茎和根几个次生结构的探讨
——植物学教材质疑(九)
2022-11-05黎维平
黎维平
(湖南师范大学生命科学学院,中国湖南 长沙 410081)
中外植物学教材中,有几个被子植物的次生结构长期被误解或忽视,近年新编教材和修订版本对教材的改进也都未涉及这几个结构。在几十年的教学中,笔者致力于突破这些难点。本文较详细地分析了这些问题,并提出了两个有关次生结构的探究性实验,供植物学理论和实验教材的编写者、修订者参考。
1 切向面上的年轮呈现“V”形还是“倒V”形?
木本植物次生生长的主要产物之一是次生木质部。次生木质部(木材)三切面(横切面、切向切面和径向切面)的形态特征以年轮或年轮线、木射线的形态为标志。
1.1 “V”形还是“倒V”形?
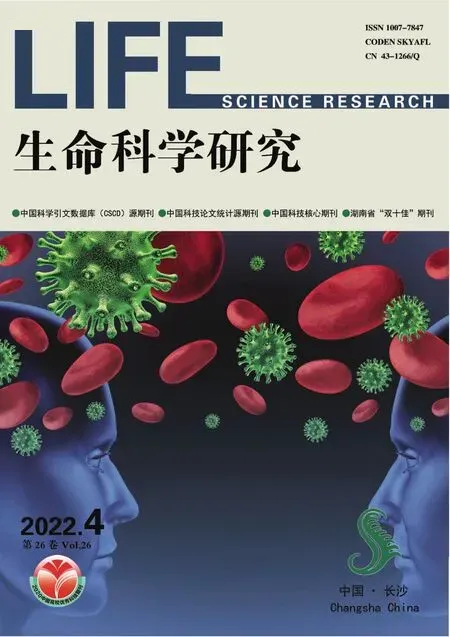
在切向切面上,年轮、年轮线为“V”形是中外植物学教材[1~21]的共识,图1A被多本教材采用[1~4,6,8~9,11~12,14~16,18~21],也有教材采用图 1B[17]。这是一个不小的错误。设想一下,一株真双子叶植物的茎,其基部横切面上有50个年轮,最新形成的年轮在最外方;从茎基到主干上端仅具1个年轮处,剥离出这个年轮,其为圆台形;径向切之,切点首先在圆台上方,继续往下切后,切面必为“倒V”形。所以,木本茎切向切面上的年轮形态应如图1C所示,但只有少量教材[22~24]持有这种观点。图1A和B的另一个问题是,相邻年轮的“V”形结构顶点相距太近,因为这个距离大致相当于茎在一年中的生长高度,应该远大于年轮宽度,而不是像图中显示的那样二者仅相差2~4倍。
1.2 “V”形年轮观点来源
十多年前,笔者质疑教材上关于木材切向切面的年轮图形时,有老师说见到的木材标本就和书本上所绘的“V”形一样。那么问题出在哪里呢?笔者在多年的观察中,也没有找到支持本文观点的完美实物。但笔者发现,同一木材切向切面上总可见到年轮既有“V”形的,也有“倒V”形的。原因是这些年轮通常是侧枝(枝迹)横切面上的年轮(枝迹的木质部)。侧枝大都与着生处的主干同龄或稍年轻;当对主干径向切时,就会对主干各节上的分枝进行不严格的横切(枝迹通常向上倾斜着穿越主干),结果木材径向切面上不可避免地会出现侧枝横切面上的年轮。多年生次生木质部的切向切面显示,其不受侧枝年轮干扰是不大可能的。年轮“V”形的结论可能是被侧枝枝迹年轮横切面误导了;图1A和B中年轮线纵向上的距离如此之小,也可用其实际上是侧枝枝迹年轮(髓心在裸子植物中偏向上方,而在被子植物中偏于下侧)的横切面来解释。所以,在受限于无理想实物照片的情况下,借助于几何图形,发挥学生的空间想象力,是教学中突破这一难点的可行途径。
1.3 年轮、晚材与年轮线在图上的标示
在年轮三切面照片、墨线图上,各教材常将标为年轮线[15]或年轮[1~2,8,20,24]的引线指向年轮中窄窄的深色部分(图1A),导致学生常将这些深色线条形结构理解为年轮线。实际上,这部分是晚材,而年轮线不是任何结构,仅为前一年的晚材和次年的早材间的界线。提醒学生注意晚材和年轮线的区别是很有必要的。编写教材时,应在图中避免年轮线这个抽象的结构以实物的形式被指出;年轮也不应仅指向晚材,可将早材和晚材分别指出,然后将二引线汇集到年轮一词。
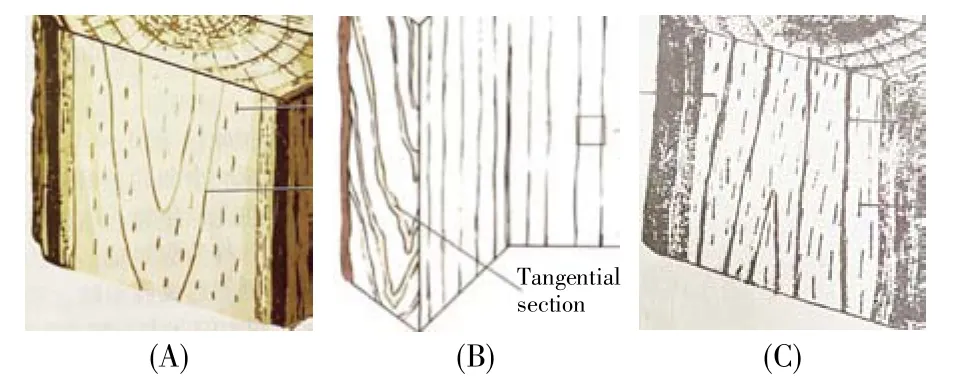
图1 木材切向面上的年轮图(A)引自马炜梁[15];(B)引自Mauseth[17];(C)引自徐有明[24]。Fig.1 Illustrations of annual rings in tangential sections of woods(A)Cited from Ma[15],showing V-shape rings;(B)Cited from Mauseth[17];(C)Cited from Xu[24].
2 幼茎横切面上的倒三角形射线是韧皮射线还是髓射线?
在有次生生长的一年生至几年生幼树茎的横切面上,形成层外方紧挨着的结构有韧皮部和数个倒三角形结构(下文简称“倒三角”)(图2)。“倒三角”是什么结构?对此,植物学教材有4种观点:1)称之为韧皮射线[8,11,14,25~29],expended phloem ray[5]或dilated phloem ray[13,29](膨大的韧皮射线),扩张的韧皮射线[22],或broad phloem ray(宽大的韧皮射线)[30];2)称之为扩增的射线细胞(proliferated ray cells[17])。根据其上下文[17]关系,实际上该教材仍将其当作韧皮射线的一种特殊类型;3)认为其为髓射线[3,7,12,19]或髓射线扩展区[31];4)忽略其存在,不标注其名称[15,32]。“倒三角”到底是什么结构?
2.1 “倒三角”主要是径向延长、切向拉宽的髓射线外段
髓射线是薄壁组织,为初生结构,位于维管束之间,连接髓部和皮层。当束间形成层发生后,髓射线被隔成两段:外侧位于初生韧皮部之间,内侧夹在初生木质部之间。在次生生长过程中,大部分植物的束间形成层向内、外添加射线薄壁细胞,延长射线。延长部分属于次生结构,所以此后的髓射线不再是纯粹的初生结构,而是两端为初生射线、中间为次生射线的嵌合射线,建议称之为“次生延长的髓射线(secondary extended pith ray,缩写为 SEPR)”。
在木栓形成层尚未出现在次生韧皮部处之前,SEPR在形成层外侧部分呈现倒三角结构。“倒三角”形成的机理包括两个方面。首先,束中形成层细胞来自近等径的髓射线细胞,其恢复分裂后为射线原始细胞,通过平周分裂延长髓射线。之后,束间形成层两侧的射线原始细胞转化成纺锤状原始细胞(而在束中形成层中,两类细胞间转化的方向通常相反,几乎都是由纺锤状原始细胞零零星星地转化为射线原始细胞,以增加射线数量),束间形成层的射线原始细胞越来越少,延长的髓射线越来越窄,致使原先大致等宽的外侧髓射线向内(向着束间形成层)逐渐成为“倒三角”。其次,茎因次生生长而增粗,各初生韧皮部间以及早期形成的次生韧皮部间的距离变大,特别是位于射线最外侧、初生韧皮部之间的髓射线细胞被明显拉宽,这也帮助了倒三角形态的形成。Mauseth[17]对倒三角结构的解释是,某些射线的薄壁细胞通过分裂和扩展而扩增(“Parenchyma cells of some rays have proliferated through cell division and expansion”[17])。射线细胞发生了扩展(“expansion”),即被拉宽,Mauseth[17]的这一观点与笔者的解释类似。但他没有注意到另一现象,即细胞越靠外被拉宽程度越大。特别是,他把“倒三角”外侧细胞较多归因于细胞分裂时,并未明确指出是什么细胞分裂。从“division and expansion”的逻辑主语是同一词“cell”来看,该细胞指的是射线细胞,这与笔者关于射线原始细胞由多到少的解释不同。“倒三角”两侧的次生韧皮部向内逐渐增加,韧皮部形成上窄下宽的梯形结构,这与束间形成层细胞逐渐由射线原始细胞转变成纺锤状原始细胞的现象十分吻合。当然,束中形成层通过细胞径向分裂而增宽,这也是次生韧皮部逐渐变宽的另一原因。
“倒三角”不一定都是SEPR的一部分,不排除早期也有束中形成层产生倒三角形射线,但其远不如髓射线的“倒三角”显著,也较少见。
在有些论文[32~33]中,双子叶植物一年生木本茎的模式图未将已有显著次生生长的髓射线韧皮部一侧画成倒三角形,而且将髓射线由里到外依次称为木射线(xylem ray)、射线细胞(ray cell)和韧皮射线(phloem ray)。髓射线外侧的倒三角形(横切面)是早期木本茎的普遍现象,应加以呈现,而所谓“射线细胞”实为形成层的射线原始细胞(ray initial)。最值得注意的是,不应将髓射线这种初生射线与次生射线(包括木射线和韧皮射线)混为一谈。
2.2 “倒三角”消失之因
“倒三角”是次生生长早期特有的现象,随着新的木栓形成层在次生韧皮部中出现,部分SEPR细胞也脱分化成为木栓形成层细胞,“倒三角”立即或逐渐被埋没在硬树皮之中。最终,新周皮内方的SEPR宽度与其他维管射线无差别,“倒三角”消失。此外,随着次生生长,SEPR数目固定不变,而次生射线不断增加,SEPR在茎的射线中所占比例越来越少,最终可被忽略;SEPR的初生结构部分已失去功能,初生韧皮部间和初生木质部间的髓射线细胞分别埋没在硬树皮和心材之中,剩下的SEPR均为次生结构,本质上与维管射线无异。就像此时区分束间形成层和束中形成层没有必要且也无可能一样,此时再要区分SEPR和维管射线,也是不仅没有必要(二者都是次生结构),也几乎没有可能(二者宽度和细胞类型均相似)。
2.3 束间形成层的其他活动情形
一些草本真双子叶植物,如茄科的白英(Solanum lyratum Thunb.),在其次生生长过程中,束间形成层一开始就是纺锤状原始细胞,通过切向分裂向内外分别产生维管组织和纤维细胞,并不延长髓射线,当然不会出现“倒三角”(黎维平,未发表)。棉[16](Gossypium sp.)和大豆[34](Glycine max)茎也存在此现象。
另一些植物,如葡萄(Vitis vinifera L.),其茎的束间形成层分为三部分,中间的细胞为纺锤状原始细胞,在原有维管束间的髓射线处增加1个新的维管束;而两侧的束间形成层细胞为射线原始细胞,其切向分裂延长出两条髓射线,因此,在其外侧也可见两个较小的“倒三角(IT)”(图2C)。
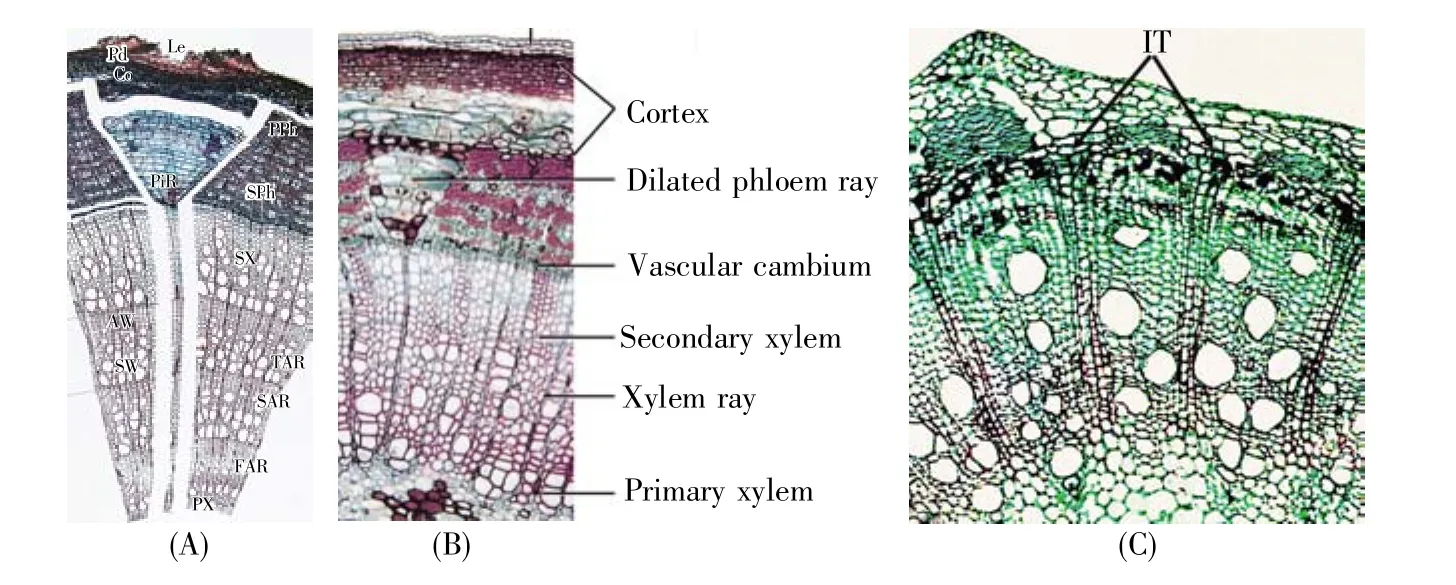
图2 木本茎横切面(局部),显示髓射线(A)5年生茎,其髓射线(PiR)已被与周边组织剥离。引自冯燕妮等[35];(B)1年生茎,引自Evert等[13];(C)葡萄茎横切面(自制、自拍),可见1束间形成层产生1次生维管束和2“倒三角(IT)”。Fig.2 Illustrations of transversal sections(a part)of wood stems,showing pith rays(medullary rays)(A)A five-year stem with a pith ray(PiR)stripped from the other adjacent tissues(cited from Feng et al.[35]);(B)A one-year stem with a pith ray called“dilated phloem ray”(cited from Evert et al.[13]);(C)The stem section(made and photographed by the author)of Vitis vinifera L.,showing two inverse trangles(ITs)produced by one interfascicular cambium.
3 根中“倒三角”射线
由于初生木质部和初生韧皮部相间排列,根无论有髓或无髓都不产生髓射线。但是根的次生结构中也会出现类似于“倒三角”的结构。这是何种结构?图3[2,35]未对其标注,而有的教材将其标为髓射线[1,9]或维管射线[12,21~22,28,34]。根中“倒三角”当然不是髓射线(初生射线),而是完完全全的维管射线(次生射线)。
根的形成层是完全的次生分生组织(茎的束中形成层是初生分生组织,来自原形成层的短期休眠细胞),大部分来自初生韧皮部内方及其与初生木质部之间的薄壁细胞,初期在横切面上为数个弧形的形成层片段;与原生木质部(木质部脊)相对的中柱鞘细胞恢复分裂后,加入形成层,将分离的弧形形成层片段连接起来,使早期的形成层在横切面上为波状形态。来自中柱鞘细胞的形成层部分为射线原始细胞,产生次生射线,即木射线和韧皮射线。
“倒三角”射线在根和茎中形成的机理显著不同。在根中,随着内方次生木质部增加,形成层被推向外方。外移的形成层为适应直径扩大的需求,分生组织细胞径向分裂。其中,来自中柱鞘细胞的形成层细胞为射线原始细胞,它们随着形成层直径增加逐渐增多。结果,由射线原始细胞所产生的子细胞分化而成的木射线在向外延伸中逐渐加宽,其在横切面上呈现为“倒三角”形。所以,根和茎中的“倒三角”有3点主要差异(图3B)[35]:1)根中的“倒三角”主要是次生木质部之间的木射线,而茎中的是韧皮部间的髓射线;2)在根的“倒三角”结构中,韧皮射线虽然量少,但为“倒三角”最宽的部分。根“倒三角”既含木射线,也含韧皮射线;而茎中的“倒三角”均是韧皮部间的射线,与木质部间射线无关;3)生长过程中,根的“倒三角”是由窄变宽、离心式延长,而茎的则是由宽变窄、向心式发展。
图3 根结构(A)根次生结构图解,引自陆时万等[2];(B)南瓜老根横切面图,引自冯燕妮等[35]。PXV:初生木质部导管;SXV:次生木质部导管。Fig.3 Illustrations of root structures(A)Secondary structures of root(cited from Lu et al.[2]);(B)An old root section of Cucurbita moschata(cited from Feng et al.[35]).PXV:Primary xylem vessel;SXV:Secondary xylem vessel.
4 栓内层细胞的辨识
经过维管形成层一定时间的活动,初生保护组织(表皮)因受到来自茎增粗的压力而面临崩溃。于是,某些皮层细胞,有的植物也可以是表皮细胞,它们恢复分裂成为木栓形成层,向内产生栓内层,并向外产生多层木栓层细胞(在原有气孔处可产生补充组织而形成皮孔),木栓层、木栓形成层、栓内层和皮孔共同组成次生保护组织(周皮)。周皮横切面的特点之一是木栓层细胞、木栓形成层细胞和栓内层细胞排列整齐,每列细胞的径向壁大致排成直线。
栓内层大多只有一层细胞,其在周皮横切面上至少有如下3个特点:1)其为形成层原始细胞第一次平周分裂的产物,为该细胞的内侧一半,内切向壁呈现弧形(因为原始细胞横切面通常呈现多边形);2)除外切向壁是新形成的壁外,其他各面保持原态,若原始细胞是厚角细胞,这些细胞壁也在角隅处加厚;3)与外方的木栓形成层细胞相比,栓内层细胞切向壁与之等宽,径向壁则与之在一条线上。
图4A[20]中被指为栓内层的细胞,其横切面半圆形,符合一般规律。但其多个细胞的径向壁并未与木栓形成层细胞在一条线上,尤其是其左起第5个细胞为从皮层插入的细胞,十分反常。此图并未显示栓内层的典型特征,不适合作为周皮模式图。
图 4B[36]被多本植物学教材[2~3,6,8~9,11,16,19,21,25,27,37]采用,可谓周皮的经典模式图。但是,图中所指的栓内层显然有误。图中被指为栓内层的细胞是椭圆形、卵形,而且与其外侧的细胞未对齐,外切向壁与其他壁一样都较厚,显然这层细胞仍是厚角组织,而不是木栓形成层平周分裂的产物。图4B中被指为木栓形成层的这层细胞大多非扁平形,完全不会是木栓形成层细胞。如果这层是栓内层呢?该层左侧第2~6个细胞具弧形内切向壁,符合栓内层细胞的特征,但右侧4个同一层的细胞具平直的内切向壁,栓内层细胞的特征不典型。此外,这层左起第2、3个细胞与其上方的两个细胞对应得很好,可推断为上方木栓形成层细胞向内平周分裂而形成;但是,这两个木栓形成层细胞向外对着1列而不是两列木栓层细胞。这种现象可解释为木栓形成层经过向外4次平周分裂产生了4个木栓层细胞;之后的第5次细胞分裂为径向垂周分裂,形成两个木栓形成层细胞;最后此2木栓形成层细胞同时平周分裂,向内产生栓内层细胞。但是,这一解释存在两个漏洞:1)这与木栓形成层原始细胞首先产生栓内层细胞的一般规律相违背;2)作为可塑性很强的初生壁,经过5次分裂后,分生组织细胞内切向壁仍保持为弧形,致使其向内产生的子细胞具有弧形且较厚的内切向壁,这种情况是不大可能存在的。退一步说,即使有这种情况,也是罕见现象,不适合当作模式。此外,图中被指为“木栓”的多层细胞含木栓形成层(图4B),称“木栓”很不恰当。
图4B还有一个问题是,周皮中外侧一部分细胞(左起第6~8列)与表皮细胞整齐排列,可由此得出“木栓形成层来自表皮细胞的脱分化”的结论。但另一些细胞(如左起第2~5列和第9列)则从表皮下第2或3层细胞起才往内整齐排列,据此又可认为“木栓形成层来自表皮细胞下的第2或第3层细胞”。进一步观察发现,右起第1~4列细胞的外侧细胞径向壁窄于内侧细胞。如果它们是木栓层细胞,应早于内部细胞形成,成熟前的生长时间更长,应更大而不应小于内部后形成的木栓层细胞。从另一方面说,这几列细胞的层数多于左侧的周皮细胞,相邻木栓层细胞有如此不同分裂速度很不寻常。如果这些细胞不是木栓层细胞,那它们为什么呈扁平状?为什么与内侧细胞排列整齐?总之,图4B疑点颇多,不适合作为周皮的模式图。
还有教材[19]对图4B有不同的标注,将图中标为栓内层的细胞标为木栓形成层,而将这层细胞内侧与之镶嵌排列而非径向成列的一层细胞标为栓内层细胞。这就错得更离谱了。
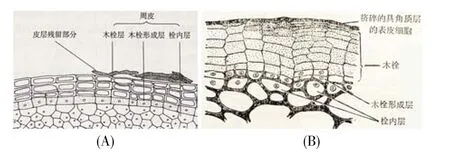
图4 周皮横切面图(A)根周皮,引自廖文波等[20];(B)茎周皮,引自伊稍[36]。Fig.4 Illustrations of periderm sections(A)A root periderm section(cited from Liao et al.[20]);(B)A periderm section of stem(cited from Esau[36]).
5 关于次生结构的探究性实验
同源器官的初生结构差异较小,而次生结构能较充分地体现植物结构对其功能的适应。反之,根据植物结构与功能相适应的原理,能深刻理解植物的结构特征。为了培养学生的观察能力、分析问题的能力,笔者设计了下面两个探究性实验。
实验1 实验材料为3根长约3 cm、直径0.2~0.3 cm的圆柱状结构。其为取自同一株葡萄的茎、卷须和果序柄。请对材料进行横切,根据结构与功能相适应的原理,判断1~3号材料各是什么器官,并阐述理由。
实验解析:1)切片、番红染色、制作临时装片,在显微镜下观察到图5显示的图像。2)根据细胞小、扁平且径向上排列整齐的特点,找到形成层区(CZ),辨别韧皮部(PP、SP)和木质部(SX、PX)。3)区分初生木质部(PX)和次生木质部(SX),进而区分导管(管腔较大)和木纤维(管腔小)。4)区分初生韧皮部(PP)和次生韧皮部(SP)。5)判断材料1(图5B)是卷须。判断理由是,其次生木质部中纤维发达,适应卷须的支持功能;次生木质部含导管少,与卷须表面积和体积之比小而蒸腾作用弱相适应;韧皮部(SP)不发达,次生生长中增加的活组织很有限,这与卷须无贮藏、大量运输有机物的功能有关。6)判断材料2(图5A)为茎。判断理由是,其次生木质部(SX)中导管较丰富,利于水分运输,因为茎上着生蒸腾作用旺盛的叶片;其韧皮部(PP和SP)较发达,利于运输有机物,这也是茎的主要功能之一。7)判断材料3(图5C)为果序柄。判断理由是,其与卷须相似,所连接的果实表面积和体积之比(比表面积)较小,蒸腾作用较弱,次生木质部(SX)缺乏运输水分的导管(不同于茎),而具较发达的纤维利于支持果实;与卷须最大的不同在于,其次生韧皮部(SP)发达,利于为果实运输有机物。

图5 葡萄茎(A)、卷须(B)和果序柄(C)的横切面(局部;自制、自拍),示维管组织CZ:形成层区;Pi:髓部;PP:初生韧皮部;PX:初生木质部;SP:次生韧皮部;SX:次生木质部。Fig.5 Transverse sections(a part,made and photographed by the author)of the stem(A),tendril(B)and peduncle of fruiting clusters(C)from a grape plant,showing vascular tissuesCZ:Cambium zone;Pi:Pith;PP:Primary phloem;PX:Primary xylem;SP:Secondary phloem;SX:Secondary xylem.
实验2 实验材料(4号)为1根长约3 cm、直径0.2~0.3 cm的圆柱状结构,取自另一物种。请对材料进行横切,根据结构与功能相适应的原理,判断其为茎、卷须和果柄中的哪类器官,阐述判断理由,并与实验1中同一类材料做比较,解释二者差异的生物学意义。
实验解析:1)切片、番红染色、制作临时装片,在显微镜下观察到图6显示的图像。2)辨别形成层(CZ)、初生木质部(PX)、次生木质部(SX)、韧皮部活组织(LPT)和韧皮纤维(PF)。3)判断4号材料为果柄。判断理由是,其次生木质部不发达,与果柄及其所连接的果实的比表面积小而蒸腾作用弱相适应;果实的支持靠发达的韧皮纤维;其韧皮部活组织发达,利于果实生长和积累有机物。4)其与葡萄的果序柄属同类结构,但适应对策有所不同,即对果实的机械支持,在葡萄中主要依靠木纤维,而4号材料[车厘子即欧洲甜樱桃(Prunus avium L.)的果柄]则主要利用韧皮纤维(PF;图6),体现了适应的多样性。

图6 车厘子果柄徒手切片横切面(局部;自制、自拍)CZ:形成层区;LPT:韧皮部活组织;PF:韧皮纤维;Pi:髓部;PX:初生木质部;SX:次生木质部。Fig.6 A hand-cut transverse section(a part,made and photographed by the author)of a fruit stalk of Prunus avium L.CZ:Cambium zone;LPT:Living phloem tissues;PF:Phloem fibers;Pi:Pith;PX:Primary xylem;SX:Secondary xylem.