粤西矿区9种先锋植物抗旱性综合评价*
2022-10-26李超楠王溢陈三雄吴龙峰王子焕
李超楠,王溢,陈三雄,吴龙峰,王子焕
(仲恺农业工程学院 园艺园林学院,广东 广州 510225)
我国石料类矿产资源丰富,是我国开采总量最大的矿产资源,约占全国矿产总产量的30%。改革开放以来,随着我国经济的飞速发展,对建筑石料需求旺盛,采石场的数量和规模得到迅速扩大,矿山数量众多。矿产资源作为国民经济重要组成部分,在推动经济发展和社会进步的同时,也带来水土流失、土地退化、生物多样性锐减、环境恶化等环境问题[1]。矿区生态恢复的目标是建立植被自我维持的生态系统,即生态系统功能恢复是一个长期的动态过程,而不是短暂的单个过程的结果[2]。植物作为生态建设的重要因素,其生长发育往往受外界环境所限制,水分限制是其中的重要因素之一[3]。许多学者研究表明,在干旱胁迫下,植物有着多样而复杂的适应和抵抗策略,包括改变自身的形态结构、生理代谢、光合效率等途径,而不同植物在面对干旱胁迫时的响应特点、适应能力和生理机制有着明显的差异[4-6]。因此,根据矿区破坏生境的地形地貌、气候、土壤等条件,选择耐旱、耐贫瘠、易成活的树种,对促进植被生态系统向自我维持的方向发展具有重要意义[7-8]。
采石场土壤受人为活动的剧烈扰动后,不仅天然表土特性丧失,而且具有极端贫瘠、持水保肥能力差、干旱或生理干旱严重等特点。植物的生长对水分亏缺较为敏感,抗旱能力较弱的植物,轻微的水分流失就能造成生长减缓,而抗旱能力强的植物会发生一系列反应,引起相关基因的表达、细胞代谢到生理反应和形态的变化从而维持生存[9]。Mehdi等[10]研究发现茶树(CamelliasinensisL.)在干旱条件下可诱导SOD、POD、CAT等酶活性的提高进而适应环境;陈简村等对4种报春苣苔属(Primulina)植物的研究发现随着干旱胁迫的加重,4种报春苣苔属植物的 POD、SOD、CAT 的活性呈上升趋势[11]。Zhou等研究入侵物种少花蒺藜草(Cenchruspauciflorus)发现,在水分亏缺情况下,长花蒺藜草的可溶性糖(SS)含量呈线性增长的趋势,可溶性蛋白在轻中度干旱胁迫下显著降低,严重干旱胁迫下则大量积累[12]。潘昕等对楝叶吴茱萸(Evodiameliaefolia)、石斑木(Rhaphiolepisindica)和任豆(Zeniainsignis)3种华南常见乔木叶片中的渗透调节物质进行研究,发现在干旱胁迫下,3 个树种的脯氨酸和可溶性糖质量比均表现出逐渐增加的趋势,在重度干旱时达到最大值,而可溶性蛋白质质量比则表现出先增大后减小的规律[13]。经过诸多学者研究表明,植物的抗旱性不是单个因子作用的结果,只有对多种因子综合评价才能够客观全面的评价植物的抗旱性水平[10-13]。本研究中对马占相思(Acaciamangium)、木荷(Schimasuperba)、黎蒴(Castanopsisfissa)、桃金娘(Rhodomyrtustomentosa)、山苍子(Litseacubeba)、银叶金合欢(Acaciapodalyriifolia)、油麻藤(Mucunasempervirens)、花叶络石(Trachelospermumjasminoides)和薜荔(Ficuspumila)9种粤西地区耐贫瘠的先锋植物进行干旱胁迫试验,通过对干旱胁迫下生理指标的测定及综合评价,筛选出抗旱性较强的植物种类,为9种先锋植物在矿区植被恢复中的配置应用及后期栽培管理提供理论依据。
1 材料与方法
1.1 试验材料
试验地点为仲恺农业工程学院海珠校区温室内进行,属于海洋性亚热带季风气候,年平均气温在20~22 ℃,年均降水量约为1 720 mm,全年中,4—6月为雨季,7—9月天气炎热,全年水热同期,利于植物生长。供试材料来源于广州德盛花木苗圃基地。试验选取长势良好且生长基本一致的马占相思、木荷、黎蒴、桃金娘、山苍子、银叶金合欢、油麻藤、花叶络石、薜荔9种植物幼苗各20株,于2021年5月进行盆栽移植,花盆直径24.5 cm,高26.5 cm,每盆栽植两株,进行编号。
1.2 试验设计
试验进行之前,对苗木进行正常的养护管理工作,待苗木定根之后,选择生长良好、长势相近的同种树苗各6盆,每盆2株,其中3盆为对照组(CK组),胁迫开始后每5 d对植物定量浇水1次,使其保持正常生长状态;另外3盆为试验组(DR组),停止供水使其维持干旱状态。在试验开始前1 d对植物进行充分灌溉,于2021年6月1日采用盆栽持续干旱方法开展试验,干旱胁迫期间每7 d(分别于6月8日、6月15日、6月22日、6月29日)进行采样,共持续4周,采取样本从上至下数第3~9片叶片,采样时间为上午的9:00—10:00。
1.3 指标测定
光合指标测定 选取每株的第3~9片叶片挂牌标记,每次采样的前1 d进行光合指标的测定,每次的测定时间为8:30—11:30和14:40—17:40,避开植物的“午休”可能对测量的影响。利用Li-6800便携式光合仪,采用开放式气路,设置光源光照强度为1 200 μmol/(m2·s),测定植物的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)等参数,所有参数8个叶片重复。
酶活及膜透性指标测定 超氧化物歧化酶(superoxide dismutase,SOD)活性采用氮蓝四唑(NBT)法;过氧化物酶(peroxide enzyme,POD)活性采用愈创木酚法;过氧化氢酶(catalase,CAT)采用钼酸铵法;丙二醛(malondialdehyde,MDA)采用硫代巴比妥酸显色法。
渗透调节物质指标测定 脯氨酸(proline,Pro)含量采用酸性茚三酮法;可溶性糖(soluble sugar,SS)含量采用蒽酮比色法;可溶性蛋白(soluble protein,SP)中采用考马斯亮蓝染色法。每项生理生化指标的测定3次重复。
1.4 抗旱性综合评价
采用模糊隶属函数法对植物进行抗旱性综合评价,隶属函数值公式为:
μ(Zi)=(Zi-Zimin)/(Zimax-Zimin)
①
μ(Zi)=1-(Zi-Zimin)/(Zimax-Zimin)
②
(i=1,2,3,...,n)
式中:μ(Zi)为各主成分的隶属函数值;为各品种第i个综合指标值;Zimax和Zimin分别为各品种第i个综合指标值的最大值和最小值。若所用指标与抗旱性呈正相关,用公式①进行计算,若所用指标与抗旱性呈负相关,则用公式②进行计算,在累加各植物材料各指标的抗旱隶属值,求其平均值。比较植物之间的抗旱隶属平均值,平均值越大,则抗旱性越强[14-15]。
1.5 数据处理
本研究试验数据利用Excel和SPSS 22.0对数据进行整理和分析,利用Origin 2018 进行图表绘制。采用单因素方差分析法分析不同干旱条件下不同植物种各试验指标之间的差异;采用隶属函数法对植物抗旱性进行综合性评价。
2 结果与分析
2.1 干旱胁迫对植物光合作用的影响
2.1.1 干旱胁迫对植物叶片净光合速率的影响
如图1(a)所示,在干旱胁迫期间9种植物叶片的净光合速率(Pn)整体呈下降趋势,均在干旱28 d时植物叶片的蒸腾速率下降至最低值,且显著低于对照组(P<0.05)。马占相思、木荷、油麻藤、花叶络石4种植物的Pn在干旱胁迫7 d时即有显著下降(P<0.05)。9种植物当中净光合速率下降幅度最大的3种植物是薜荔、银叶金合欢和马占相思,降幅分别为106.52%、104.58%和98.11%。在干旱胁迫最后时期,银叶金合欢与薜荔的净光合速率降至负值,说明此时植物的呼吸速率大于光合速率。
2.1.2 干旱胁迫对植物叶片蒸腾速率的影响
如图1(b)所示,在干旱胁迫期间9种植物叶片的蒸腾速率(Tr)持续下降,在干旱28 d时植物叶片的蒸腾速率均下降至最低值,但与对照组相比,9种植物叶片的蒸腾速率显著降低所对应时间则不同。马占相思、山苍子、银叶金合欢、油麻藤和花叶络石在干旱7 d 时即显著低于对照组,木荷和薜荔在干旱胁迫14 d时与对照组相比变化显著,桃金娘的Tr在干旱胁迫21 d时显著小于对照,黎蒴的Tr则在干旱胁迫28 d时显著小于对照(P<0.05)。9种植物中马占相思的下降比例最大,降幅为94.92%,黎蒴的下降幅度最小为35.31%。
2.1.3 干旱胁迫对植物叶片气孔导度的影响
如图1(c)所示,在干旱胁迫期间9种植物叶片的气孔导度(Gs)持续下降,均在干旱28 d时植物叶片的气孔导度下降至最低值,且除马占相思外其他植物叶片的气孔导度在干旱胁迫7 d时即与对照组相比有显著变化,马占相思在干旱胁迫14 d时的气孔导度显著小于对照组(P<0.05)。随着干旱胁迫时间的延长,马占相思与其他8种植物相比降幅较大,为95.79%,然后是桃金娘和木荷,分别下降了92.67% 和88.26% 。
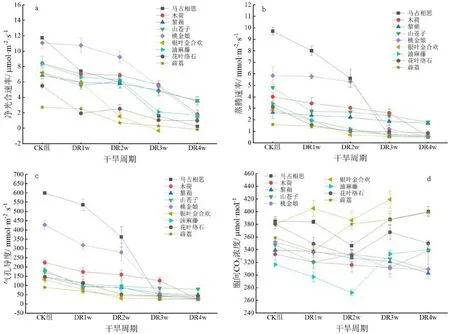
图1 干旱胁迫对植物光合作用的影响注:CK组为对照组,DR1w为干旱胁迫7 d,DR2w为干旱胁迫14 d,以此类推。下同Fig.1 Effects of drought stress on plant photosynthesis
2.1.4 干旱胁迫对植物叶片胞间CO2浓度的影响
如图1(d)所示,在干旱胁迫期间9种植物叶片的胞间CO2浓度(Ci)变化趋势不同。随着干旱胁迫时间周期的增加,马占相思、木荷、油麻藤、花叶络石和薜荔5种植物叶片的胞间CO2浓度表现为先降低后增加,但5种植物降至最低值的干旱时间不同;黎蒴、山苍子、桃金娘植物叶片的胞间CO2浓度整体上则呈现出下降的趋势,且干旱28 d时均显著低于对照组(P<0.05);银叶金合欢的叶片胞间CO2浓度随着干旱胁迫时间的延长呈上升趋势,在胁迫第14 d时略降,至第21 d时显著高于对照组(P<0.05),第28 d时植株死亡。
2.2 干旱胁迫对植物酶促保护系统及膜透性的影响
2.2.1 干旱胁迫对植物超氧化物歧化酶活性的影响
如图2(a)所示,随干旱胁迫时间的增加,9种植物的超氧化物歧化酶(SOD)活性呈现出不同的变化趋势。与对照组相比,多数植物在经过干旱胁迫后的SOD活性增加,木荷、黎蒴分别在干旱胁迫7 、14 d后SOD活性达到最大值,且与对照有显著差异(P<0.05);桃金娘、银叶金合欢(第28 d死亡)、花叶络石在干旱胁迫21 d时达到最大值,马占相思、山苍子、油麻藤、薜荔的SOD活性先降低后增加,在第28 d周干旱时上升,马占相思和山苍子第28 d干旱胁迫时SOD活性略高于对照组处理,与对照组无显著差异,油麻藤、薜荔则显著低于对照(P<0.05)。
2.2.2 干旱胁迫对植物过氧化物酶活性的影响
如图2(b)所示,9种植物的过氧化物酶(POD)活性在随着干旱胁迫周期的增加,变化趋势不同。马占相思、黎蒴、桃金娘、银叶金合欢的POD活性随着干旱周期的增加总体呈现下降的趋势,仅干旱7 d时黎蒴的POD活性显著高于对照组(P<0.05),除银叶金合欢在干旱14 d前,与对照无显著差异,另3种植物其余时间周期的POD活性均显著低于对照处理(P<0.05);木荷、油麻藤、花叶络石在干旱前期,POD活性显著增加,木荷在干旱14 d时,其POD活性达到最大值,油麻藤和花叶络石在干旱7 d的POD活性达到最大值,至干旱28 d时,木荷、油麻藤与对照相比显著降低,花叶络石无显著变化;随着干旱胁迫周期的增加,山苍子和薜荔的POD活性显著增加,且均在干旱28 d时POD活性达到峰值。
2.2.3 干旱胁迫对植物过氧化氢酶活性的影响
如图2(c)所示,随着干旱胁迫时间周期的增加,9种植物的过氧化氢酶(CAT)活性呈现出出不同的变化趋势。其中,山苍子、桃金娘、银叶金合欢、油麻藤4种植物的CAT活性在干旱胁迫期间呈现为先降低后增加的趋势,不同植物的下降幅度和下降至最低值时相应的干旱胁迫时间不同。马占相思在干旱胁迫14~21 d时,CAT活性呈下降趋势,但是在干旱28 d受到重度胁迫时,CAT活性显著增加。木荷、黎蒴、花叶络石3种植物的CAT活性整体呈现出上升的趋势,黎蒴在干旱胁迫28 d时CAT活性有所下降但变化不显著,花叶络石在干旱胁迫21 d时有暂时的显著下降(P<0.05);薜荔随着干旱胁迫时间周期的延长呈现先增加后降低的趋势。
2.2.4 干旱胁迫对植物丙二醛含量的影响
如图2(d)所示,随着干旱胁迫时间周期的延长9种植物的变化趋势有明显的不同。马占相思、黎蒴、银叶金合欢3种植物的丙二醛(MDA)含量整体上呈现不同程度的升高,马占相思在干旱胁迫7 d时的MDA含量显著降低,随后持续升高但与对照组相比无显著变化(P<0.05),黎蒴、银叶金合欢的上升幅度较大。其他6种植物随着干旱胁迫周期的延长表现出降低的趋势,不同植物下降幅度和达到最低值对应的时间不同,木荷、桃金娘、花叶络石、薜荔均在干旱7 d时,MDA含量显著下降达到最低值(P<0.05);花叶络石7 d至14 d上升趋势较大,且14 d时的MDA含量达到最大值。油麻藤在干旱胁迫14 d时的MDA含量达到最低值,山苍子在干旱28 d时MDA含量最低。

图2 干旱胁迫对植物酶促保护系统及膜透性的影响Fig.2 Effects of drought stress on the enzymatic protection system and membrane permeability of plants
2.3 干旱胁迫对植物渗透调节物质的影响
2.3.1 干旱胁迫对植物可溶性蛋白含量的影响
如图3(a)所示,与对照相比,各干旱胁迫周期下9种植物的可溶性蛋白(SP)含量整体上呈现不同程度的升高,但不同植物的SP含量达到最大值所对应的胁迫时间和增加幅度不同。马占相思、木荷、山苍子、桃金娘、油麻藤和薜荔均在干旱胁迫至第28 d时,其SP含量达到最高,且与对照组相比变化显著(P<0.05),而黎蒴、银叶金合欢、花叶络石的SP含量在干旱胁迫21 d时达到峰值,黎蒴和花叶络石在干旱胁迫28 d时SP含量下降(银叶金合欢在第28 d死亡)。木荷、黎蒴、山苍子、花叶络石、薜荔在干旱胁迫14 d时SP含量短暂降低,山苍子和薜荔的可溶性蛋白含量在干旱胁迫第21 d时略低于对照组,花叶络石在干旱胁迫第14 d时可溶性蛋白含量最低。
2.3.2 干旱胁迫对植物可溶性糖含量的影响
如图3(b)所示,马占相思、桃金娘、银叶金合欢、花叶络石、薜荔的可溶性糖(SS)含量随着干旱胁迫时间的延长整体呈现增加的趋势,但木荷、山苍子、油麻藤的可溶性糖含量则在总体上呈现降低的趋势。马占相思和薜荔在干旱胁迫7 d至21 d,其可溶性糖含量平稳上升但均低于对照组,第28 d时则显著高于对照组(P<0.05)。桃金娘、银叶金合欢、花叶络石的可溶性糖含量在干旱胁迫期间均高于对照组,桃金娘在干旱胁迫14 d时可溶性糖含量出现短暂下降,银叶金合欢和络石则在同时期可溶性糖含量达到最大值。干旱胁迫期间,木荷、油麻藤的可溶性糖含量均显著低于对照组(P<0.05),但二者的可溶性糖含量在干旱胁迫中后期无显著变化。山苍子的可溶性糖含量则在干旱胁迫7 d时上升最大至峰值,随后显著下降至第28 d时的可溶性糖含量显著低于对照组(P<0.05)。黎蒴的可溶性糖含量随干旱胁迫周期不同变化趋势不同。
2.3.3 干旱胁迫对植物脯氨酸含量的影响
如图3(c)所示,干旱胁迫对9种植物脯氨酸(Pro)含量的影响较为显著。随着干旱胁迫时间周期的增加,马占相思、木荷、黎蒴、桃金娘、银叶金合欢、油麻藤和薜荔7种植物的Pro含量整体呈上升趋势,且其他时间周期内的Pro含量均显著高于对照组(P<0.05)。马占相思和在第21 d时Pro含量大幅度增加,与对照组相比分别增加了25.72倍,至第28 d时Pro含量达到峰值。山苍子、花叶络石的Pro含量则随着干旱胁迫周期的增加而呈现降低的趋势,山苍子在干旱胁迫期间的Pro含量始终显著低于对照组,花叶络石在干旱胁迫初期Pro含量呈上升趋势,干旱胁迫14 d后则显著低于对照组(P<0.05)。



图3 干旱胁迫对植物渗透调节物质的影响Fig.3 Effects of drought stress on plant osmoregulatory substances
2.4 9种植物的抗旱性综合性评价
在逆境条件下,植物通过调动多个因子相互作用共同抵抗外在环境的影响。如果用单一的生理生化指标对植物的抗旱性强弱进行评估,得出的结论无法全面的反映出植物应对干旱环境时的综合能力。因此在对植物进行抗旱性评价时,避免由于单个指标造成的差异,往往会采用多种指标评价以提高对抗旱性树种选择的科学合理性。在本次研究中,采用了7个生理生化指标和4个光合指标对9种植物的抗旱性进行模糊隶属函数评价,计算各指标不同植物的隶属函数值,求其平均值并进行排序。隶属函数分析结果表明,在干旱胁迫21 d时(表1),9种植物的抗旱性强弱排序为:山苍子>木荷>黎蒴>花叶络石>银叶金合欢>油麻藤>桃金娘>薜荔>马占相思 ;在干旱胁迫28 d时(表2),8种植物(银叶金合欢死亡)的抗旱性强弱排序为:山苍子>花叶络石>油麻藤>木荷>黎蒴>桃金娘>薜荔>马占相思。

表1 9种植物抗旱性隶属函数综合评价结果(DR21 d)Tab.1 Results of a comprehensive evaluation of the drought resistance membership function of nine plant species (3 weeks of drought stress)

表2 9种植物抗旱性隶属函数综合评价结果(DR28 d)Tab.2 Results of a comprehensive evaluation of the drought resistance membership function of nine plant species (4 weeks of drought stress)
3 讨论与结论
植物叶片的光合系统对干旱胁迫较为敏感,光合响应相关数据通常作为衡量抗旱性的重要指标。在干旱条件下,植物气孔开度减小和气孔阻力增加以减少叶片水分的散失,同时导致CO2吸收量下降、光合速率降低[16-17]。吴雪怡等[17]通过研究表示干旱胁迫下植物的Pn均会下降,不同植物的下降幅度和敏感程度不同,下降幅度越小说明植物的抗旱性越强。在本研究中,9种植物的Pn均随干旱胁迫时间延长而下降,其中黎蒴的下降幅度最小,可能说明黎蒴的Pn受干旱胁迫的影响较小;而薜荔的Pn下降幅度最大,至干旱第28 d时,薜荔叶片的Pn已达到负值,说明其呼吸速率大于光合速率,重度干旱胁迫严重抑制了薜荔的净光合速率。研究表明,在缺水条件下,植物的叶片气孔关闭,CO2进入叶片受阻,导致植物的光合速率下降,此时,植物Ci降低,Pn降低,此阶段植物叶片光合作用主要受气孔限制影响,然而随着干旱胁迫的加剧,植物叶片的光合结构被破坏,碳的同化能力减弱,Ci升高,Pn降低,Tr和Gs均降低,此时植物叶片主要受非气孔限制影响[18-21]。本试验结果表明,黎蒴和桃金娘的Ci随干旱胁迫时间延长而不断降低,表明二者在干旱胁迫期间可能未受到非气孔限制的影响;银叶金合欢的Ci随着胁迫时间的延长而呈上升趋势,马占相思、薜荔的Ci先降低后增加,且3种植物Pn、Tr和Gs下降幅度较大,表明在光合参数评价体系中银叶金合欢、马占相思和薜荔的抗旱能力较差。
当植物受到干旱胁迫时,植物体内的活性氧(ROS)积累,引发氧化胁迫,不利于植物生长。非生物胁迫诱导抗氧化酶活性提高以响应不利的外界环境(如干旱条件),SOD、POD和CAT发挥协同作用,共同抵御干旱胁迫对植物所造成的伤害[22-23]。SOD进行歧化反应消耗过量的O2-, CAT分解歧化反应中产生的H2O2,POD清除细胞内有害的自由基并催化物质进行分解,以减少ROS对植物细胞的伤害[23-25]。Mohammad等[26]研究发现,抗旱性强的油菜(Brassicacampestris)品种中SOD、POD和CAT活性更高,具有更强地清除ROS的能力。在本研究中,花叶络石和山苍子在干旱胁迫后期,SOD、POD和CAT活性与对照组相比均有所升高,说明花叶络石和山苍子相较其他植物可以较好地清除过量的ROS以平衡氧化系统。Shahidi等[27]的研究表明,高羊茅(Festucaarundinacea)和黑麦草(Loliumperenne)在重度干旱胁迫下,CAT和SOD活性显著降低,减弱了植物清除ROS的能力。朱秀红等[28]在探究胁迫对白花泡桐(Paulowniafortunei)的生理变化时,同样表明当胁迫严重时,ROS的急剧增加超出植物的自我保护能力导致SOD、POD、CAT活性下降。本试验结果显示黎蒴和木荷在受到干旱胁迫后,3种酶活性先增加后下降,表明黎蒴、木荷在干旱初期能抵抗胁迫带来的伤害,但当干旱持续时,超过了其抗氧化酶系统的承受能力。植物处于逆境胁迫时,通常会发生膜脂过氧化作用,MDA含量能反映出膜脂过氧化程度和植物的抗逆性能力,有研究指出,抗旱性弱的植物处于逆境环境时MDA增幅较大[4,29]。本研究中,黎蒴和银叶金合欢的增幅最大,说明二者的细胞膜系统受到的伤害较为严重,除此之外的7种植物则整体上呈现出先降低后增加的趋势,表明这些植物均受到了氧化伤害,但是对氧化伤害具有一定的抗性。
渗透调节是植物响应干旱条件的一种重要机制,脯氨酸(Pro)、可溶性蛋白(SP)和可溶性糖(SS)是植物叶片渗透调节中的重要物质[30-31]。植物在受到干旱胁迫时将脯氨酸运输到细胞质中,通过增加细胞质浓度降低渗透势,从而保持细胞的渗透平衡,同时可溶性蛋白和可溶性糖通过降低细胞水势,提高植物叶片的吸水和保水能力[16]。李泽东等[32]研究表明皂荚(Gleditsiasinensis)的Pro对逆境胁迫的影响较为敏感,在干旱条件下,皂荚的Pro含量大幅增加,且植物Pro的累加程度与植物种类相关。本研究中,在干旱胁迫后期,马占相思和薜荔的Pro含量显著增长,与对照组相比分别增加了40倍、6.5倍,由此可见,在干旱胁迫严重时,脯氨酸含量在二者的抗旱性响应中发挥重要作用。此外,木荷、山苍子、桃金娘、花叶络石4种植物的Pro含量呈先增加后降低的趋势,可能是因为Pro对植物抗旱调节具有一定的限度,此时细胞的膨压无法维持,Pro含量会呈现下降的趋势[11,33]。可溶性蛋白是植物重要的渗透调节物质,在干旱胁迫下,除山苍子、薜荔外其他7种植物的SP含量总体上显著升高,与罗青红等[34]的研究结果相一致,SP含量显著上升保证了植物代谢的正常进行;山苍子、薜荔的SP含量先增加后降低。韩蕊莲等[35]研究表明,随着干旱胁迫时间的延长,植物体内的分解代谢大于合成代谢,导致可溶性蛋白降解,这可能也是本试验结果的成因。可溶性糖使生物体内重要的能源和碳源,参与植株生命代谢的许多过程[16]。潘昕等[36]研究青藏高原的6种耐旱性植物表示,抗旱性越强的植物,可溶性糖含量增加的幅度越大。本试验中,桃金娘、银叶金合欢、花叶络石在干旱胁迫各周期均显著高于对照,说明3种植物的渗透调节能力较好,且银叶金合欢的SP含量、Pro也随干旱胁迫时间显著增加,表明银叶金合欢的渗透调节能力比其他植物强。但综合所有评价指标发现,银叶金合欢的抗旱性较差,说明植物的抗旱性不是由单个因子决定的,而是多个因子相互协同作用的结果。
在干旱胁迫的过程中,植物会因胁迫时间的不同和胁迫程度的差异表现出阶段性的特征,包括适应环境、伤害、调节修复甚至再受伤害[37]。通过对9种植物在干旱胁迫下持续4个周期的光合响应指标、抗氧化酶系统和渗透调节物质等性状的研究,植物的抗旱性强弱表达并不相同。本研究也发现,利用综合分析所得到的结果与植物在干旱胁迫中的实际表现更为一致,如在渗透调节系统中银叶金合欢的抗旱性系数较高,但在干旱胁迫28 d时植株已枯萎致死,由此证明,单一的评价指标难以客观地评价植物的抗旱性。因此,用单个指标来描述某一植物的抗旱性强弱可能过于片面,植物的抗旱性强弱是由多个因子协同作用而成,需要综合分析才能科学地评价植物的抗旱能力。
综上所述,根据9种植物的干旱表现和隶属函数综合分析结果,山苍子、花叶络石和木荷相较其他植物具有更强的抗旱性,在粤西矿区进行生态修复时可选择作为恢复植被的先锋树种,并根据修复区域的干旱程度进行选择种植。在本研究结果中,对各抗旱指标的分析表明,可溶性糖含量在干旱胁迫期间波动较大,不适宜作为评价指标;光合作用参数、酶促保护系统、MDA含量、Pro含量和SP含量在一定程度上可以反映植物的抗旱性能力。
