水位影响泥炭沼泽土壤有机碳分解的生物化学机制研究进展
2022-10-27徐志伟辛沐蓉王钰婷刘莎莎王升忠
徐志伟,辛沐蓉,王钰婷,刘莎莎,王升忠,*
1 东北师范大学地理科学学院,长白山地理过程与生态安全教育部重点实验室,长春 130024 2 东北师范大学泥炭沼泽研究所,长白山湿地与生态吉林省联合重点实验室,长春 130024 3 东北师范大学,国家环境保护湿地生态与植被恢复重点实验室,长春 130024
政府气候变化专门委员会(IPCC)报告称,北纬地区10年内平均代际气温增幅为0.6℃,是全球平均水平的两倍[1]。随着气温的升高,降水量及其变异性、干旱频率及其严重程度以及蒸散量预计也会增加[1]。另有研究表明,由于气候变化和农业、林业及泥炭开采的需要,温带、北方地区及热带地区泥炭沼泽水位开始呈现下降趋势,而排水将会使泥炭地水位下降10—50 cm[2],并已经导致全球10%—15%的泥炭沼泽退化[3]。研究表明,排水退化泥炭沼泽温室气体(GHG)排放量约占全球人为GHG排放量的5%[4—5],而为了在2100年前将全球变暖幅度控制在1.5—2℃,这一比值将会达到12%—41%[6]。随着对泥炭沼泽碳储量的关注不断增加,人们越来越重视退化泥炭沼泽的复湿工作[7]。退化泥炭沼泽的复湿过程可以立即有效减少或阻止净碳损失[5],甚至增加碳汇功能[8]。根据《巴黎气候协定》2050—2070年实现“碳中和”的目标,预计需要约500000 km2的退化泥炭沼泽需要进行复湿工作,即平均每年超过10000 km2[9]。2021—2030年联合国生态系统恢复十年计划必须迎接这一挑战。
泥炭沼泽占陆地表面积的3%,但其碳储量估计在530—1055 Gt之间[10—12],约占全球陆地土壤碳储量的30%[11]。泥炭沼泽对气候变化及人类活动的响应最为敏感,在维持全球碳平衡及调节区域气候中发挥着重要的开关作用。水文条件是泥炭沼泽生态系统最重要的环境要素,影响其碳输入和输出之间的平衡作用,决定着碳循环各个关键过程的作用机制与强度[13]。自然泥炭沼泽的滞水条件限制植物残体很难被微生物完全分解[14],进而积累了大量的有机碳(SOC),使其成为大气 CO2的吸收“汇”和 CH4的排放“源”[15]。然而,水位下降加速了土壤SOC的快速分解[16—18],降低其碳储量,并可能进一步影响全球气候变暖进程[19]。长期排水作用会改变地表植被类型,使得形成泥炭的物质主要来源由藓类植物被灌木/乔木所取代[20—21],并直接或者间接地影响植被对CO2的固定与释放过程,影响生态系统SOC的输入[22—23]。此外,SOC分解改变了泥炭的水理性质,导致容重增加、孔隙度和水力传导率降低、地下水位波动更大,这势必会影响泥炭沼泽SOC的存储与分解过程,并使得严重退化泥炭沼泽的复湿可能在几十年内也无法恢复其自然状态下的生态功能[24]。有研究指出复湿后CH4排放量甚至高于其自然泥炭沼[25]。可见,关于退化泥炭沼泽及其复湿过程中土壤碳排放过程存在争议[26],这意味着未来泥炭地的碳排放存在相当大的不确定性[27—28]。因此,关于退化泥炭沼泽及其复湿过程中碳循环过程及其变化机制的研究,是当前全球变化生态学及陆地生态系统碳循环关键过程研究中的前沿性科学问题。
微生物是最丰富的土壤生物,强烈依赖于水分,它们通过水与环境相互作用、获取资源和繁衍[29]。微生物通过两种重要但截然不同的方式控制土壤碳循环过程,一方面微生物释放的酶是生态系统SOC分解的重要功能性物质[30—31],在木质素降解、腐殖质化、土壤SOC矿化分解等关键生态过程中发挥着重要作用[32];另一方面,微生物将SOC以更为稳定的微生物残体的形式存储在碳库中,进而促进SOC的存储[33]。土壤胞外酶活性越来越多地应用于预测气候变化影响的土壤碳循环模型[34—35]。然而,有研究表明SOC的氧化分解和CO2的产生是由于非酶或非生物反应,亦或是生物-非生物机制的耦合过程[36]。研究人员将SOC的分解归因于铁和锰等活性金属中间产物的存在,它们介导了分子氧对大分子SOC的氧化,并已被证明增强了农田[37]、森林[38]和湿地[39]等不同生态系统土壤SOC的氧化分解。目前,有关泥炭沼泽土壤微生物在SOC中扮演的角色及SOC分解的生物或非生物调控机制尚不明确。因此,有必要对现有研究结果进行归纳和总结,以便于更好地把握全球变化背景下泥炭沼泽土壤碳循环过程。
对土壤碳-气候反馈的未来预测依赖于对控制SOC分解机制的准确理解以及它们如何对气候变化作出反应。因此,厘清泥炭沼泽水位、铁化学、酶活性和SOC分解之间相互作用机制,对于预测生态系统-气候/泥炭沼泽水位管理反馈作用具有重要意义。本文重点综述了干旱/排水/再湿对泥炭沼泽土壤SOC组分、分子结构、碳排放的影响,并从酶活性、铁化学及微生物角度归纳总结了泥炭沼泽SOC分解的生物化学机制,将有助于更好地预测未来气候变化背景下泥炭沼泽土壤SOC储量的变化趋势,为今后深入开展泥炭沼泽土壤SOC分解机制的相关研究工作奠定基础,并为制定提高退化泥炭地生态系统碳“汇”功能的政策措施提供科学参考。
1 水位对泥炭沼泽SOC循环的影响
1.1 对土壤SOC组分的影响
长期排水作用下泥炭沼泽表层泥炭的好氧分解导致SOC的质量下降,土壤中易分解SOC比例降低而难分解SOC比例增加[40]。通过对雨养泥炭沼泽和森林沼泽的研究发现,长期排水作用显著降低了底物可利用性和枯落物质量[22,31],限制了土壤微生物的生长,进而导致土壤微生物碳含量降低[41]。研究指出泥炭地排水,导致水位下降,迅速地引起土壤SOC氧化分解,增加了溶解性有机碳输出生态系统[42],导致泥炭地碳储量降低。现有研究表明,水位波动、干旱、含氧量、温度和植被变化是影响泥炭沼泽酚类物质累积的主要因素[43]。排水/干旱作用会通过增强酚氧化酶活性而促进酚类物质的分解,进而降低泥炭沼泽土壤中酚类物质的含量[44]。然而,近来也有研究发现水位降低通过增加维管束植物来增加泥炭沼泽枯落物和孔隙水中酚类物质含量[17]。可见,泥炭沼泽酚类物质对环境变化响应具有复杂性[45],其对水位变化的响应还取决于持续时间及土地利用方式等因素。
1.2 对土壤SOC分子结构的影响

1.3 对土壤SOC分解的影响
水位控制着土壤地表的有氧层厚度,是调节进出泥炭沼泽碳通量过程的关键调节因子[52]。因此,泥炭沼泽碳循环对由于气候变化/人类活动导致的水位变化十分敏感。排水会通过改变土壤理化性质直接影响SOC分解过程。曝气环境会加速SOC的分解,使得泥炭沼泽成为碳“源”[16—18]。早期研究表明,水位下降或者排水后,由于植被类型、气候条件及水位变化的差异,泥炭沼泽土壤CO2排放通量将会增加13%—200%[53—55],而土壤CH4排放通量将会减少25%—100%[56],甚至成为大气CH4吸收汇[57]。研究表明,在60 cm土壤深度内,土壤CO2排放通量随着水位埋深的增加而增加[58]。然而,排水后泥炭孔隙度、pH、温度的降低及干旱胁迫对SOC的分解有抑制作用[58]。有证据表明,排水仅会导致富营养泥炭地碳的损失,而对贫营养泥炭地土壤碳排放几乎没有影响,甚至降低了CO2排放[59—60]。泥炭沼泽中灯芯草等植物具有发达的通气组织,这可能会使其在排水后仍旧是CH4的微弱排放源[61]。此外,植被类型的变化也会影响土壤碳排放对排水的响应。研究发现,泥炭沼泽排水造林后,其较高的总初级生产力抵消了CO2排放量的成倍增加,使其仍旧表现为CO2的吸收“汇”[62]。
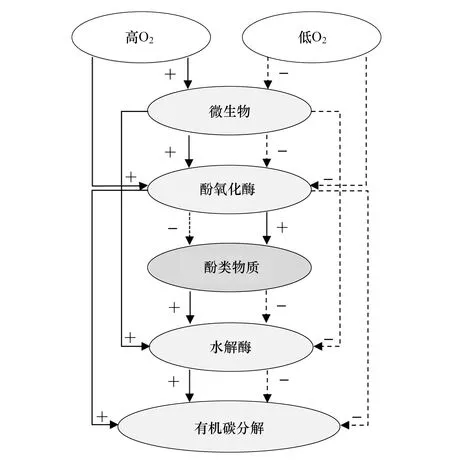
图1 氧在土壤有机碳分解过程中的作用的示意图 Fig.1 Schematic diagram of the role of O2 in soil organic carbon decomposition process右侧虚线箭头表示“酶锁”机制中的低氧作用。左侧实线箭头表示增加氧的作用,表示“酶锁”打开; “+”符号表示刺激(积极)作用,而“-”符号表示抑制(消极)作用(改编自参考文献[16])
在退化泥炭沼泽水位恢复过程中,水位的升高降低了CO2排放,但只有在持续淹水恢复条件下土壤CO2排放量才出现显著降低,而周期性淹水恢复对CO2排放量并没有显著影响[63]。然而,也有研究表明水位恢复12年后,泥炭沼泽仍处于CO2净排放状态,并占全年温室气体总排放量的33%,其主要原因是凋落物输入的激发效应促进了SOC分解,而地上凋落物的覆盖则降低了总初级生产量[64]。同时,研究表明退化泥炭沼泽水位恢复并不一定能够促进CH4的排放[65]。这一方面是因为恢复初期阶段土壤可利用性养分含量较低,限制了产甲烷菌微生物的活性[66]。另一方面,CH4只有在稳定厌氧条件下才能产生,而恢复的初期阶段土壤结构、容重等土壤物理性并没有恢复[66],泥炭沼泽不能有效保持水位,缺乏CH4生成的必要环境条件。此外,产甲烷菌可以直接利用Fe(III)作为电子受体,所以在Fe(III)氧化物存在的条件下,产甲烷菌能够以Fe(III)作为电子受体,进而抑制CH4的产生[67]。综上,退化泥炭沼泽及其水位恢复过程中泥炭沼泽SOC分解过程在不同地点和研究中高度不一致[68],进而增加了泥炭沼泽碳循环对未来气候变化响应的不确定性,并对“缺氧是维持泥炭地碳存储的关键”的传统理论提出了质疑,其主要原因是针对退化泥炭沼泽及其恢复过程中土壤SOC分解机制尚不明晰。
2 泥炭沼泽土壤SOC分解的生物化学机制
2.1 “酶锁”机制
土壤氧化酶被认为是影响泥炭沼泽碳循环的关键。然而,泥炭沼泽较低的pH[69]、温度[70]及含氧量[71]环境抑制了酚氧化酶活性和酚类物质的分解速率,导致厌氧还原的泥炭沼泽中积累大量的酚类物质,而酚类物质对葡糖苷酶、磷酸酶和硫酸酯酶等多种土壤水解酶活性具有显著的抑制作用[16]。同时,酚类物质可通过减少微生物可利用基质[72]、限制微生物新陈代谢[73]、降低Fe(II)含量[74]进而限制SOC分解。极低的土壤酶活性是导致沼泽环境中SOC低分解速率的重要原因,使得泥炭沼泽积累大量的SOC,这一机制被称作泥炭沼泽SOC分解的“酶锁”机制[71]。
长期以来,湿地SOC对干旱/排水的响应主要集中在O2增强胞外酶活性与SOC分解之间的正反馈[16,71](图1)。泥炭沼泽水位降低,氧含量的增加促进氧化酶活性,通过打开“酶锁”进一步刺激纤维素类化合物的降解[75]。然而,近期研究人员对这一机制提出了挑战,有关退化泥炭沼泽恢复过程中水解酶和氧化酶对水位变化的响应规律还存在不确定性。Wiedermann等[76]研究发现土壤水解酶活性随着干旱/排水作用而降低,而长期再湿的实验结果发现酚氧化酶活性在高含水量条件下更高[77],有关干旱/排水作用对土壤酚氧化酶及SOC分解的积极、中性和消极影响也均有报道[16,43,77]。同时,泥炭沼泽水位埋深及土壤湿度存在季节及年际变异,并导致氧化还原环境的变化,进而影响土壤酶活性及SOC矿化过程[77]。
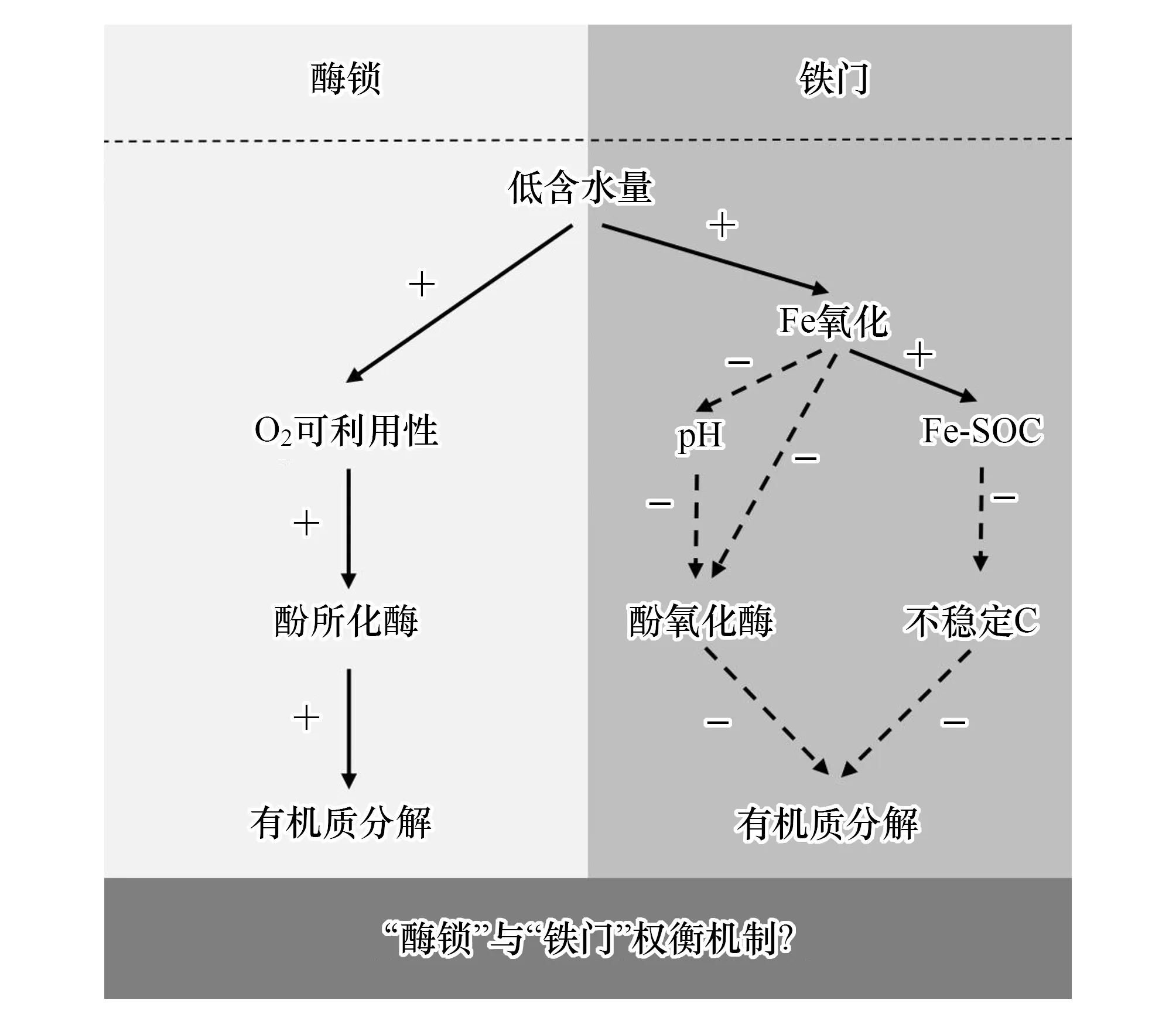
图2 泥炭地退化过程中O2和Fe在酚氧化酶及SOC分解中的作用示意图Fig.2 Schematic diagram of the roles of O2 and Fe in phenol oxidase and SOC decomposition during peatland degradationSOC:土壤有机碳Soil organic carbon, 左侧为氧在调节酚氧化酶活性和SOC分解中的主导地位,右侧为Fe(III)在酚氧化酶与SOC分解中的主导地位;“+”符号表示刺激(积极)作用,而“-”符号表示抑制(消极)作用。(改编自参考文献[77,92])
在泥炭沼泽水位变化的过程中可能存在除O2以外的其他因素控制土壤酚类物质及酶活性的变化规律。这些问题是揭示退化泥炭沼泽土壤SOC分解的“酶锁”机制的关键。随着水位的变化,除了可用O2含量的变化,还有生物群落、湿度、pH及养分可利用性的变化。Freeman等[78]研究发现水位下降带来的氧的供给的增加并非是引起酚氧化酶活性增加和酚类物质浓度变化的必然因素,土壤水分胁迫、pH及凋落物类型也具有重要作用。Williams 等[69]研究发现,泥炭中的酚氧化酶活性受曝气的影响较小,而受pH和酶抑制剂的影响较大。长期排水会使泥炭沼泽土壤pH值降低[79]。土壤酚氧化酶活性的最适pH值为近为8[32,45],这就意味着酚氧化酶活性会随着泥炭酸化而降低[45],进而抵消O2对氧化酶活性的激发作用。Xiang等[80]研究发现短期干旱引起的泥炭酸化是酚氧化酶活性降低的主要原因。此外,土壤酸化可抑制土壤脲酶、过氧化氢酶、磷酸酶等多种酶活性[81],并增加矿物吸附有机物的能力[82]。当pH值适宜时,苯酚氧化酶活性更多地取决于湿地植被类型和泥炭的植物成分,而不是气候因素。研究表明,长期排水后泥炭沼泽植物群落演替为以灌木或乔木为主的木本植物群落。木本植物凋落物会释放更多的酚类物质[83],进而抑制土壤水解酶活性。
2.2 “铁门”机制
铁和SOC通过多种途径形成Fe-SOC复合体在一定程度上对土壤有机物起到了保护作用[89]。针对不同气候变化背景下的碳循环过程的研究结果表明,Fe(III)的存在使富含矿物质的土壤和沉积物中的SOC更为稳定[90—91]。研究发现,Fe-SOC在土壤SOC中的占有重要比例,如沉积物中21.5%的SOC[90]及森林中37.8%的SOC是通过这种方式固定下来的[91]。在湿地氧化还原界面的铁氧化物对芳烃和酚类物质表现出很强的亲和力[89],在O2含量增加时会更多的保护亲水型SOC和羧基型碳不被矿化[91]。在富含有机物的泥炭沼泽中,在水位降低后,Fe(II)被氧化为较难溶的Fe(III),Fe氧化通过抑制酚氧化酶活性和促进铁-木质素结合,在抑制干旱时泥炭沼泽碳损失方面起到“铁门”的作用[77],这一机制与前述的“酶锁”机制相反(图2)。研究表明,以泥炭藓为优势植物群落的泥炭地,由于木质素含量和Fe含量低,干旱作用下土壤SOC的分解以“酶锁”机制占优势,而以维管束植物为优势群落的矿质泥炭地,由于其木质素和Fe含量高,“铁门”对SOC起保护作用[77]。“酶锁”与“铁门”机制在泥炭沼泽碳循环过程中同时发生作用,“酶锁”机制在淹水的初期阶段发挥主导作用,而“铁门”机制在淹水的后期阶段发挥作用[93],但究竟何种机制占主导作用及其背后的权衡机制仍是未知。
然而,土壤Fe在生态系统生物地球化学循环中扮演多种角色,在保护土壤碳同时也会导致碳的损失。有研究表明在富营养的泥炭沼泽中,干旱作用下铁可能通过使酚类化合物的沉淀而加速SOC的分解[93]。Fe的转化是湿地水位变化过程中一个重要的生物地球化学过程,影响土壤碳的活化和稳定[94]。土壤Fe(III)是湿地生态系统中一种重要的电子受体,它可以在厌氧环境中在微生物的作用下发生异化铁还原过程[95],且是湿地土壤SOC分解过程中最为重要的机制之一[77]。厌氧条件下,Fe(III)还原为Fe(II)并释放过氧化氢(H2O2)进而刺激Fenton化学反应,产生可溶的活性氧(如羟基自由基或超氧化物)来氧化大分子SOC[96]。此外,Fe(II)可以增强苯酚的氧化活性[32],而室内的Fe(II)添加实验也表明Fe(II)促进水解酶的活性[92],进而促进SOC的分解。Chen 等[97]通过13C标记的可溶性SOC和57FeII标记的Fe添加实验测定SOC矿化速率的研究表明Fe只有在稳定的水位状态下,且与可溶性SOC同时添加时对SOC起到保护作用。泥炭沼泽土壤尤其是表层土壤一般处于周期性的厌氧状态,所以Fe可能对SOC的保护作用是微弱的,而活性铁需要它们自身的物理化学保护特性来促进SOC持久性。此外,植物根系分泌的有机酸类物质对Fe结合的SOC起到溶解作用,进一步减弱了Fe对SOC 的保护作用[98]。因此,了解研究区的生物和地球化学背景对于理解气候变化下Fe(III)-SOC的长期命运至关重要[97]。
2.3 微生物机制
微生物是土壤SOC转化的主要驱动因素,而自然界中土壤Fe的氧化还原过程也是在微生物的介导作用下完成。北方中高纬度的泥炭沼泽,约有25%的CO2是土壤微生物代谢产生的[99]。甲烷氧化作用可以氧化湿地中产生的60%—90%的CH4,然后再逸出到大气中[100]。泥炭沼泽水位越高,其厌氧程度越大,产甲烷菌的活性就越高[101],而长期排水会减少产甲烷菌的丰度[102],当土壤再次湿润后,它们恢复得更快[103]。然而,也有研究表明在泥炭沼泽恢复过程中微生物群落的恢复需要10—15年的时间[104]。因此,在泥炭沼泽水位恢复过程中产甲烷菌并无显著变化[105],从而导致水位恢复后泥炭沼泽CH4排放量依旧较低。通过对开放水域及其周边环形湿地以及水位恢复区湿地的研究表明,甲烷排放与土壤产甲烷菌丰度无显著相关性,而产甲烷菌与甲烷氧化菌的比值和水位动态共同驱动湿地CH4的平衡[106]。水位对甲烷氧化菌的影响存在差异。Kettunen等[107]研究指出 CH4产生量在地表水位20 cm以下出现最大值,而CH4氧化能力是在水位10 cm以下出现的最大值。Sun等[108]发现,当水位在地表下20—30 cm时,湿地氧化CH4的能力大于产生能力。
一般而言,泥炭沼泽营养贫瘠,土壤产甲烷菌以氢营养产甲烷为主,但当干扰发生时,pH值和营养物质增加,产甲烷菌可能从氢营养产甲烷向乙酰分解类型转变[102]。土壤微生物对环境变化更加敏感、响应更快速。Fierer[109]研究发现土壤pH、SOC质量、氧化还原电位及土壤湿度等因素在决定土壤细菌群落结构中占有重要地位。研究表明由水位变化导致的植被及群落结构的变化能够影响土壤微生物群落的组成,进而改变湿地甲烷产生与转化的生物化学过程[110]。土壤产甲烷菌在中性到弱碱性条件下的丰度更高[111],而泥炭沼泽的酸性环境可能会抑制产甲烷菌的生长。Fe(III)是优先于CO2被利用的重要电子受体,从而能够抑制甲烷的产生,因为铁还原菌利用乙酸和氢气的浓度阈值远远低于产甲烷菌[112]。此外,产甲烷菌可以直接利用 Fe(III)作为电子受体[67,113],所以在Fe(III)氧化物存在条件下,产甲烷菌能够以Fe(III)为电子受体,从而减少甲烷的产生[114]。泥炭沼泽退化及水位恢复过程中植被群落及上述非生物因素的变化,将影响土壤微生物的群落结构、功能基因和代谢途径等。同时,由于泥炭沼泽垂直剖面微环境的差异使得不同微生物种群的增殖会产生较大的异质性。
3 总结与展望
综上,泥炭沼泽排水/再湿过程会显著改变土壤的氧化还原条件,进而改变土壤微生物群落和酶活性,驱动铁氧化还原过程,影响SOC分解速率[77]。然而,有关退化泥炭沼泽及其恢复过程中SOC分解的影响机制的认识尚不清楚,目前亟待加强的相关研究有以下几个方面:
(1)泥炭沼泽水位恢复过程中能否提高CH4排放?在排水/恢复过程中植物群落及环境因子的差异可能会影响土壤SOC分子化学组成与结构特征,而这种差异的来源及其对碳排放的影响还未可知。目前,对于自然泥炭沼泽CH4循环微生物特征已有系统研究,但退化泥炭沼泽及其恢复过程中产甲烷菌、甲烷氧化菌丰度及群落结构如何变化,退化泥炭沼泽及其恢复过程中生物及非生物因素对碳排放的贡献及其对碳排放的影响机制还不清楚。
(2)根据“酶锁”机制,排水作用会增强氧化酶和水解酶酶活性。然而近期有关排水及干旱对酶活性的影响的相反的研究结果,使得排水究竟能否打开泥炭沼泽“酶锁”存在不确定性。在泥炭沼泽水位变化过程中,有关氧化酶及水解酶活性是否会对其做出可预测的反应仍然存在争议。除O2外,是否存在其他竞争或混杂机制影响“酶锁”机制在泥炭沼泽碳循环过程中发挥作用,需要进一步探索未来气候变化背景下氧化酶-酚类物质-水解酶活性之间的作用机制。
(3)“全球铁联系”研究是国际地圈-生物圈计划 (International Geosphere- Biosphere Programme,IGBP)的三大研究主题(铁、碳、火)之一[115]。然而,针对泥炭沼泽Fe-SOC对泥炭沼泽SOC中的地位认识不足。对于铁在泥炭沼泽SOC分解中的作用还存在争议,就Fe的泥炭沼泽中的各种功能对土壤碳循环的贡献存在空白。植物根际及非根际土壤Fe的异化还原能力存在差异[116],而长期排水作用后泥炭沼泽植物群落结构发生变化,应关注由此造成的植物根系活动的差异对土壤铁的转化过程的影响。鉴于Fe(III)具有加速和抑制SOC分解过程的双重作用,应同时关注Fe的氧化和还原过程,探究酶-SOC分解/碳排放-铁之间的作用机制。未来研究将土壤水分与SOC分解的生物地球化学机制联系起来,有助于提高预测泥炭沼泽碳储存稳定性的能力,同时也为退化泥炭沼泽的恢复提供有效建议。
