK处理,增幅分别为3.4%和5.5%。可见施用复合颗粒可有效促进酚酸胁迫下番茄的增产提质。
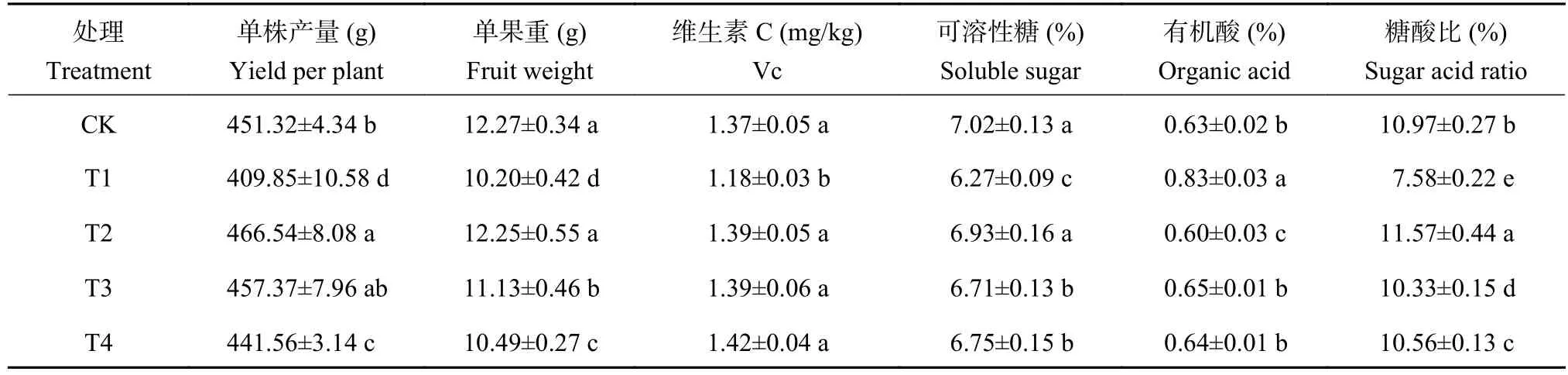
表2 不同处理的番茄果实产量和品质Table 2 Tomato yield and quality under different treatments
2.3 不同处理对番茄根际土壤理化性质的影响
由表3可以看出,T1处理的番茄根际土中残留的酚酸总量比CK处理高出36.5%。但与T1处理相比,添加复合颗粒 (T2)、生物炭 (T3)、过氧化钙(T4)处理根际土壤中酚酸残余量均显著降低,降幅达41.1%~44.6%,以T2处理的去除效果最佳,其根际土壤残余酚酸量显著低于T3和T4处理。与CK相比,T1处理的土壤pH下降了0.17个单位,有机质、碱解氮和速效钾含量分别降低了3.7%、0.9%和12.3%,EC值显著增加了10.9% (P<0.05)。与T1处理相比,T2处理的番茄根际土壤EC值下降17.7%,pH提高了0.77个单位,有机质和碱解氮含量分别增加77.4%和2.8%。此外,与CK相比,T2处理的土壤pH提高了0.60个单位,土壤酚酸含量和EC值分别降低了24.3%和8.7%,有机质和碱解氮含量分别提升70.7%和1.9%。可见,施用生物炭-过氧化钙复合颗粒能够有效降低土壤中残余酚酸含量和EC值,提高土壤pH和有机质含量。
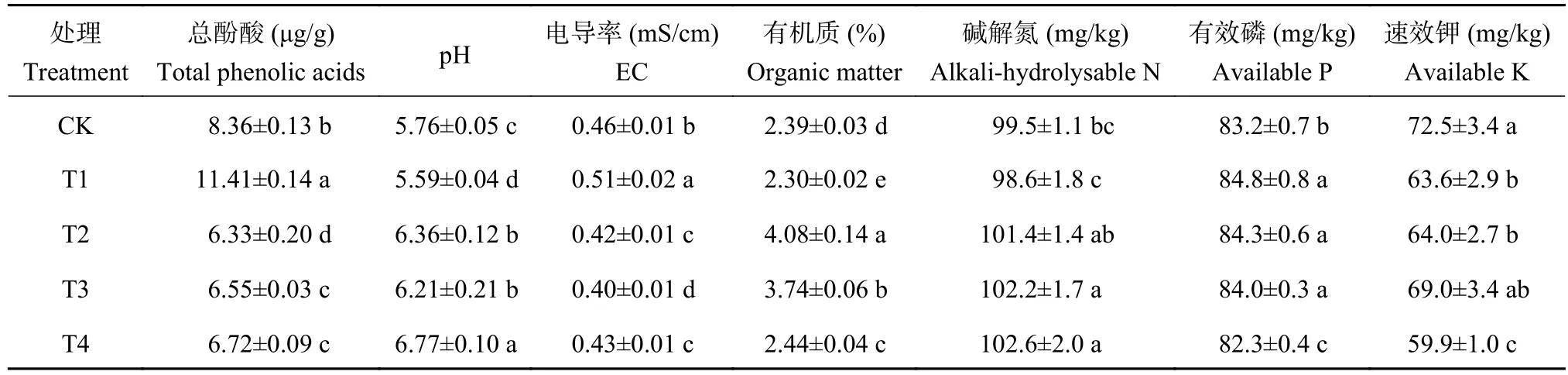
表3 不同处理番茄采收期根际土壤理化指标Table 3 Physicochemical indxes of rhizosphere soil in tomato harvesting period under different treatments
2.4 不同处理对根际土壤微生物多样性的影响
如图1a和图1b所示,与CK相比,酚酸胁迫下(T1)与土壤样本细菌群落丰富度相关的ACE指数降低4.8%,反映物种丰富度和均匀度的香农指数增加了5.6%。说明添加外源酚酸会降低根际土壤细菌群落丰富度,略微提高均匀度。与T1处理相比,施用复合颗粒、生物炭及过氧化钙颗粒的T2~T4处理,均显著提高番茄根际土细菌群落的ACE和香农指数。其中T2处理的增幅最大,ACE和香农指数分别增加了19.5%与13.9%。图1a中T1处理的番茄根际土中真菌ACE指数与CK相比提高了23.7%,差异达显著水平;图1b中真菌香农指数差异未达显著水平。同T1处理相比,施用复合颗粒 (T2)处理的根际土壤真菌群落ACE和香农指数分别下降13.2%和16.7%,且差异显著。而T2处理土壤样品细菌和真菌的ACE指数与CK相比分别增加13.7%和7.4%,细菌的香农指数提高了20.6%,真菌的香农指数下降了18.8%。

图1 不同处理根际土壤中细菌和真菌的ACE指数(a)、香农指数(b),真菌与细菌OUT比值(c)Fig.1 ACE(a) and Shannon index (b) of bacteria and fungus in rhizosphere soil, and the ITS1/16S observed species ratio (c) of different treatments
本研究使用各处理番茄根际土壤样品中真菌与细菌OUT数的比值,来衡量不同处理对根际土壤细菌和真菌群落之间均衡性的影响[33],结果如图1c所示。酚酸胁迫的T1处理的ITS1/16S OTU比值显著高于CK,表明外源酚酸胁迫对土壤中细菌与真菌之间的平衡状态产生影响,驱动土壤微生物区系向“真菌型”演变。而与T1处理相比,T2~T4处理均能显著降低酚酸胁迫下土壤ITS1/16S的OTU比值,且添加复合颗粒 (T2)处理具有最低的比值。以上结果表明,施用复合颗粒能够有效缓解外源酚酸对土壤微生物的趋化作用,增加根际土中细菌群落多样性,降低真菌多样性,优化土壤微生物群落结构,驱动番茄根际土壤微生物区系向“细菌型”转变。
2.5 不同处理对根际土壤细菌和真菌群落结构的影响
图2a为5个处理的番茄根际土壤中门水平上的细菌群落组成,各处理根际土壤中平均相对丰度>2%的细菌类群包括变形菌门 (相对丰度为33.8%~44.2%)、酸杆菌门 (12.%~25.6%)、放线菌门(8.6%~16.7%)、绿弯菌门 (5.7%~12.0%)、芽单胞菌门 (7.8%~11.4%)、拟杆菌门 (1.7%~5.6%)。这6类细菌占全部细菌群落的89.4%~94.8%。酚酸胁迫(T1)处理中酸杆菌门的相对丰度显著高于CK,并导致其余5类细菌群落的相对丰度不同程度地下降,且差异均达显著水平 (P<0.05)。施用复合颗粒(T2)和过氧化钙颗粒 (T4)处理可显著降低酸杆菌门的相对丰度,而施用生物炭颗粒 (T3)处理对酸杆菌门丰度的影响未达显著水平。此外,T2~T4处理均可不同程度地提高酚酸胁迫下绿弯菌门和拟杆菌门的相对丰度。
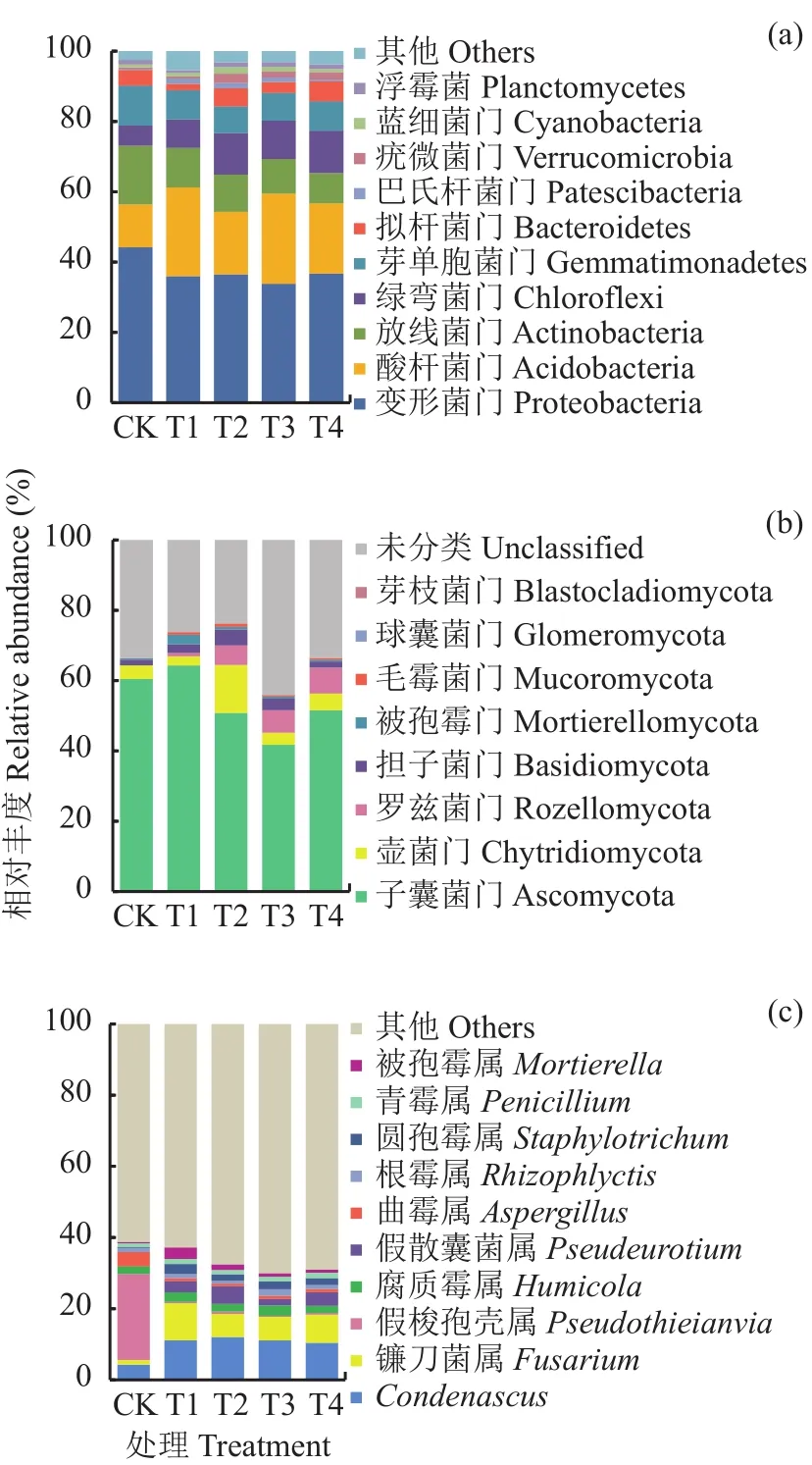
图2 不同处理番茄根际土壤细菌门水平(a)、真菌门水平(b)和属水平(c)群落结构Fig.2 Community structure of bacterial microbiota at the phylum level (a), fungal microbiota at the phylum and genus levels (b , c) under different treatments
从图2b中可以看出,5个处理的番茄根际土壤在门水平上平均相对丰度>1%的真菌菌群包括子囊菌门 (41.7%~64.3%)、壶菌门 (2.6%~1.36%)、罗兹菌门 (0.02%~6.4%)、担子菌门 (1.5%~4.6%),被孢霉门 (0.2~3.1%),这5类真菌占到已分类菌群的98.3%~99.9%。与CK处理相比,酚酸胁迫 (T1)处理导致子囊菌门、担子菌门和被孢霉门的相对丰度呈现不同程度地增加。施用固体调理剂的3个处理与T1处理相比,根际土壤在真菌门水平上的菌群丰度发生明显变化,其中子囊菌门和被孢霉门丰度均显著下降,罗兹菌门丰度显著上升 (P<0.05)。
从图2c可以看出,与CK相比,酚酸胁迫(T1)处理导致番茄根际土壤中Condenascus、镰孢菌属、假散囊菌属的相对丰度显著增加 (P<0.05),施用3种固体调理剂的处理中,镰孢菌属、假散囊菌属的相对丰度均出现不同程度地下降,其中施用生物炭-过氧化钙复合颗粒处理 (T2)植株和生物炭颗粒处理(T3)中,镰孢菌属的相对丰度与T1处理相比分别显著下降了38.1%和36.6% (P<0.05)。
2.6 番茄产量、品质与土壤环境因子及微生物群落丰度的关联分析
将番茄产量、糖酸比、土壤环境因子、平均相对丰度大于2%的细菌和大于1%的真菌群落丰度进行Spearman相关性分析(图3),发现本研究中番茄根际土壤pH、有机质 (OM)与土壤总酚酸 (TPhA)、电导率(EC)值均呈极显著负相关。pH和OM与细菌中的绿弯菌门相对丰度呈极显著正相关关系,与拟杆菌门相对丰度呈显著正相关,与芽单胞菌门相对丰度呈负相关;与真菌中的罗兹菌门相对丰度呈极显著正相关,与壶菌门相对丰度呈显著正相关;与子囊菌门相对丰度呈极显著负相关。而TPhA和EC与以上菌群丰度的相关性正好相反。由此可见,土壤pH、OM与TPhA、EC对土壤中主要微生物的丰度具有相反的调控作用。图3还表明,番茄产量与土壤OM、壶菌门相对丰度极显著正相关,与土壤TPhA和EC值呈极显著的负相关,并且与酸杆菌门和子囊菌门相对丰度也有非常显著的负相关关系;体现番茄品质的糖酸比 (SAR)也与土壤OM、壶菌门、拟杆菌门相对丰度呈极显著正相关,与土壤pH呈显著正相关,与TPhA和酸杆菌门相对丰度也呈显著负相关。

图3 番茄产量、糖酸比与土壤理化性质、优势菌群相对丰度之间的相关性热图Fig.3 Correlations between tomato yield, sugar-acid ratio, soil physicochemical properties, and relative abundance of dominant bacterial and fungi communities
采用RDA冗余分析,探讨各处理根际土壤环境因子与土壤主要微生物群落丰度之间的相关关系(图4)。RDA分析结果中第1、第2排序轴的解释度分别为47.6%和31.6%,前两个轴累积解释了群落变异程度的79.2%,土壤环境因子中TphA对群落变异的解释度为38.5% (P=0.002),pH的解释度为27.9%(P=0.002),OM的解释度为14.8% (P=0.008),EC的解释度为12.0% (P=0.004),因此,TPhA、pH、OM、EC是驱动群落变异的主要土壤环境因子。RDA分析所得结果与上述Spearman相关性热图分析结果一致。
从图4可以看出,各处理组样本点能够获得良好地分离,说明酚酸胁迫及3种调理剂添加均对土壤微生物群落结构产生显著影响。施用外源酚酸(T1)处理与土壤TPhA和EC值呈正相关关系,且与子囊菌门、被孢霉门、酸杆菌门丰度也呈现正相关。施用生物炭-过氧化钙复合颗粒的T2处理样本点与pH和OM呈显著正相关,与TPhA、EC呈显著负相关,且与壶菌门和拟杆菌门的丰度呈正相关关系,而这两类微生物与番茄产量和糖酸比也显著正相关。此外,T2处理样本点与番茄枯萎病相关的子囊菌门的丰度存在负相关关系。可见,施用复合颗粒的T2处理与施用酚酸的T1处理对与番茄产量和品质相关的关键土壤环境因子和主要微生物菌群丰度具有相反的调控作用。
图4中T2处理样本点在pH、OM变量上和在TPhA、EC变量延长线上的各垂直投影点到原点的平均距离总体上大于T3和T4处理,表明T2处理样本点与关键土壤环境因子 (pH、OM、TPhA、EC)之间的相互作用要强于T3和T4处理。因此,进一步使用LefSe分析 (LDA effect size analysis)挖掘不同分类水平上CK、T1和T2处理的标志性微生物(biomarker),以评估酚酸胁迫和施用复合颗粒处理下土壤细菌和真菌的差异物种。筛选标准为:线性判别分析值 (LDA值)不小于4,且显著性检验P值小于0.05。从分析结果可知,酚酸胁迫T1处理与CK相比 (图5),显著富集的细菌群落有酸杆菌门,富集的真菌有被孢霉门和子囊菌门,其中,被孢霉门中的被孢霉属 (Mortierella)、子囊菌门中的镰孢菌属(Fusarium)、Condenascus、圆孢霉属 (Staphylotrichum)、假散囊菌属 (Pseudeurotium)均发生显著富集。生物炭-过氧化钙复合颗粒处理 (T2)与酚酸胁迫处理(T1)相比 (图5),显著富集的细菌有拟杆菌门和绿弯菌门,显著富集的真菌有罗兹菌门。进一步对筛选出的biomaker在不同组样本中的丰度进行比较(图6)。与CK相比,T1处理显著提高了酸杆菌门、被孢霉门、子囊菌门、镰孢菌属、被孢霉属的相对丰度。而与T1处理相比,施用复合颗粒T2处理对以上微生物的丰度具有抑制作用,同时,与T1处理相比,T2处理土壤中拟杆菌门、绿弯菌门和罗兹菌门的相对丰度均明显增加。
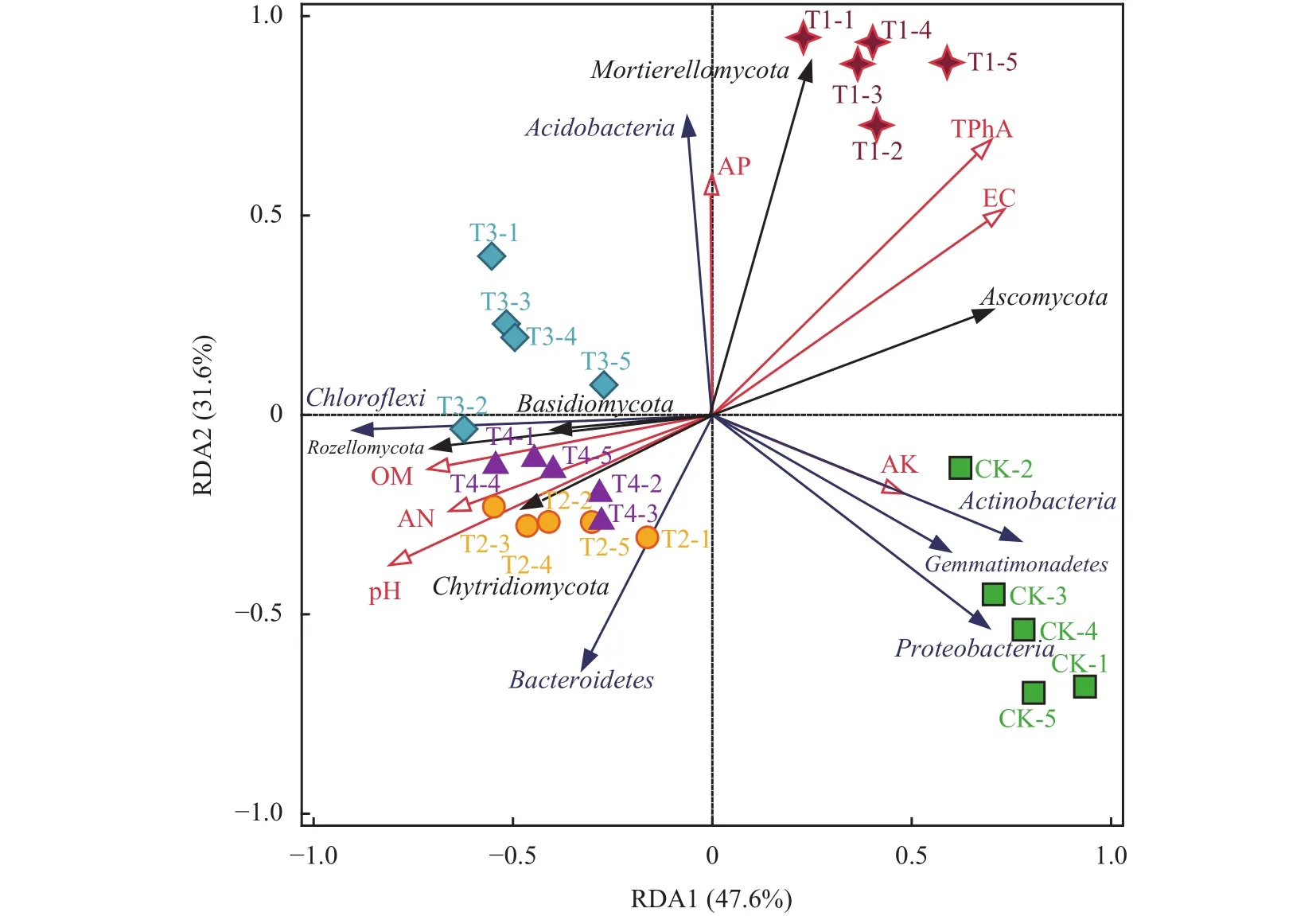
图4 主要细菌、真菌群落与土壤环境因子间的冗余分析Fig.4 Redundancy analysis of major bacterial and fungal communities, and rhizosphere soil environmental factors
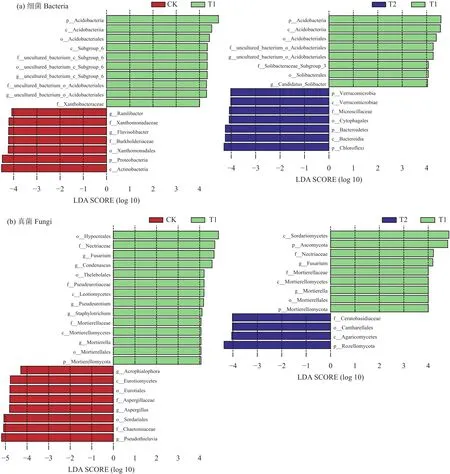
图5 基于LEfSe分析的土壤样品细菌(a)与真菌(b)分类信息的LDA值柱状分布图Fig.5 LDA distribution column map based on LEfSe analysis of soil sample classification information of bacteria (a) and fungi (b)
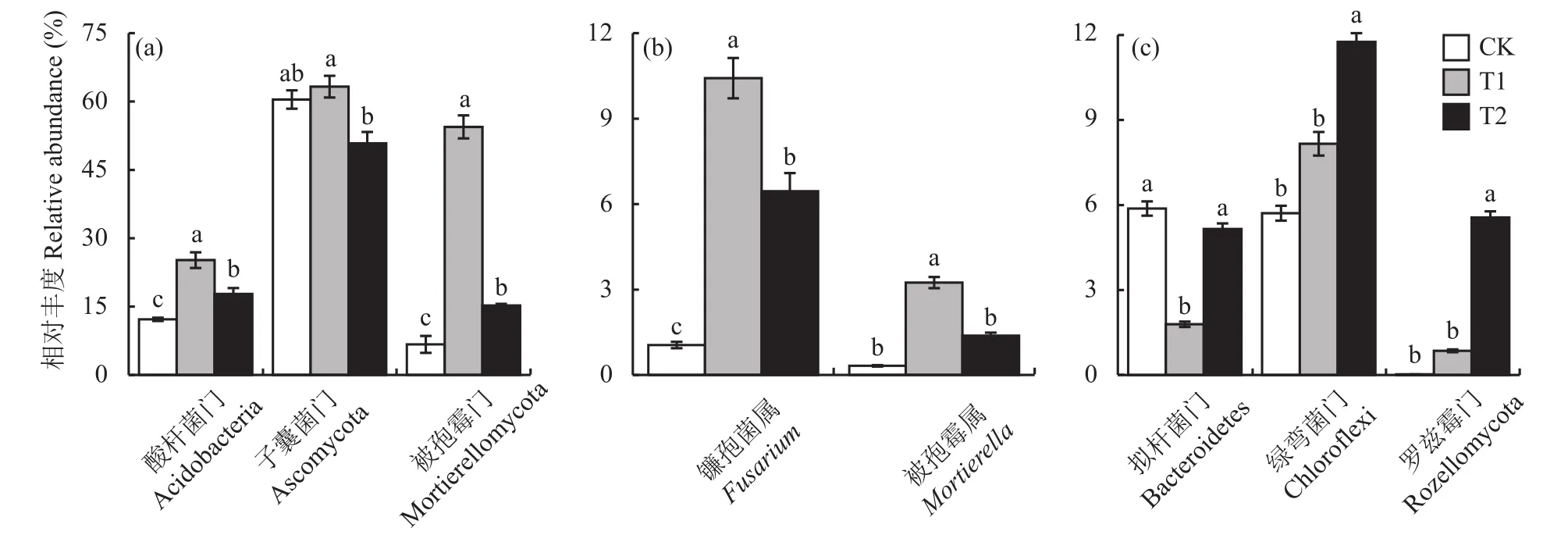
图6 不同处理根际土壤中标志性微生物的相对丰度比较Fig.6 Comparison of relative abundances of biomarker in rhizosphere soil under different treatments
3 讨论
3.1 施用生物炭-过氧化钙复合颗粒可有效去除土壤中残余酚酸,改善土壤理化性状
各类化感物质中,酚酸类物质受关注度最高且具有明显的化感活性。已有研究表明,连作条件下土壤中逐年累积的酚酸会导致土壤pH下降,EC值升高,并对与土壤碳循环有关的蔗糖酶和多酚氧化酶及与氮素转化相关的脲酶和蛋白酶均表现出活性抑制效应,从而导致土壤中有机质和有效氮含量出现下降的现象[34-36]。生物炭-过氧化钙复合颗粒中生物炭表面含有丰富的碱性官能团,且过氧化钙遇水分解生成强碱性的氢氧化钙,因此复合颗粒施用于土壤中能够有效地改善土壤酸性。同时,以生物炭为原料制备所得的复合颗粒孔隙结构丰富,能增强土壤中水分和养分的吸持,使土壤EC值显著下降,有机质及碱解氮含量增加[37]。生物炭-过氧化钙复合颗粒能够通过“吸附-氧化”协同作用显著减少土壤中的残余酚酸[38],此外,复合颗粒的施用可能加快土壤中微生物对酚酸的代谢降解。本研究中,复合颗粒可能通过以上两种非生物与生物降解途径实现对番茄根际土壤中酚酸的有效去除。
3.2 施用生物炭-过氧化钙复合颗粒可提高土壤微生物多样性,调控微生物群落结构
研究表明,土壤中累积的酚酸会使土壤酸化,养分失衡,造成多数细菌可利用的底物减少,从而导致细菌多样性下降[5,39]。而酸杆菌门是嗜酸菌,土壤酸碱度下降,对该类细菌的生长繁殖具有促进作用,同时酸杆菌可利用广泛的底物,且偏好营养不良的生态位[40-41]。子囊菌门和被孢霉门已被证实在连作条件下会大量累积,且与作物枯萎病发病率呈正相关[42],其中酚酸对子囊菌门中的镰孢菌属具有明显的生长刺激作用,是造成连作条件下番茄等作物枯萎病发生的关键原因之一[43-44]。本研究通过根际土壤微生物高通量测序也得到类似结果,即施加外源酚酸后土壤细菌多样性整体下降而真菌群落多样性增加,并驱动根际土壤微生物区系由“细菌型”演变为“真菌型”。物种组成分析和组间差异显著性分析结果进一步表明,外源酚酸的施用导致土壤酸杆菌门、子囊菌门、被孢霉门、镰孢菌属和被孢霉属的相对丰度显著增加。
施用生物炭-过氧化钙复合颗粒能够阻控土壤酸化,改善土壤养分状况,从而一定程度地恢复土壤生态平衡。研究发现,施用生物炭-过氧化钙复合颗粒后,土壤细菌多样性与酚酸胁迫处理相比显著增加,同时,拟杆菌门、绿弯菌门和罗兹菌门的相对丰度也明显提高。其中拟杆菌门和绿弯菌门已被证实具有较多根际促生菌和病原拮抗菌,是土壤健康的标志性微生物种群[40,45]。生物炭-过氧化钙复合颗粒的施用还显著减少番茄根际土壤中酸杆菌门、子囊菌门、被孢霉门、镰孢菌属和被孢霉属的相对丰度。生物炭对土壤微生物群落的介导效应已被广泛证实[46],Wu等[47]研究发现,生物炭会对太子参根际土壤细菌和真菌的丰度及代谢特征产生刺激作用,尤其显著影响镰孢菌的代谢过程并抑制菌丝体生长,从而有效降低病原菌丰度。与酚酸胁迫处理(T1)相比,本研究中施用生物炭-过氧化钙复合颗粒(T2)处理与施用生物炭 (T3)处理分别使镰孢菌属的相对丰度下降38.1%和36.6%,可见两种材料均对土壤中镰孢菌属具有抑制作用,且生物炭-过氧化钙复合颗粒的抑制效果更优。
3.3 施用生物炭-过氧化钙复合颗粒可减轻番茄酚酸化感胁迫,提高果实产量和品质
酚酸类化感物质对作物的生长表现出直接的抑制胁迫和间接的刺激作用。研究发现,高浓度的酚酸类物质会导致植物产生氧化应激效应,通过脂质过氧化作用造成细胞膜结构与功能受损,进而影响植物的离子吸收、细胞分裂、光合作用等生物学功能[48-50]。本研究发现,外源酚酸胁迫处理显著降低番茄植株根系活力和生物量,果实产量和糖酸比也出现不同程度地下降,可见本试验所采用的酚酸胁迫处理明显抑制了番茄植株营养生长与生殖生长。添加生物炭-过氧化钙复合型土壤调理剂对酚酸胁迫下番茄苗生长表现出一定的促进作用,有效提高根系活力和根冠比,且果实产量和果实糖酸比均获得不同程度地提高。
已有研究证实,施用生物炭能够有效促进番茄根系生长并增强叶片光合作用,从而提高果实产量、改善果实品质[51-53]。本研究比较了施用生物炭-过氧化钙复合颗粒和生物炭颗粒两种处理方式,结果表明复合颗粒对酚酸胁迫条件下番茄的促生、增产、提质效果要优于生物炭。
为探究生物炭-过氧化钙复合颗粒在酚酸胁迫下促进番茄生长及提高果实品质的机制,本研究通过相关性与关联分析和差异显著性分析,进一步揭示“复合颗粒-土壤环境因子-根际土壤微生物-植株”之间的互作关系。本研究表明,与番茄果实产量及品质极显著正相关的土壤环境因子为土壤pH和有机质,关键微生物有拟杆菌门及壶菌门;而土壤环境因子中总酚酸、EC值,以及微生物中酸杆菌门、子囊菌门丰度均与番茄果实的产量及品质具有负相关关系。施用生物炭-过氧化钙复合颗粒可以一定程度地提高土壤pH、有机质,降低土壤残余总酚酸和EC值。上述土壤环境因子变化可促进拟杆菌门、壶菌门等与番茄果实产量和质量显著正相关的微生物种群丰度上升,而抑制酸杆菌门、子囊菌门等与果实产量和品质显著负相关的微生物种群丰度。
4 结论
在诸多土壤环境因子中,总酚酸、pH、有机质、EC是驱动微生物群落变异和调控番茄产量与品质的关键因子。生物炭-过氧化钙复合颗粒可有效降低土壤残余总酚酸含量和EC值,并提高土壤pH和有机质含量,恢复土壤细菌和真菌群落之间的均衡,降低酸杆菌门、子囊菌门等有害真菌的相对丰度,进而有效缓解番茄酚酸胁迫,提高番茄植株根系活力,增加果实产量,并提高果实可溶性糖含量、维生素C含量和糖酸比。因此,生物炭-过氧化钙复合颗粒是非常有应用前景的茄果类蔬菜连作障碍土壤改良剂。
