2016—2020年浙江地区猪圆环病毒2型分子流行病学分析
2022-11-08徐丽华苏菲李军星余斌叶十一杨富文邓利荣毛慧敏袁秀芳
徐丽华,苏菲,李军星,余斌,叶十一,杨富文,邓利荣,毛慧敏,袁秀芳*
(1.浙江省农业科学院畜牧兽医研究所,杭州 310021;2.江山康农农业发展有限公司,浙江 衢州 324100)
猪圆环病毒(porcine circovirus,PCV)属于圆环病毒科(Circoviridae)圆环病毒属(Circovirus),为无囊膜单股环状负链DNA 病毒。该病毒目前共分为猪圆环病毒1 型(PCV type 1, PCV1)、PCV2 和PCV3 3种血清型,PCV1首先于1974年在猪肾传代细胞系(PK-15 细胞)中发现,对猪没有致病性[1]。PCV3由PALINSKI等[2]于2016年首次报道,分离自发生猪皮炎和肾病综合征(porcine dermatitis and nephropathy syndrome,PDNS)的母猪脏器和流产胎儿。PCV2 是引起断奶仔猪多系统衰竭综合征(postweaning multisystemic wasting syndrome,PMWS)、猪呼吸道疾病综合征(porcine respiratory disease complex,PRDC)和猪繁殖障碍等相关疾病的主要病原,目前在全球养殖场蔓延和流行,给养猪业造成了巨大的经济损失[3]。
PCV2基因组大小为1 766~1 768 bp,主要包含5 个开放阅读框(open reading frames, ORFs),其中以ORF1基因和ORF2基因最为重要。ORF1基因含有945 个碱基,编码314 个氨基酸,表达PCV2 滚环复制相关的Rep 蛋白,也是相对保守的蛋白;ORF2基因位于DNA的互补链上,含有702或705个碱基,编码234或235 个氨基酸,是病毒的主要结构蛋白(Cap蛋白),与病毒的免疫原性有关,具有较高的突变率。根据全基因遗传距离和ORF2基因的遗传距离,PCV2被分为PCV2a、PCV2b和PCV2c 3个基因型。近年来,有学者根据ORF2基因序列的进化树分析,将PCV2 分为5 个基因型,即PCV2a、PCV2b、PCV2c、PCV2d 和PCV2e[4]。其中PCV2e 在传统分类上属于PCV2a的一个分支,而PCV2c仅在丹麦和巴西有报道,且与猪圆环病毒相关疾病(porcine circovirus associated diseases,PCVAD)无关,而PCV2a、PCV2b 和PCV2d 均 已 经 被 证 实 与PCVAD 有 关[5-6]。根据PCV2的遗传演化轨迹,在2000年前PCV2a是国内优势基因型,随后由于基因变异,PCV2b 逐渐代替PCV2a,而近几年在中国和其他国家PCV2d已逐渐成为新的优势基因型[4,6-8]。
本研究对2016—2020 年间采集自浙江省不同地区猪场的疑似病料,利用聚合酶链反应(polymerase chain reaction,PCR)技术进行PCV2 检测和阳性率统计。对部分阳性样品进行全基因组克隆和测序,并与GenBank 中收录的PCV2 疫苗株和参考株序列进行遗传变异分析,以期了解和掌握浙江地区PCV2 的流行动态和遗传变化趋势,从而为PCV2的防控提供理论依据。
1 材料与方法
1.1 主要试剂和仪器
病毒核酸提取试剂盒(生产批号为Y503-G12、Y502-A10 等)购自江苏省济凡生物科技(常州)有限公司;高成功率PCR酶KOD FX购自日本TOYOBO公司;1 000 bp DNA分子标志物、dNTPs、TOP10感受态细胞等均购自生工生物工程(上海)股份有限公司;胶回收试剂盒、pMD19-T 载体均购自宝生物工程(大连)有限公司;无水乙醇、琼脂糖、胰蛋白胨、酵母浸出物、氨苄西林(ampicillin,AMP+)等化学试剂均为国产或进口分析纯。PCV2阳性毒株由本实验室分离保存。
ChemiDocXRS+凝胶成像系统购自美国Bio-Rad公司;Tone 96G PCR仪购自德国Biometra公司;D-37520 Osterode 高速离心机购自德国Sigma 公司;Auto-Pure 32SH 全自动病毒核酸提取仪购自杭州奥盛仪器有限公司;Tissueiyser-24全自动样品快速研磨仪购自上海净信科技公司。
1.2 病料采集及处理
于2016—2020 年间,采集浙江省不同地区(杭州、嘉兴、湖州、台州、宁波、温州、绍兴、金华和衢州)疑似PCV2 感染猪的病料,包括血液、肺脏、脾脏、腹股沟淋巴结和胃底组织等样品,于-70 ℃冰箱中保存,备用。
取保存的病料0.1 g,置于小型离心管(Eppendorf 管)中,加入pH 7.4 磷酸缓冲盐液(phosphate buffer saline, PBS)0.5 mL,用全自动研磨仪(90 Hz,60 s)研磨成匀浆,于-70 ℃条件下冻融3 次,以8 000g在4 ℃下离心5 min,取上清液0.15 mL,用病毒核酸提取试剂盒提取核酸,操作过程严格按照试剂盒说明书进行,吸取病毒核酸50 μL,于-20 ℃条件下保存,备用。
1.3 引物设计
下载GenBank中收录的PCV2全基因组序列,利用Primer Premier 5软件设计PCV2检测引物(F1/F2)和PCV2 全基因组测序引物(PCV2-1039F/PCV2-1039R,PCV2-867F/PCV2-867R),委托生工生物工程(上海)股份有限公司合成,引物信息见表1。
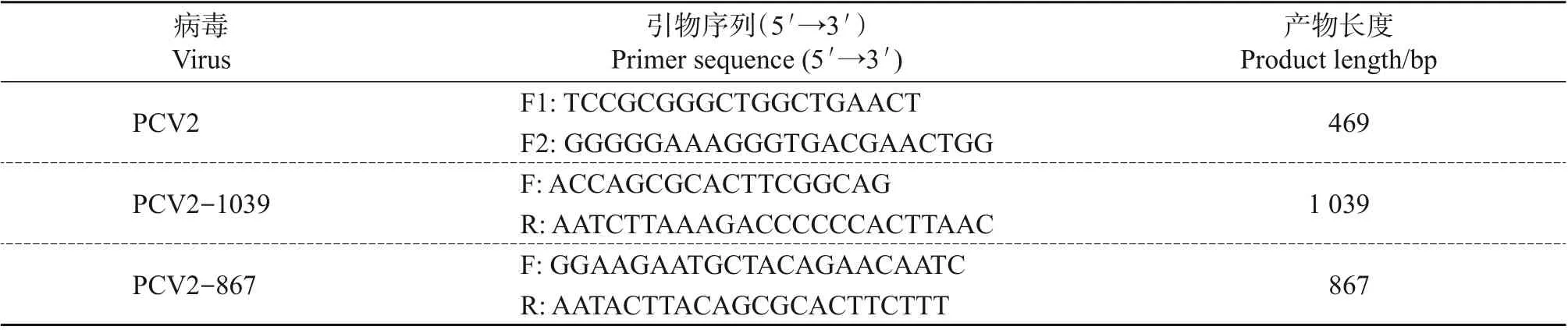
表1 用于PCR的引物Table 1 Primers used for PCR
1.4 样品中PCV2 的PCR 检测
以上述提取的病毒核酸为模板,用病原检测引物进行PCV2 检测。PCR 反应体系(25 μL):2×PCR缓冲液12.5 μL,2 mmol/L dNTPs 5.0 μL,上、下游引物各1.5 μL(浓度为5 μmol/L),KOD FX(1.0 U/μL)0.5 μL,病毒核酸DNA 2.0 μL,ddH2O 2.0 μL。混匀分装后置于PCR 扩增仪中,扩增条件为:94 ℃预变性2 min;98 ℃变性10 s,68 ℃退火及延伸1 min,35 个循环;68 ℃延伸7 min。反应完成后,取5 μL PCR 产物经1.2% 琼脂糖凝胶电泳检测,用ChemiDocXRS+凝胶成像系统观察拍照。
1.5 PCV2 全基因组扩增和序列测定
选取PCV2 检测为阳性的部分样品的核酸,用2对全基因组序列扩增引物分别进行扩增。PCR反应体系为25 μL,反应液配制和扩增条件同1.4 节。扩增完成后,加入0.5 μL rTaq聚合酶,72 ℃延伸10 min。反应完成后,取5 μL PCR产物经1.2%琼脂糖凝胶电泳检测并切胶回收。
将胶回收产物克隆到pMD19-T 载体中,转化入TOP10感受态细胞,随后均匀涂布于含有氨苄西林抗性的Luria-Bertani(LB)平板上,于37 ℃条件下培养12~14 h。挑取单菌落,分别接种至LB培养基(含AMP+抗性)中,经菌液PCR 鉴定正确后送至生工生物工程(上海)股份有限公司测序。
1.6 序列分析
将1.5 节中获得的序列进行拼接得到完整的PCV2 序列,运用DNAStar 7.1 软件和MEGA 5.1 软件,与GenBank 中收录的16 株PCV2 参考株序列进行同源性比较,以及进行核苷酸和氨基酸的遗传进化分析。参考株序列见表2,其中PCV2-SH 株、PCV2-LG 株和PCV2-ZJ 株为疫苗株序列名称。PCV2-SH 株序列从上海发病场克隆测序得到,PCV2-ZJ 株序列从浙江发病场克隆测序得到,两者均属于PCV2b 型,全基因组长度为1 767 bp。PCV2-LG 株序列从吉林发病场克隆测序得到,属于PCV2a 型,全基因组长度为1 768 bp。其余参考株来自不同国家,全基因组长度为1 766~1 768 bp。
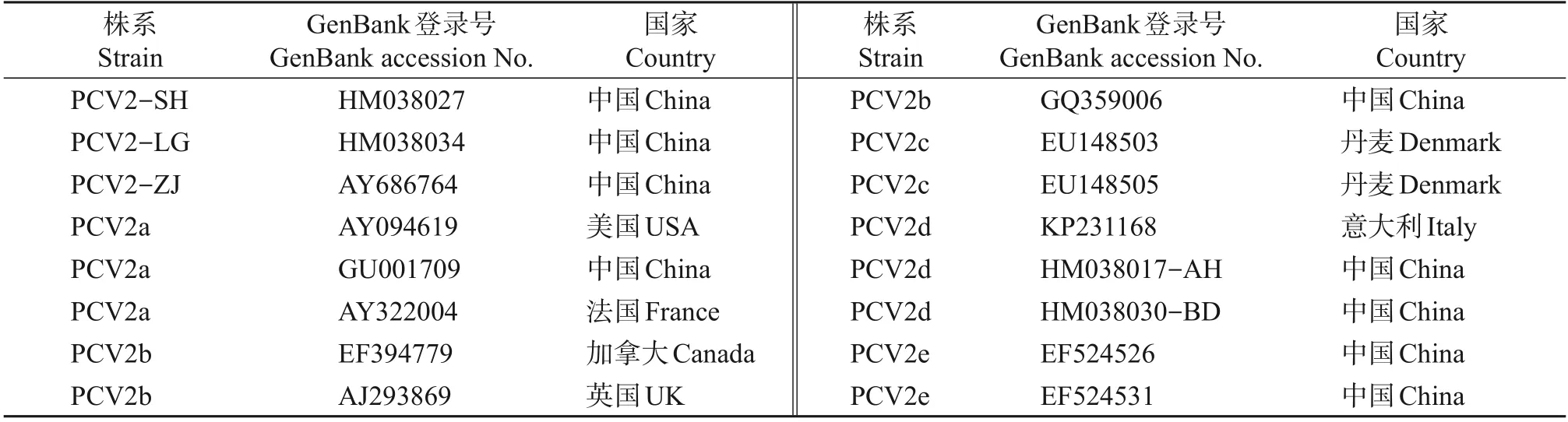
表2 PCV2参考株信息Table 2 Information of PCV2 reference strains
2 结果与分析
2.1 临床样品的检测结果
对2016—2020年间浙江省不同地区的1 725份疑似猪圆环病毒2 型感染的临床样品进行PCV2 的特异性扩增,同时设立阳性对照组和阴性对照组。凝胶电泳结果显示扩增出469 bp的特异性条带,与预期片段长度相同(图1)。其中,检测到PCV2阳性样品为359 份,平均阳性率为20.8%(359/1 725)。2016—2020 年PCV2 阳性率分别为38.1%、23.2%、24.1%、12.5%和10.7%。PCV2阳性率逐年下降,2016年最高(38.1%),而2020年下降到10.7%(表3)。浙江省不同地区均检测到PCV2,且不同区域间阳性率没有明显差异,说明PCV2 在浙江省不同地区感染较为普遍,且没有存在明显的地区差异。

图1 猪圆环病毒2型PCR扩增结果Fig.1 PCR amplification results of PCV2
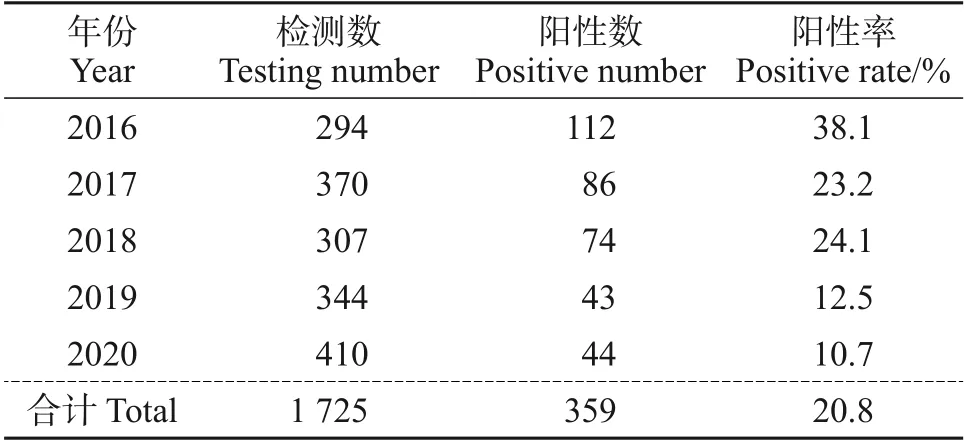
表3 不同年份(2016—2020年)PCV2的阳性率Table 3 Positive rates of PCV2 from 2016 to 2020
2.2 全基因组扩增和测序结果
在PCV2检测为阳性的样品中按年份随机选取条带较亮的样品共36份,其中,2016年7份、2017年7 份、2018 年9 份、2019 年7 份、2020 年6 份,进行PCV2 全基因组的扩增、克隆和序列测定。经测序获得36 株PCV2 全基因序列,其中全长为1 766 bp的有4 株,1 767 bp 有25 株,1 768 bp 有7 株。序列根据样品采集区域名称的拼音首字母和采集年月日(YYYYMMDD)进行组合命名。
2.3 PCV2 全基因组序列同源性比较和序列分析
将测序所得36 株序列与GenBank 中的16 株参考株序列进行同源性比较和遗传进化分析。结果表明:36 株PCV2 序列的核苷酸同源性为94.0%~99.9%,与国产疫苗株(SH 株、LG 株和ZJ 株)的核苷酸同源性为94.7%~98.5%,与13 株参考株的核苷酸同源性为92.9%~99.8%,其中DQ20200115 株与EU148505 株亲缘关系最远,同源性为92.9%;XS20160217 株与PCV2d-AH 株亲缘关系最近,同源性为99.8%。按照基因型分类,36 株PCV2 序列分为3 个基因型,其中11 株为PCV2a 型,8 株为PCV2b 型,17 株为PCV2d 型,没有分离到PCV2c型和PCV2e型。随时间迁移,属于PCV2d亚型的序列数量呈增加趋势,其中2019年最多,有7株(图2)。

图2 基于PCV2全基因组的系统进化树Fig.2 Phylogenetic trees based on sequences of PCV2 whole genome
2.4 PCV2 ORF1 基因核苷酸及氨基酸序列的同源性比较
本实验测得36株PCV2序列的ORF1基因全长均为945 bp,所得序列之间的核苷酸同源性为97.1%~100.0%,其中HZ20170510 株与JX20170804 株、DQ20160422 株与LX20181101 株、JH20200608 株与QZ20200521 株、PY20190323 株与WZ20181116 株核苷酸同源性最高(100.0%);与国产疫苗株(SH株、LG株和ZJ 株)的核苷酸同源性为96.9%~98.9%;与13株参考株的核苷酸同源性为95.9%~100.0%,其中JS20190419 株和PCV2e EF524526 株同源性最低(95.9%),QZ20200521 株、JH20200608 株和PCV2d HM038030株同源性最高(100.0%)。各PCV2序列的ORF1基因可编码315个氨基酸,36株PCV2序列的氨基酸同源性为98.1%~100.0%,与国产疫苗株(SH株、LG株和ZJ株)的氨基酸同源性为97.8%~100.0%,与13株参考株的氨基酸同源性为97.4%~100.0%。
2.5 PCV2 ORF2 基因核苷酸及氨基酸序列的同源性比较
本实验测得36 株序列PCV2ORF2基因全长分别为705 bp(16 株)和702 bp(20 株)。所得序列之间的核苷酸同源性为88.7%~100.0%,其中DQ20201115株与JX20200715株、HZ20200323株与QZ20200521株同源性最高(100.0%);与国产疫苗株(SH 株、LG 株和ZJ 株)的核苷酸同源性为90.2%~99.6%;与13 株参考株的核苷酸同源性为85.0%~100.0%。另外所得序列DQ20201115株、JX20200715株和参考株PCV2c EU148503、PCV2c EU148505 的同 源 性 最 低(85.0%),JS20190419 株 与PCV2d HM038017 株、PCV2d HM038030 株的同源性最高(100.0%)(图3)。

图3 36株序列ORF2基因核苷酸序列同源性比较Fig.3 Homology comparisons of ORF2 gene nucleotide sequences of 36 strains
PCV2ORF2基因可分别编码235或234个氨基酸,相互之间氨基酸同源性为86.3%~100.0%,与国产疫苗株(SH株、LG株和ZJ株)的氨基酸同源性为87.2%~98.3%,与13 株参考株的氨基酸同源性为82.1%~100.0%。
2.6 Cap 蛋白的氨基酸序列分析
ORF2基因编码病毒的主要结构蛋白(Cap 蛋白),其唯一糖基化位点位于第143—145位氨基酸,测序所得36株序列糖基化位点均为天冬酰胺(N)、酪氨酸(Y)、丝氨酸(S)(NYS),属于高度保守[9-11]。从图4可以看出,不同亚型的氨基酸序列差异明显,同一亚型的氨基酸序列也存在个别点突变,但没有发现插入或缺失。变异位点分布在Cap蛋白的不同位置,但其变异度比较高的主要区域为第53—91、121—136、190—210位氨基酸。另外也存在一些散在的突变位点,分别位于第30、85、101、124、161、171位氨基酸等,这些点突变没有显示和亚型相关,但随时间推移可能会出现新的抗原变异。

图4 PCV2 Cap蛋白的氨基酸序列多重比对分析Fig.4 Multiple alignment analysis of amino acid sequences of PCV2 Cap protein
与PCV2-LG 株氨基酸序列相比,所得序列中PCV2a 亚型的突变位置主要为T47A、R63S、A68S、M72L、I76L、N77D、V123I、T131M/I、R132K、A133V、T134P、L136Q、L185M;与PCV2-ZJ 株氨基酸序列相比,PCV2b 亚型的突变位置主要为R59K、K63R、A190T;与PCV2d HM038030 株序列相比,PCV2d 亚型的突变位置主要为R168G;与疫苗株序列相比,优势基因型PCV2d 亚型的特异性突变位点主要集中在I53、N68、L89、T121、R168、I215。而Cap 蛋白的B 细胞抗原表位分别位于第47—63、65—87、113—139、165—200 位氨基酸和C端的最后4位氨基酸,T细胞抗原表位分别位于第61—160、210—230位氨基酸[10-12]。显示不同亚型的突变位点均位于B细胞和T细胞抗原表位区域,抗原表位在免疫压力下易发生突变,从而改变PCV2蛋白的抗原性和致病力。另外,17株PCV2d亚型的序列中,除了序列LQ20171115 和LS20200831外,其余15株蛋白质长度均为235个氨基酸,分析显示其终止密码子发生突变,Cap 蛋白C 末端增加了1个赖氨酸K,导致了Cap蛋白大小的改变。
3 讨论与结论
自2000 年首次证实PCV2 在我国猪场感染以来,全国各地众多猪场陆续发现PCV2 的流行和暴发,其感染率高且常和猪蓝耳病毒、猪伪狂犬病毒、副猪嗜血杆菌等混合感染,给养殖企业带来不可估量的经济损失[7-8,12-15]。本研究对2016—2020年间采集的浙江省不同地区1 725 份疑似猪圆环病毒2 型感染的临床样品进行了PCV2 检测,样本平均阳性率为20.8%(359/1 725),其中2016—2020年PCV2阳性率分别为38.1%、23.2%、24.1%、12.5%和10.7%。5 年来PCV2 阳性率总体呈逐年下降趋势,原因有:一方面2014年浙江省实施“五水共治”,对省内养猪个体户的养殖场进行了拆除整治,养猪环境得到很大改善,猪场发病率明显下降;另一方面2018 年非洲猪瘟在我国多个省市呈点状暴发后,全国禁止生猪跨省调运,阻碍了猪场疫病的传播。与王小敏等[10]在华东地区(2010—2015 年)、邓文芳等[11]在华中地区(2017—2018年)的PCV2分子流行病学调查相比,本研究中PCV2 阳性率较低,可能与近几年PCV2灭活疫苗在浙江地区普遍使用和养殖企业生物安全意识提高有关;与吴明臻等[12]在浙江省金华地区的PCV2流行病学调查相比,本研究中PCV2阳性率较高,其原因可能是该研究中样品来源是猪场血清样本,而本研究采集的都是临床疑似PCV2 发病样品。
本研究中,2016—2020年浙江省不同地区36株PCV2全基因序列的核苷酸长度为1 766~1 768 bp,核苷酸同源性为94.0%~99.9%,与国产疫苗株序列(SH 株、LG 株和ZJ 株)的核苷酸同源性为94.7%~98.5%,与13 株参考株序列的核苷酸同源性为92.9%~99.8%,这与吴瑗等[13]在浙江省及周边地区(2007—2012年)的PCV2全基因序列遗传分析结果相似,说明浙江地区PCV2毒株序列整体比较稳定,没有明显的时间差异和地域差异,与国内外不同PCV2毒株序列的差异也不大。
目前,大量文献[9-12,16-18]报道了国内PCV2 的流行逐渐从PCV2b 亚型向PCV2d 亚型转变。浙江省不同地区PCV2 流行情况是PCV2a(11/36)、PCV2b(8/36)和PCV2d(17/36)3 种亚型共存,没有发现PCV2c、PCV2e亚型,目前PCV2d是浙江地区PCV2流行的优势毒株亚型。接种疫苗是应对PCV2感染的主要手段,而国内现有的PCV2 商品化疫苗仅针对PCV2a(LG株)和PCV2b(SH株和ZJ株)2个亚型,对其他亚型毒株感染不能提供完全的交叉保护力,致使在临床PCV2感染中疫苗的防控效果不甚理想。
PCV2ORF1基因编码与病毒复制相关的Rep蛋白。本研究中36株序列ORF1基因编码的蛋白质长度均为315 个氨基酸,所得序列相互间氨基酸同源性为98.1%~100.0%,与国产疫苗株序列的氨基酸同源性为97.8%~100.0%,与参考株序列的氨基酸同源性为97.4%~100.0%。说明PCV2ORF1基因相对比较稳定,变异不大。
相对于Rep蛋白,PCV2ORF2基因编码的Cap蛋白是病毒刺激机体产生中和抗体的主要结构蛋白。Cap蛋白易突变,其变异程度远远高于其他蛋白,使PCV2毒株的变异进化、新毒株的不断出现成为可能。本研究中,36株序列中该蛋白质的长度分别为234或235 个氨基酸,各序列间氨基酸同源性为86.3%~100.0%,与国产疫苗株序列的氨基酸同源性为87.2%~98.3%,与参考株序列的氨基酸同源性为82.1%~100.0%,说明大部分序列的ORF2基因发生了变异。位于不同亚型的氨基酸序列差异明显,同一亚型的序列也存在个别点突变,但没有发现插入或缺失。在B 细胞抗原表位区(第47—63、65—87、113—139、165—200 位氨基酸和C 端最后4 位氨基酸)和T 细胞抗原表位区(第61—160、210—230 位氨基酸)的氨基酸序列存在较多变异,这与赵军等[14]在四川地区对PCV2的调查分析结果相似。这些位点的变异可能会改变Cap 蛋白的免疫原性,特别是N75、I76、T131、I206位点的变异,因为这些氨基酸位点处于Cap 蛋白的免疫反应区域内[15-18],有可能导致PCV2 的致病性增强。另外,PCV2d 亚型中有15 株序列在C 端出现了1 个赖氨酸K 的延伸,这与郭龙军等[19]、王小敏等[10]的报道一致;OLVERA等[20]也证实由于ORF2基因终止密码子发生突变,Cap 蛋白C端出现了1个赖氨酸延伸的现象。赖氨酸是一种限制性氨基酸,对机体免疫系统活性有一定的影响,推测该氨基酸的延伸也可能影响Cap 蛋白的免疫功能或增加病毒的致病性。
综上所述,本研究对2016—2020年间浙江省不同地区PCV2 流行情况进行了调查,并对全基因组序列和Cap 蛋白进行了遗传变异分析,结果显示浙江地区PCV2d为优势基因型。本研究阐明了近5年浙江地区PCV2的流行变化趋势,丰富了PCV2的流行病学资料,为浙江地区养殖企业针对PCV2 感染疫苗的选择和研究机构新型疫苗的研发提供了一定的理论依据。
