丁酸通过诱导自噬抑制宫颈癌细胞增殖的研究
2022-11-05郭君萍张珂季夏薇玉环吴芳全施迪邦张嘉珉王方岩
郭君萍 张珂 季夏薇 玉环 吴芳全 施迪邦 张嘉珉 王方岩
宫颈癌是全球女性的第四大常见恶性肿瘤,也是癌症相关死亡的第四大原因,严重威胁女性的健康[1]。在中国,宫颈癌患者早期诊断率低,临床上现行的手术、放疗、化疗等治疗手段效果也不理想。目前,宫颈癌的发生、发展机制仍未完全阐明,深入探索其内在机制可能提供新的诊疗思路。近年来越来越多研究发现自噬在宫颈癌发生、发展中的重要作用[2-3]。自噬是指细胞通过溶酶体的作用对自身多余的蛋白质和受损的细胞器进行清除,维持细胞内稳态[4]。然而,过度的自噬会导致细胞自噬性死亡[5]。诱导过度自噬已成为部分宫颈癌治疗药物的作用靶点,例如二甲双胍联合奈非那韦的抗肿瘤作用与诱导宫颈癌细胞中的自噬体形成有关[6],一些合成的抗肿瘤肽如UM-6通过诱导细胞自噬在体外抑制宫颈癌细胞[7]。丁酸是肠道微生物发酵膳食纤维产生的具有生物活性的短链脂肪酸,不仅能提供肠上皮细胞70%的能量,还能抑制宫颈癌、肝癌、结直肠癌、肺癌等多种肿瘤的发生、发展[8-11]。有研究表明,丁酸可通过诱导活性氧(reactive oxygen species,ROS)的积累,抑制Akt/mTOR途径,从而促进肝癌细胞自噬蛋白表达和自噬体形成[12]。有关宫颈癌的一些研究发现,丁酸能够显著抑制肿瘤细胞的增殖,但是否通过自噬以及其确切的机制还有待阐明。基于此,本研究利用离体培养的宫颈癌Hela细胞系,通过检测自噬相关蛋白,探讨丁酸抑制宫颈癌的具体作用机制,现报道如下。
1 材料和方法
1.1 材料
1.1.1 主要试剂 丁酸钠(纯度>98%)购自中国Aladdin公司,溶于DMEM培养基(中国Gibco公司)中配制成100 mmol/L原液,并用0.22 μm过滤膜过滤,保存在-20℃环境中。3-甲基腺嘌呤(3-Methyladenine,3-MA)购自美国MCE公司。MTT细胞增殖及细胞毒性检测试剂盒(型号:C0009S)购自中国Beyotime公司。细胞计数试剂盒(cell counting Kit-8,CCK-8)购自日本同仁化学研究所。微管相关蛋白1轻链3(microtubule-associated protein 1 light chain 3,LC3)抗体购买于美国CST公司,p62抗体购买于中国Affinity公司,甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphatedehydrogenase,GAPDH)抗体、山羊二抗购自中国HUABIO公司,所有抗体均以1∶1 000比例稀释,保存在4℃环境中。
1.1.2 细胞 人类宫颈癌Hela细胞购于美国ATCC公司,以含10%FBS的DMEM培养液培养,置于含5%CO2、37℃培养箱(德国Eppendorf AG公司,型号:6731KN019434)中培养。根据细胞生长情况换液,细胞生长密度为70%~80%时进行传代。
1.2 方法
1.2.1 MTT检测细胞增殖能力 将处于生长对数期的Hela细胞按2 000个/孔加入96孔板,培养24 h使细胞贴壁,加入5 mmol/L的丁酸钠培养12、24、48 h(丁酸处理组),对照组不作处理。MTT粉末溶于稀释液中配成5 mg/ml的工作液,-20℃避光保存。每孔加入10 μl的MTT工作液,在培养箱中孵育4 h,再加入100 μl的Formazan溶解液,混合均匀,37℃孵育3 h至紫色结晶全部溶解,使用多功能酶标仪(美国ThermoScientific公司,型号:VarioskanLUX)在570 nm测定吸光度。
1.2.2 透射电子显微镜观察细胞超微结构 将培养48 h的丁酸处理组细胞与对照组细胞分别收集到1.5 ml EP管中,2.5%戊二醛固定24 h以上,再使用四氧化锇固定细胞30 min,梯度乙醇溶液脱水,包埋,半薄切片,超薄切片。最后,在透射电子显微镜(日本HITACHI公司,型号:H7500)下观察自噬囊泡、自噬溶酶体的形成,并进行拍照。
1.2.3 mGFP-RFP-LC3双荧光实验检测细胞自噬流将Hela细胞接种在12孔板的细胞爬片上,24 h完全贴壁后,每孔加入脂质体Lip2000(美国Invitrogen公司)转染mGFP-RFP-LC3质粒(中国麟美生物公司科技有限公司)1 μg,培养8 h后,5 mmol/L丁酸钠分别处理12、24、48 h,对照组不作处理。弃培养液,PBS清洗,加入4%多聚甲醛溶液室温固定15 min,重复PBS清洗过程,0.1%Triton X-100处理5 min后再次PBS清洗,1%FBS室温固定30 min,含DAPI抗荧光淬灭封片液封片后在荧光显微镜(日本尼康公司,型号:CI-L)下观察细胞自噬流。
1.2.4 Western blot法检测细胞自噬相关蛋白LC3、p62表达 将Hela细胞接种到6孔板,5 mmol/L丁酸钠处理48 h后收集(丁酸处理组),对照组不作处理,加入RAPI裂解液和蛋白酶抑制剂提取蛋白。使用二甲胺基丙酸蛋白质测定试剂盒检测蛋白质浓度,12%十二烷基硫酸钠聚丙烯酰胺凝胶电泳分离细胞蛋白质,并转移到聚偏二氟乙烯膜(BIO-RAD)中,5%脱脂奶粉室温封闭1.5 h,含吐温的Tris缓冲盐溶液(Tris buffered saline with Tween,TBST)清洗3次,敷一抗4℃摇床过夜。第2天回收一抗,TBST清洗3次,室温孵育二抗2 h。使用化学发光成像系统(美国GE公司)扫描,Image J软件通过密度测定法对蛋白质条带进行定量分析。
1.2.5 CCK-8法检测自噬抑制剂3-MA对细胞的半抑制浓度(IC50) 将处于生长对数期的Hela细胞按3 000个/孔接种到96孔板中,完全贴壁后加入含有不同浓度3-MA的培养基在细胞培养箱里培养48 h,弃去培养基,每孔加入100 μl含10%CCK-8的混合液,37℃培养箱孵育2 h,用多功能酶标仪在450 nm波长处检测吸光度,计算细胞活力,细胞活力=(A实验组-A空白)/(A对照组-A空白)。
1.2.6 实时荧光定量PCR(real-time quantitative polymerase chain reaction,RT-qPCR)法检测细胞LC3、p62 mRNA表达 用5 mmol/L丁酸钠处理Hela细胞12、24、48 h后收集细胞(丁酸处理组),对照组不作处理。Trizol试剂提取总RNA,使用基因扩增核糖核酸聚合酶链反应试剂盒进行逆转录,加入特异性引物(生工生物工程股份有限公司)、SYBR聚合酶(翌圣生物科技股份有限公司),在高通量荧光定量PCR仪(新加坡ABI公司)上进行RT-qPCR。数据采用2-ΔΔCt方法进行分析。引物序列见表1。

表1 引物序列
1.2.7 流式细胞术检测细胞线粒体ROS水平 使用线粒体超氧化物荧光探针检测细胞线粒体中的ROS相对表达水平。将细胞培养在6孔板中,分为对照组、丁酸组(加入5 mmol/L丁酸钠处理48 h)、3-MA+丁酸组(3-MA加5 mmol/L丁酸钠作用48 h)。加入PBS配制的5 μmol/L ROS工作液,于37℃避光孵育10 min,收集细胞线粒体ROS水平检测细胞线粒体ROS水平。
1.3 统计学处理 采用SPSS 21.0统计软件。计量资料多组比较采用单因素方差分析,组间两两比较采用LSD-t检验,两组比较采用两独立样本t检验。P<0.05为差异有统计学意义。
2 结果
2.1 丁酸处理组与对照组细胞增殖能力比较 细胞培养12、24 h时,两组细胞增殖能力比较差异均无统计学意义(均P>0.05);培养48 h后,与对照组相比,丁酸处理组细胞增殖能力明显减弱(P<0.05),即丁酸抑制Hela细胞增殖,在48 h时最显著,见图1。
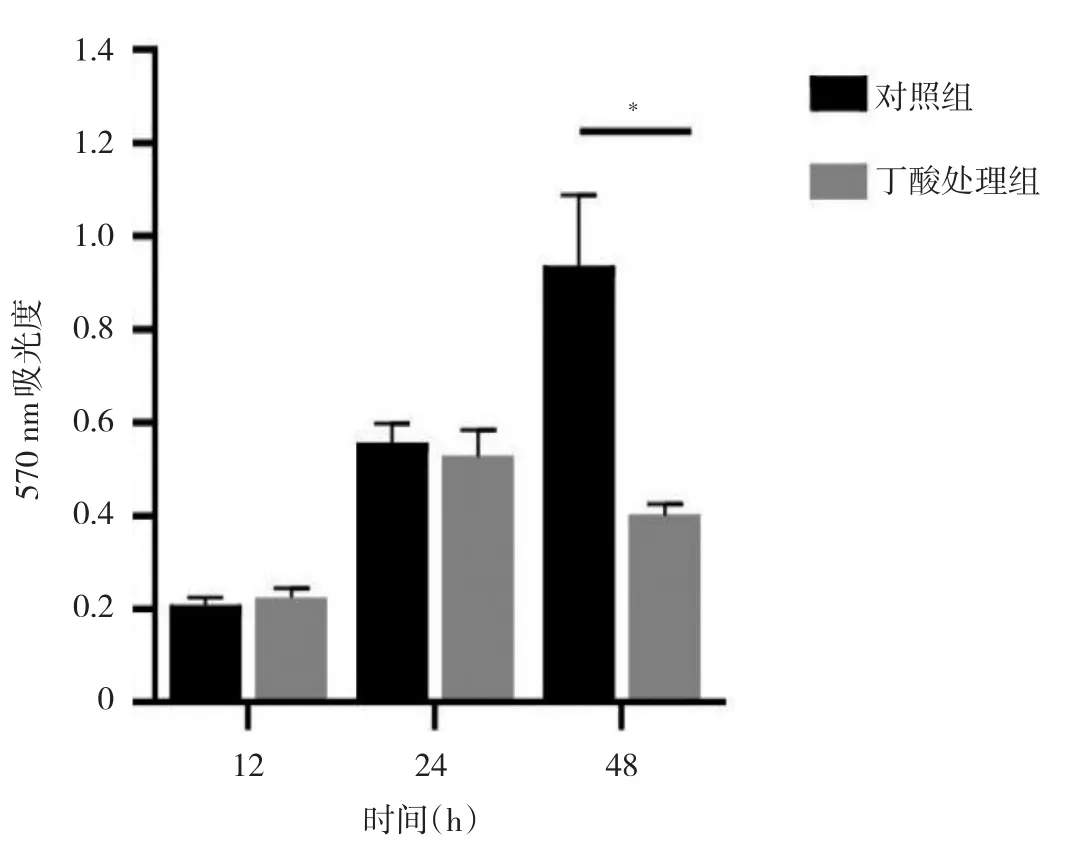
图1 丁酸处理组与对照组细胞增殖能力比较
2.2 丁酸处理组与对照组细胞超微结构观察所见对照组细胞中可观察到正常的细胞核和核膜,线粒体、内质网等细胞器结构清晰,无水肿、空泡样改变,见图2a、b。丁酸处理组细胞中可观察到大量双层膜结构的自噬体,囊泡内包含受损的线粒体等细胞器,呈典型的自噬改变,见图2c、d。即丁酸可促进宫颈癌细胞自噬。

图2 丁酸处理组与对照组细胞超微结构观察所见(a、b:对照组;c、d:丁酸处理组;a、c:×15 000;b、d:×30 000)
2.3 丁酸处理组与对照组细胞自噬流观察 LC3被红色荧光和绿色荧光标记,当自噬小体与溶酶体结合成自噬溶酶体时,绿色荧光在酸性环境下发生淬灭,只能检测到红色荧光。随着丁酸处理时间增加,绿色荧光逐渐减弱,红色荧光逐渐增强,即丁酸促进自噬体向自噬溶酶体的转化增多,自噬流水平升高,见图3(插页)。
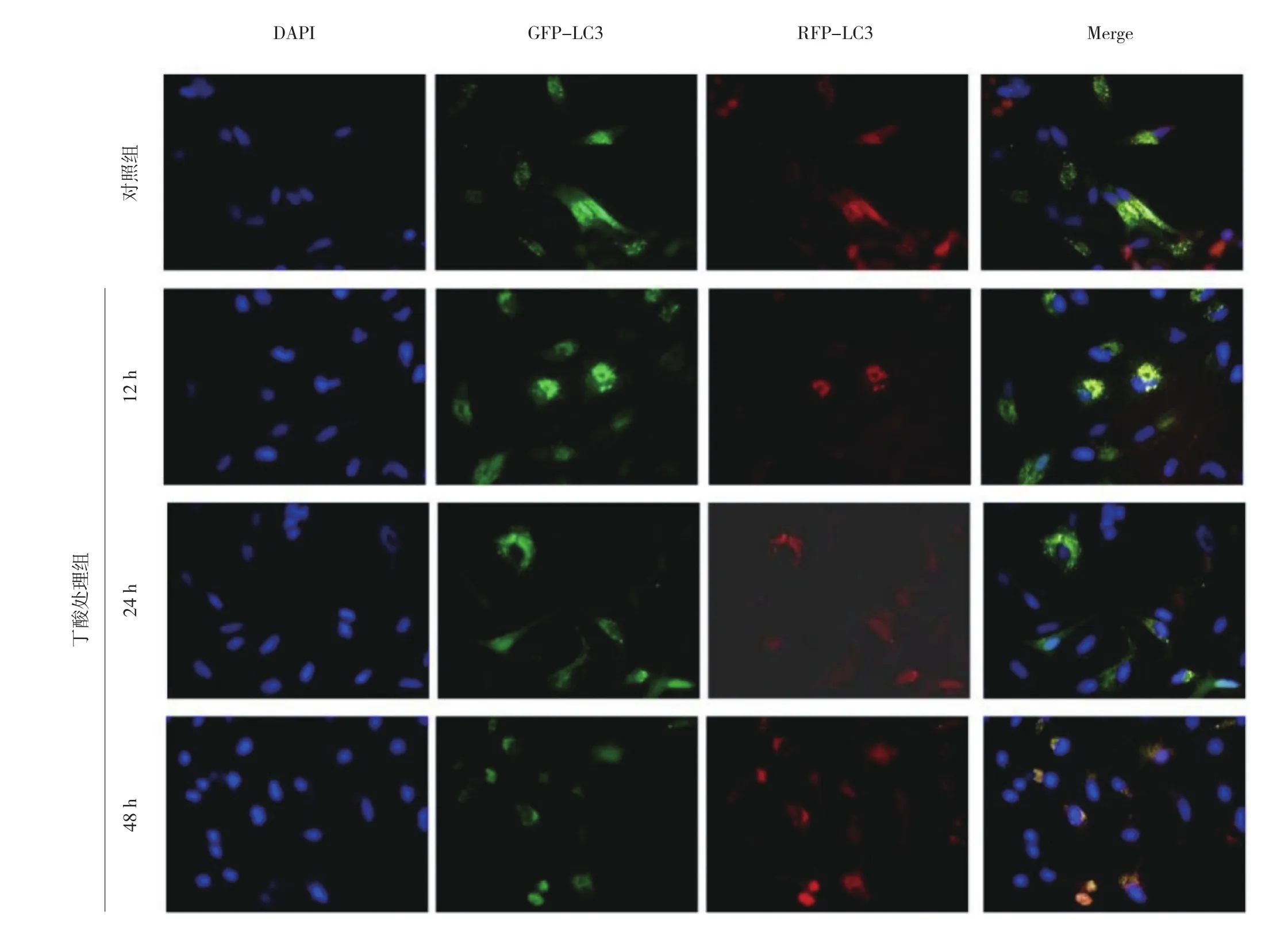
图3 丁酸处理组与对照组细胞自噬流观察所见
2.4 丁酸处理组与对照组细胞LC3、p62蛋白和mRNA表达水平比较 不管是蛋白水平还是mRNA水平,与对照组相比,丁酸处理组随着丁酸处理时间增加,LC3-Ⅰ向LC3-Ⅱ的转化增多,p62的表达则随着处理时间增加而减少,见图4。即丁酸诱导Hela细胞自噬。
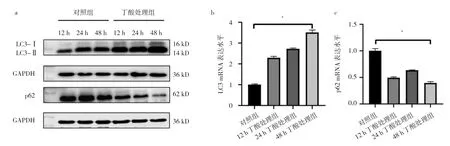
图4 丁酸处理组与对照组细胞LC3、p62蛋白和mRNA表达水平比较(a:LC3、p62蛋白表达水平比较电泳图;b:LC3 mRNA表达水平比较;c:p62 mRNA表达水平比较)
2.5 3-MA对细胞的IC50检测及对细胞自噬的影响随着3-MA浓度增加,3-MA对细胞的抑制作用也增强。本研究选择5 mmol/L作为3-MA的工作浓度,它抑制作用较强,毒性较小。Western blot结果表明,丁酸诱导的LC3-Ⅱ活化可被3-MA抑制,即丁酸对宫颈癌细胞自噬起促进作用,见图5。

图5 3-MA对细胞的IC50检测及对细胞自噬的影响(a:不同浓度3-MA作用于细胞的IC50检测;b:LC3蛋白表达电泳图)
2.6 对照组、丁酸组、3-MA+丁酸组细胞线粒体ROS水平比较 流式细胞术显示,与对照组相比,丁酸组ROS水平升高(P<0.05),3-MA+丁酸组ROS水平低于丁酸组但高于对照组(均P<0.05),见图6。即丁酸诱导自噬可能通过线粒体途径,而ROS在其中发挥重要作用。
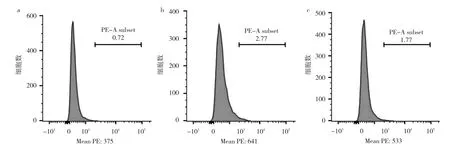
图6 对照组、丁酸组、3-MA+丁酸组细胞线粒体ROS水平比较流式细胞图(a:对照组;b:丁酸处理组;c:3-MA+丁酸组)
3 讨论
丁酸是生物活性最活跃的短链脂肪酸,可以通过多种机制发挥抗炎和抗肿瘤作用。作为组蛋白乙酰化抑制剂,丁酸增加组蛋白乙酰化水平,调控增殖、凋亡、代谢等基因表达,发挥抗肿瘤作用[13-14]。此外,丁酸还能结合GPR41、43、109a抑制肿瘤的发生、发展[15-16]。本研究结果显示,丁酸显著抑制宫颈癌细胞的增殖,且这种抑制作用与细胞自噬有关。
细胞自噬可以通过消除受损的细胞器,回收代谢产物调节内平衡,促进癌细胞存活。研究表明,抑制自噬可以缓解卵巢癌[17-18]、结直肠癌[19]、肺癌[20]等的化疗药物耐药性。在胰腺癌细胞中,SMAD4基因敲除引发的放射治疗抵抗力与ROS积累、辐射诱导的自噬水平升高有关[21]。自噬也被证明在头颈部鳞状细胞癌中保护肿瘤细胞免受放射治疗引起的细胞损伤[22]。此外,自噬阻断剂氯喹被报道可增强IL-2免疫治疗的抗肿瘤作用[23-24]。然而过度的自噬会诱发细胞程序性死亡,自噬性死亡已成为抑制癌症的重要手段[25]。在乳腺癌细胞中,氟苯达唑通过诱导细胞自噬性死亡抑制细胞增殖和迁移[26]。自噬介导的细胞死亡也被发现与前列腺凋亡反应4蛋白抑制恶性胶质瘤细胞有关[27]。丁酸作为不同肿瘤治疗的潜力药物,其抗肿瘤作用也被报道与细胞自噬有关。小剂量的丁酸和槲皮素联合应用可通过抑制保护性自噬增加胶质母细胞瘤的凋亡[28]。大剂量的丁酸则通过促进自噬性死亡抑制肾细胞瘤增殖[29]。
本研究结果表明,丁酸能显著抑制宫颈癌细胞的增殖,进一步通过透射电子显微镜观察到丁酸处理后的宫颈癌细胞中出现大量吞噬了破损的线粒体等细胞器的自噬体,呈典型的自噬变化。因此,笔者推测细胞自噬是丁酸抑制宫颈癌细胞增殖的关键机制。自噬作为真核细胞内保守的一种进化形式,首先是在内质网或者其他细胞膜上形成吞噬泡以响应应激信号通路,接着抗胸腺细胞球蛋白5(antithymocyte globulin,ATG5)和ATG12耦联形成复合体,LC3被半胱氨酸蛋白酶ATG4切割生成LC3-Ⅰ,后者被ATG7激活后,在ATG3的作用下与磷脂酰乙醇胺结合转化为LC3-Ⅱ,促进自噬体膜扩增[30]。不断增长的吞噬膜通过膜上的LC3-Ⅱ与细胞器、蛋白质上的分子相互作用促进其选择性摄取与降解,而p62是其中重要的衔接蛋白[31]。最后自噬泡与溶酶体结合并被溶酶体酶降解。在本研究中,mRFP-GFP-LC3双荧光实验显示绿色荧光减弱,红色荧光增强,提示自噬流增加,RT-qPCR和蛋白印记分析结果均表明LC3-Ⅰ转化为LC3-Ⅱ增多,p62表达减少,以上结果均提示丁酸激活自噬。使用自噬抑制剂3-MA可以抑制丁酸诱导的LC3-Ⅱ活化,进一步证明丁酸确实促进了宫颈癌细胞自噬。
自噬过程常常伴随ROS水平的升高,进一步研究发现ROS作为信号分子,能触发细胞自噬过程的发生。甘草酸被报道通过ROS-线粒体途径诱导乳腺癌细胞自噬与死亡,在抑制癌细胞生长中起重要作用[32]。异黄酮产生ROS破坏线粒体,导致自噬并诱导凋亡,显著抑制肿瘤细胞[33]。在骨肉瘤细胞中,芬太尼诱导ROS生成,磷酸化c-Jun N端激酶(c-Jun N-terminal kinase,JNK),从而触发自噬性死亡发挥抗肿瘤作用[34]。本研究也观察到自噬体吞噬了受损的线粒体。因此,本研究检测了宫颈癌细胞中线粒体的ROS水平,发现丁酸促进ROS在线粒体内的积累,并且这种ROS的表达增多能够被3-MA抑制,提示丁酸可能通过线粒体途径诱导宫颈癌细胞自噬。
综上所述,丁酸通过诱导自噬抑制宫颈癌细胞增殖,而线粒体ROS可能是其重要的触发机制。
