木薯二倍体及其同源四倍体叶片形态、生理及抗螨特征比较
2022-10-29韦婉羚黄珍玲陈会鲜杨海霞梁振华李恒锐
韦婉羚 黄珍玲 陈会鲜 杨海霞 兰 秀 梁振华 李恒锐
(广西南亚热带农业科学研究所,广西 龙州 532415)
木薯(ManihotesculentaCrantz)是热带、亚热带地区重要的粮食、饲料作物,也是重要的工业原料,深加工产品多达3 000种,涉及国计民生的各个领域[1]。朱砂叶螨(Tetranychuscinnabarinus)是重要的世界危险性农业害螨,目前已经成为为害木薯最严重的有害生物之一,可使木薯减产20%~70%,甚至绝收[2-3]。目前,各木薯产区对于朱砂叶螨的防治仍依赖于化学药剂,但存在喷药难、农药有效利用率低、使用频率与剂量不断加大和“3R”等严重问题,因此,寻求符合环保要求的新防治策略和方法成为我国现代木薯产业发展中亟待解决的重要课题[4]。选育具有高抗朱砂叶螨的优质高产木薯新品种具有重要的生物学及经济学意义。
植物多倍体通常具有营养生长优势和更强的环境适应性,因而多倍体育种是植物遗传改良的重要途径[5]。多倍体植物由于染色体加倍,具有器官巨大、可孕性低、生活能力强、内含物多、抗逆性强、耐贮运等特点,目前多倍体育种技术已经受到广泛关注与深入研究[6]。Chao等[7]和Liu等[8]的研究分别证明了多倍体拟南芥、四倍体大麦抗盐性强于其二倍体;Ruiz等[9]研究了四倍体卡里佐枳橙的抗硼过量能力,证明其四倍体抗性比二倍体更强。可知在非生物逆境胁迫下,多倍体植物的抗逆性普遍高于二倍体。在生物胁迫方面,姬万丽[10]和张弛[11]分别研究了四倍体西瓜枯萎病抗性和红阳猕猴桃溃疡病抗性,结果均表明四倍体植株抗病性比二倍体更强。由此可见,多倍体优良的抗逆性表现已经被充分证实。多倍体化是提高植物抗逆性的有效手段。
近年来国内开展了木薯多倍体诱导研究。安飞飞等[12]研究发现,华南8号四倍体植株叶片较其二倍体厚,且叶色深绿,块根干物率和粗淀粉含量均较其二倍体显著降低。李恒锐等[13]研究表明,四倍体木薯的开花结实物候期均迟于二倍体,且四倍体的雌花、雄花、花药、果实、种子的形态大小均大于二倍体。凡杰[14]对获得的四倍体植株与二倍体供体种进行蛋白质组学研究,发现木薯加倍后,与胁迫以及信号传导相关的差异表达蛋白质表达水平上调,并据此推测多倍体的抗性会提高。魏望[15]研究发现,低温胁迫下,四倍体木薯叶片遭受低温损伤和叶片萎蔫下垂更加严重,且叶片黄化面积和数目均高于二倍体,四倍体木薯的抗寒性弱于其二倍体。尚小红等[16]研究发现,在干旱胁迫下,四倍体木薯的总抗氧化能力强于二倍体,能产生更多的超氧化物歧化酶以清除活性氧,减少植株体内过氧化氢对细胞的破坏,四倍体受到干旱胁迫的影响明显小于其二倍体。李迁等[17]对国家木薯种质资源圃227份核心木薯种质进行抗朱砂叶螨的抗性评价,发现华南124和华南205多倍体植株的抗螨性均强于其二倍体。但目前关于木薯同源四倍体在抗朱砂叶螨方面的深入研究仍鲜见报道。因此,本研究通过对木薯品种华南205、南植199的二倍体及其同源四倍体植株的叶片形态结构和逆境生理指标进行测定比较,并观察自然感螨情况下植株的田间表现,鉴定其抗螨性,分析其抗性与叶片形态结构和逆境生理指标间的相关性,探究木薯同源四倍体抗朱砂叶螨的优势机理,以期为木薯抗螨性育种提供科学指导。
1 材料与方法
1.1 试验材料
供试材料为广西主栽木薯品种南植199(NZ199)和华南205(SC205)的二倍体及其同源四倍体成熟种茎。其中二倍体(2x)材料(染色体数为2n=2x=36条),来自广西南亚热带农业科学研究所木薯种质资源圃;四倍体(4x)材料(染色体数为2n=4x=72条),由广西农业科学院经济作物研究所严华兵课题组鉴定后提供。
1.2 试验地概况
试验地位于广西壮族自治区崇左市龙州县境内的广西南亚热带农业科学研究所木薯试验基地(22°20′16″N,106°47′19″E),海拔138 m左右。试验地属南亚热带季风气候,年均气温≥23.1℃,年降雨量≥1 266 mm,年日照时数约1 521 h,年均相对湿度约88%,全年无霜期361 d。
1.3 试验方法
1.3.1 试验设计 木薯于2021年3月21日种植,试验地为壤土,肥力中等。木薯种植株行距为0.8 m×1.0 m,每小区种植25株,采用完全随机区组设计,每个品种为1个处理,3次重复。种植前施用有机肥 12 000 kg·hm-2作为基肥。种后50 d进行第1次追肥,施尿素112.5 kg·hm-2,复合肥(N∶P∶K为15∶15∶15)150 kg·hm-2,氯化钾112.5 kg·hm-2。种后120 d进行第2次追肥,施尿素225 kg·hm-2,复合肥(N∶P∶K为15∶15∶15)300 kg·hm-2,氯化钾300 kg·hm-2,其他栽培管理与常规管理一致。自然感螨,不喷施农药。于2021年8月16日(木薯朱砂叶螨为害高峰期)调查木薯叶片受朱砂叶螨为害情况,随后观测各小区叶片形态及中裂片长宽,并采集植株顶芽下第5片完全展开的未受害叶片,用于叶片显微结构观测和逆境生理指标测定。
1.3.2 叶片形态及中裂片长宽观测 每小区随机选取15片叶观测叶片裂片数、中裂片形状、中裂片叶缘、顶端形状,以及中裂片长与宽,取平均值。中裂片长为叶基部至叶尖的距离,中裂片宽为裂叶最宽处叶缘之间的距离。
1.3.3 叶片显微解剖结构的观测 叶片横切面组织结构:于中裂片中脉两侧剪取约0.5 cm2的叶片块,用甲醛-乙酸-乙醇(formaldehyde-acetic acid-ethanol,FAA)固定液进行固定、常规石蜡切片法切片,切片厚度为12 μm,番红-固绿对染,制成永久切片,显微镜镜检合格后使用Eclipse Ci-L拍照显微镜(Nikon,日本)选取组织的目的区域进行100倍和400倍成像。成像完成后使用Image-Pro Plus 6.0分析软件,以mm作为标准单位,观测并计算叶片厚度、下表皮厚度、栅栏组织厚度、海绵组织厚度,计算叶片组织结构紧密度(cell tense ratio,CTR)、组织结构疏松度(spongy ratio,SR)和栅海比:
栅海比=栅栏组织厚度/海绵组织厚度
(1)
CTR=栅栏组织厚度/叶片厚度×100%
(2)
SR=海绵组织厚度/叶片厚度×100%
(3)
每项指标均测定50个数值,求平均值。
气孔特征:于中裂片中脉两侧剪取约0.5 cm2的叶片块,用2.5%戊二醛固定,后经脱水、干燥,再进行导电处理,最后使用SU8100扫描电子显微镜(Hitachi,日本)对样本进行观察并采集图像。应用Image-Pro Plus 6.0软件测量计算每平方毫米气孔密度、气孔长度和宽度。
1.3.4 叶片逆境生理指标及测定 丙二醛(malondialdehyde, MDA)含量、单宁含量、多酚氧化酶(polyphenol oxidase, PPO)活性、苯丙氨酸解氨酶(phenylanine ammonialyase, PAL)活性均参照试剂盒说明书测定,试剂盒均购自苏州格锐思生物科技有限公司。
1.3.5 田间鉴定方法 木薯种质抗螨性鉴定评价方法及评级标准参照《NY/T 2445-2013 木薯种质资源抗虫性鉴定技术规程》[18]。
采用五点取样法进行调查:从木薯植株上、中、下3个部位的东、南、西、北4个方位各随机选1片叶片为代表,每株共调查12片叶,调查受害程度,根据叶片受害症状确定和记录受害级别(表1),计算螨害指数。随后,根据螨害指数将木薯种质的抗螨性分为免疫、高抗、抗、中抗、感和高感共6级(表2)。

表1 木薯叶片受朱砂叶螨为害的分级标准

表2 木薯种质对朱砂叶螨的田间抗螨性鉴定评级标准
螨害指数计算公式:
(4)
式中,I表示螨害指数%;S为叶片受害级别;NS为该受害级别叶片数;N为调查总叶片数。
1.4 统计分析
数据采用Excel 2013进行处理和制表,用SPSS 20.0进行统计分析,采取单因素方差分析和Duncan’s检验法进行差异显著性分析,Pearson法进行相关性分析。
2 结果与分析
2.1 叶片形态组织特征比较
2.1.1 叶片形态特征比较 由图1可知,染色体经过加倍后,叶片裂叶数、叶缘类型不变,但叶片裂叶的形状有所改变。华南205二倍体叶片的中裂叶形状为线形,其同源四倍体为提琴形,两种倍性叶片的裂叶数均为9片;南植199二倍体叶片的中裂叶形状为披针形,而其同源四倍体为倒卵形,两种倍性叶片的裂叶数均为7片;两品种的叶顶端形状均由二倍体的渐尖变成四倍体的尾尖,叶片整体更宽更短。
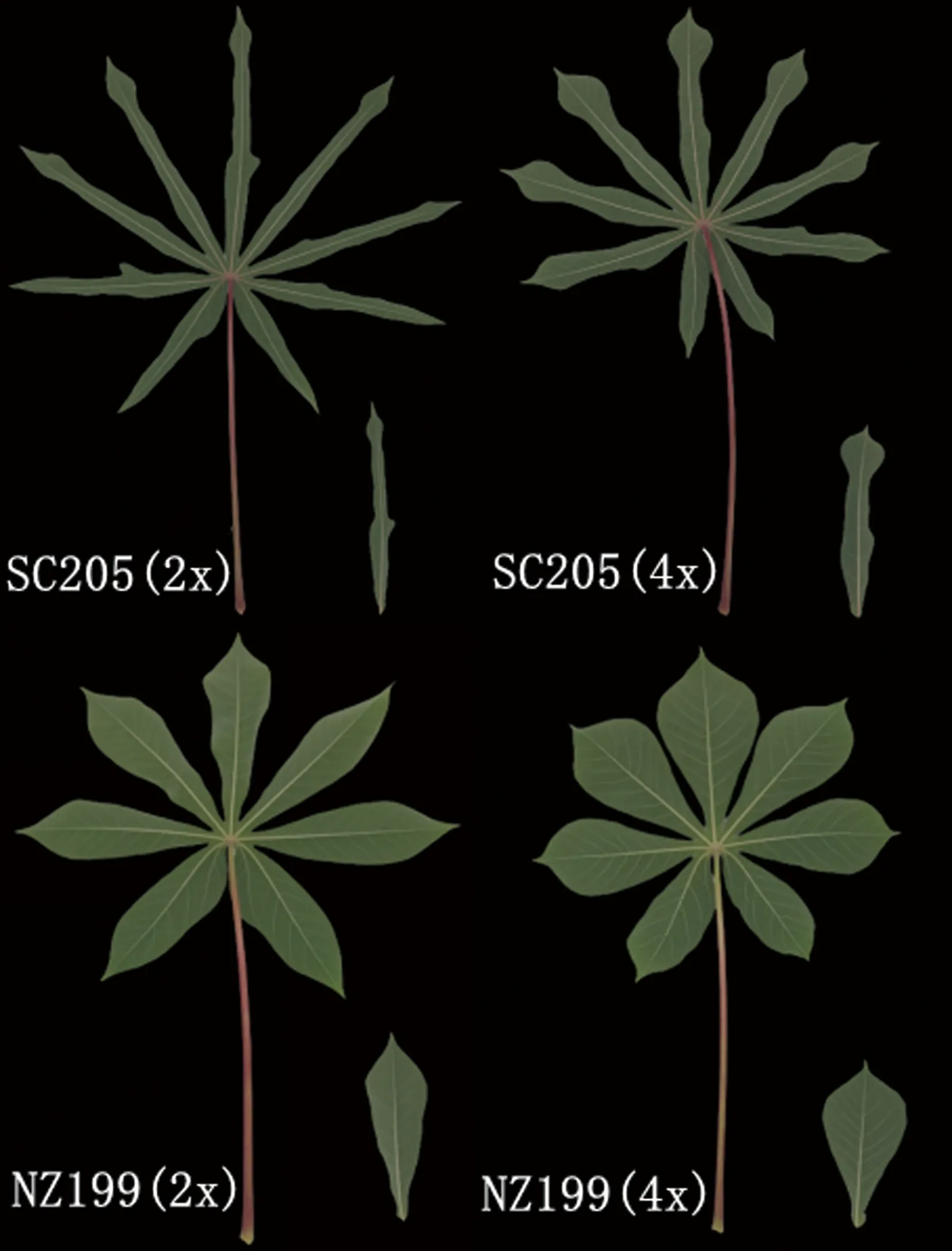
图1 二倍体及其同源四倍体木薯叶片比较
方差分析结果表明(表3),两木薯品种的二倍体及其四倍体植株叶片的中裂叶长、宽、叶形指数均差异显著(P<0.05)。华南205同源四倍体、南植199的同源四倍体中裂叶长分别是其二倍体的0.81倍和0.87倍,中裂叶宽分别是1.46倍和1.28倍,叶形指数分别是0.55倍和0.68倍。可见,随着倍数的增加,木薯叶长和叶形指数显著变小,而叶宽显著变大。
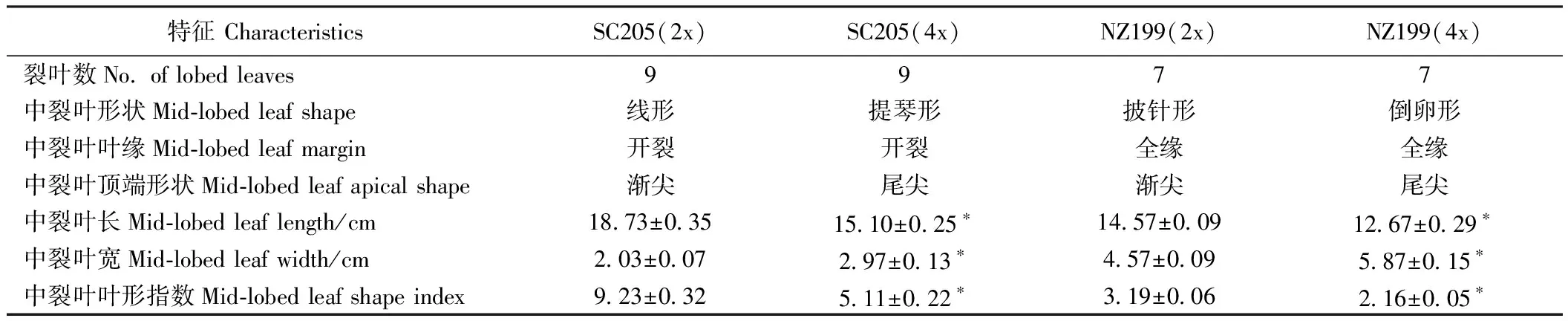
表3 二倍体及其同源四倍体木薯叶片特征比较
2.1.2 叶片解剖结构比较 由图2可知,不同木薯叶片细胞排列顺序基本相同,栅栏组织、海绵组织明显分化。栅栏组织由多层柱状细胞组成,每层细胞排列整齐、紧密,近等长。海绵组织由排列疏松的圆形或椭圆形细胞组成。由表4可知,华南205同源四倍体、南植199同源四倍体的叶片厚度分别是其二倍体的1.17倍和1.14倍,栅栏组织厚度分别是1.15倍和1.25倍,海绵组织厚度分别是1.09倍和1.06倍,下表皮厚度分别是1.14倍和1.22倍,下角质层分别是1.11倍和1.12倍,栅海比分别是1.05倍和1.18倍。两品种的叶片厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度、下角质层厚度均随倍性增大而增大,其同源四倍体与二倍体间差异显著。华南205同源四倍体、南植199同源四倍体的组织结构疏松度分别是其二倍体的0.91倍和0.92倍,组织结构疏松度随倍性的增大而变小;而组织结构紧密度变化无规律,两品种的二倍体及其同源四倍体间差异不显著。
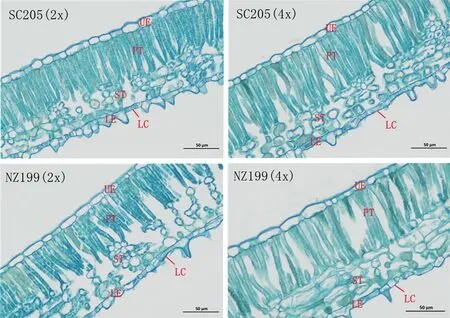
注:UE:上表皮;LE:下表皮;PT:栅栏组织;ST:海绵组织;LC:下角质层。
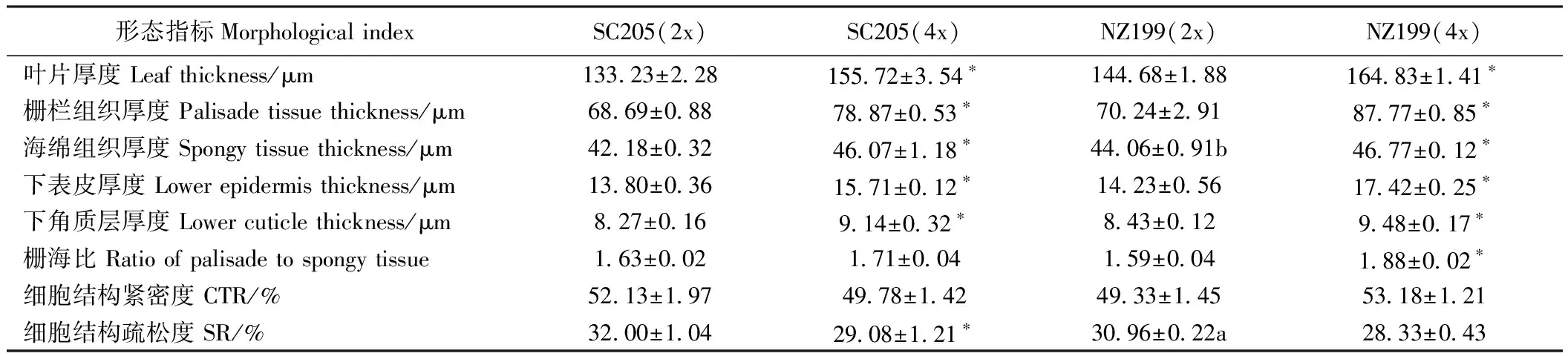
表4 二倍体及其同源四倍体木薯叶片横切面解剖结构比较
2.1.3 气孔特征比较 由表5可知,华南205同源四倍体、南植199同源四倍体的气孔长度分别是其二倍体的1.29倍和1.58倍,气孔宽度分别是0.99倍和0.75倍,气孔密度分别是0.76倍和0.57倍。同源四倍体气孔长度显著大于其二倍体,同源四倍体气孔宽度小于其二倍体但差异不显著,但同源四倍体的气孔密度显著低于其二倍体。可见,同源四倍体气孔增大,气孔密度比其二倍性植株小(图3)。
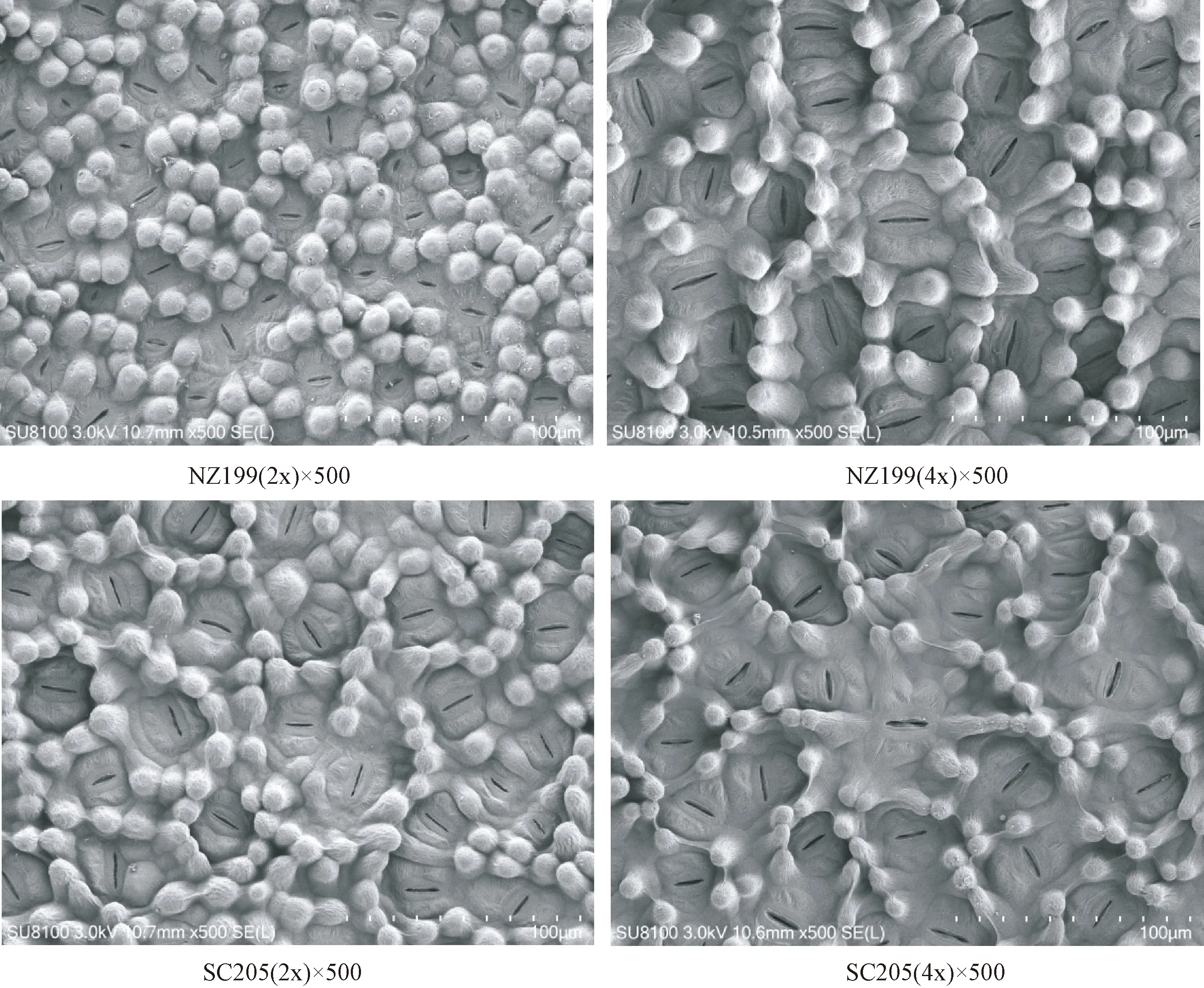
图3 二倍体及其同源四倍体木薯叶片气孔特征显微示意图
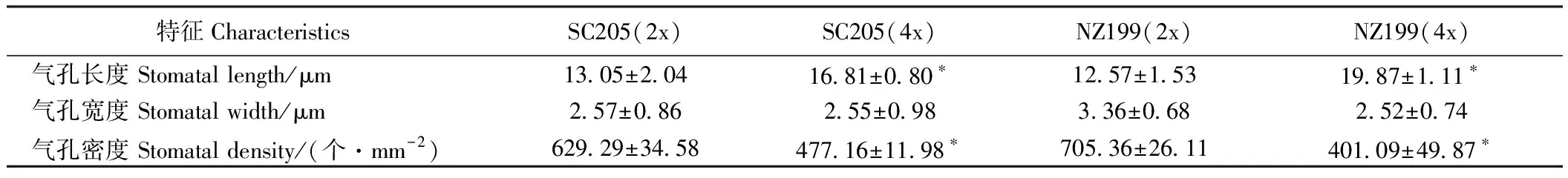
表5 二倍体及其同源四倍体木薯叶片气孔特征比较
2.2 叶片抗逆生理指标比较
由表6可知,华南205同源四倍体、南植199同源四倍体的丙二醛含量分别是其二倍体的0.92倍和0.89倍,单宁含量分别是1.04倍和1.04倍,多酚氧化酶活性分别是1.29倍和1.13倍,苯丙氨酸解氨酶活性分别是1.30倍和1.73倍。两品种的同源四倍体丙二醛含量较二倍体降低,而单宁含量和苯丙氨酸解氨酶活性显著升高,华南205同源四倍体多酚氧化酶活性显著高于其二倍体,而南植199同源四倍体高于其二倍体,但差异不显著。
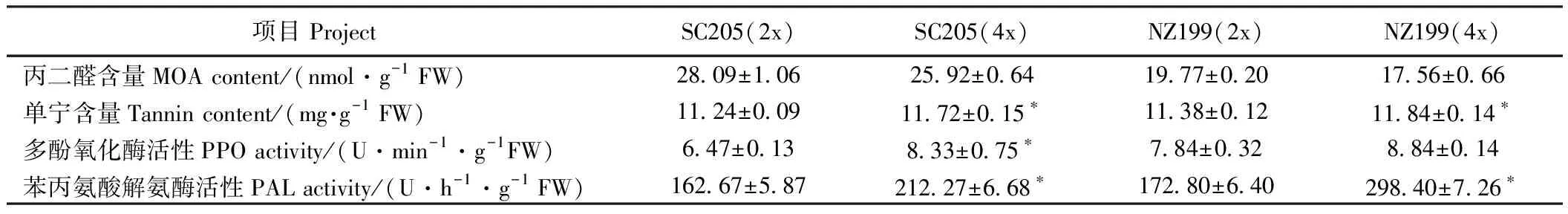
表6 二倍体及其同源四倍体木薯叶片逆境生理指标
2.3 木薯对朱砂叶螨抗性比较
由图4可知,华南205二倍体叶面黄色斑点最多且成片,受害最严重,而南植199同源四倍体叶面黄色小斑点最少,受害轻微。结合表7可知,华南205同源四倍体螨害指数较其二倍体下降39.5个百分点,南植199同源四倍体螨害指数较其二倍体下降16.7个百分点。木薯品种华南205、南植199同源四倍体受螨害程度均较其二倍体低(图4),抗螨性由强到弱排序为:南植199同源四倍体(抗)>华南205同源四倍体(中抗)>南植199二倍体(中抗)>华南205二倍体(感)。木薯同源四倍体的抗螨性均强于其二倍体。

图4 二倍体及其同源四倍体木薯叶片受朱砂叶螨为害的田间表现
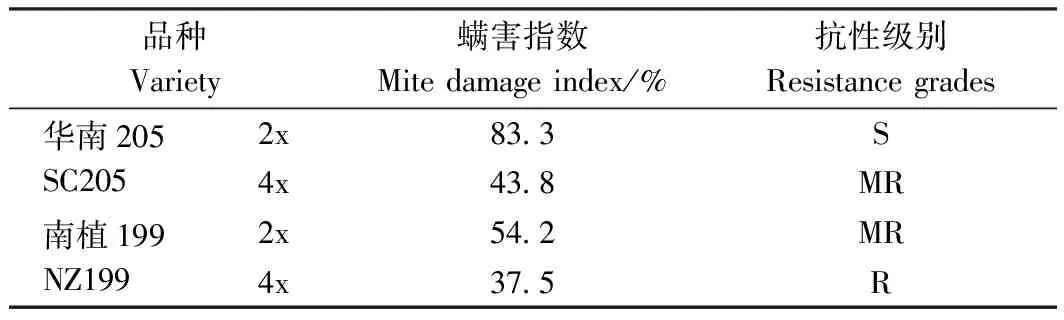
表7 二倍体及其同源四倍体木薯对朱砂叶螨的田间抗性鉴定结果
2.4 木薯抗螨性与叶片形态结构特征和逆境生理指标的相关性分析
对不同倍性木薯螨害指数与部分叶片形态结构特征和逆境生理指标进行相关性分析(表8),结果表明,螨害指数与叶片厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度、下角质层厚度、多酚氧化酶活性以及苯丙氨酸解氨酶活性呈极显著负相关,与单宁含量呈显著负相关;与气孔密度、丙二醛含量呈显著正相关。
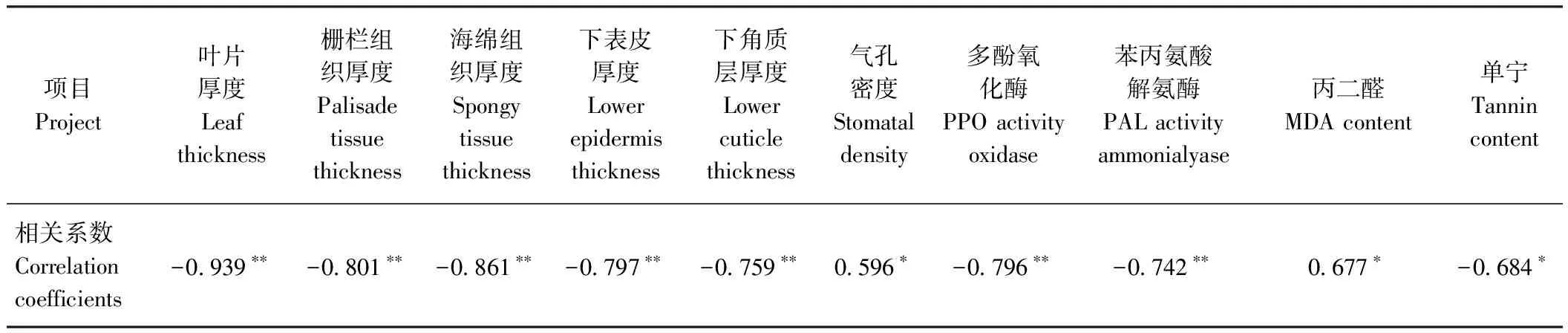
表8 木薯螨害指数与叶片形态结构特征和逆境生理指标的相关性分析
3 讨论
3.1 二倍体及其同源四倍体木薯叶片形态结构特征
不同倍性植株的表型存在差异,主要体现在细胞学、形态学及生理生化等方面[19]。本试验结果表明,与二倍体相比,同源四倍体叶片更加厚实,小叶叶长更短、叶宽更宽,叶形指数显著变小(表3),气孔增大,气孔密度变小(表5),该试验结果与魏望[15]和周慧文等[20]的研究结果一致。同源四倍体叶片大,叶片与光的接触面积增加,这可能是同源四倍体植株生长更健壮的重要原因。Skorupska[21]研究发现,气孔密度与抗螨性呈显著负相关。气孔作为害螨取食的天然孔道,分布越密,害螨越易于取食,植株抗性越弱。木薯同源四倍体植株的气孔大、气孔密度小,不易于害螨取食,抗性强。叶片的显微结构可以反映植物对环境的适应能力,是植物抗逆综合评定必不可少的一部分。研究表明,下表皮、海绵组织和栅栏组织厚度等组织结构与抗螨性呈显著正相关[22-23]。本研究结果表明,同源四倍体叶片栅栏组织厚度、海绵组织厚度、下表皮厚度、下角质层厚度、栅海比均比其二倍体大(表4)。害螨多群集于叶背,其取食的叶绿体主要存在于栅栏组织中,害螨口针需经过下表皮和海绵组织才能到达栅栏组织,所以下表皮和海绵组织越厚越不利于害螨取食。由此推测,染色体加倍后木薯叶片厚度、下表皮厚度等物理性状的改变,可能会直接导致其抗螨性发生变化。
3.2 二倍体及其同源四倍体木薯叶片抗逆境生理指标特征
丙二醛是膜脂过氧化作用的主要产物之一。丙二醛含量在细胞内的积累常会导致膜脂过氧化,渗透系统受到破坏,因而其含量高低与抗逆性密切相关[24-25]。关于黑皮冬瓜[25]、蓖麻[26]的研究表明,四倍体植株丙二醛含量低于其二倍体。次生代谢物质在昆虫营养上无重要意义,但能显著影响昆虫取食、生长和繁殖。单宁与植物抗虫关系密切,被公认为是在作物防御害虫取食中发挥直接作用的次生代谢物质,是植物抗虫的重要化学屏障[27-28]。Ropek等[29]发现抗虫蚕豆品种能够有效抵御蚕豆蚜为害,与其叶组织中单宁含量含有较高有关。吴岩等[24]认为木薯对木瓜秀粉蚧的抗性与叶组织中总酚、单宁含量显著正相关,与丙二醛含量显著负相关。本试验结果表明,木薯同源四倍体丙二醛含量显著低于其二倍体(表6)。说明同源四倍体中丙二醛含量维持在较低水平,在逆境胁迫下受到的损伤较轻,抗逆性强。木薯同源四倍体叶片单宁含量显著高于其二倍体,由此推测取食同源四倍体可能对害螨生长发育和种群数量扩大的抑制作用更强,因而同源四倍体植株受害更轻,抗性更强。
苯丙氨酸解氨酶和多酚氧化酶活性与植物抗虫性密切相关。苯丙氨酸解氨酶可促进木质素的合成,木质素含量的增加可阻碍害虫进一步取食危害,从而达到抵抗虫害的目的[30]。多酚氧化酶可将酚类物质氧化为毒性更强的醌类物质,醌类物质对昆虫的生长发育繁殖有明显的抑制作用,是一种重要的抗性生理指标[31]。尹艳等[32]研究发现,四倍体头花蓼苯丙氨酸解氨酶、查尔酮异构酶、过氧化氢酶、超氧化物歧化酶活性均显著高于二倍体。王宇[33]发现小麦高抗蚜品种的PAL和PPO活性均显著高于其他抗性品种。本试验发现,同源四倍体植株叶片的多酚氧化酶以及苯丙氨酸解氨酶活性均比其二倍体高(表6)。说明同源四倍体催化次生代谢物质(酚类、醌类等)合成的能力更强,同源四倍体具有更强大的化学防御物质基础,从而不利于害螨生存,有利于减少害螨种群数量,减轻植株受害程度。
3.3 二倍体及其同源四倍体木薯抗螨性特征
大量研究表明,植物的抗虫性与叶片厚度等物理性状及单宁、丙二醛等化学物质含量息息相关,如螨害指数与叶片厚度、下表皮和栅栏组织厚度呈显著负相关,与气孔密度呈显著正相关[21-23];抗螨种质叶片苯丙氨酸解氨酶和多酚氧化酶活性[34-35]、单宁含量[36-37]显著高于感螨种质;丙二醛含量与叶片为害指数呈显著正相关[38]。本研究中,抗螨性鉴定发现,木薯同源四倍体的抗螨性均强于其二倍体(表7、图4)。相关性分析结果表明(表8),抗螨性越强的木薯品种,其叶片厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度和下角质层厚度以及叶片单宁含量、多酚氧化酶活性和苯丙氨酸解氨酶活性越大,气孔密度和丙二醛含量越小。上述结果说明,木薯的物理屏障和化学物质含量对朱砂叶螨寄主选择性具有重要影响。由此推测,木薯同源四倍体的抗螨性比其二倍体强的原因,可能是由于基因的剂量效应,增加了同源四倍体木薯固有的物理防御和化学防御机制,并直接影响了朱砂叶螨的敏感性和行为表现,从而减少了来自朱砂叶螨取食的侵害,增加植物在环境中的生存能力。
4 结论
本研究以华南205、南植199的二倍体及其同源四倍体为试验材料,通过形态结构、抗逆生理指标和抗螨性的比较及相关性分析,发现同源四倍体在抗螨性、叶片形态结构和生理指标方面均优于其二倍体。作为一种新的种质资源和育种方向,同源四倍体木薯具有较高的利用价值和前景。
