鱼腥草抗氧化应激机制研究进展
2022-10-21杨亚军曲静苗贵东
杨亚军,曲静,苗贵东
1. 兴义民族师范学院体育学院(兴义 562400);2. 上海体育学院运动科学学院(上海 200438);3. 兴义民族师范学院生物与化学学院(兴义 562400)
鱼腥草(Houttuynia cordataThunb,HCT)属三白草科植物蕺菜,主产于日本、韩国、中国西南部。HCT作为根茎叶类蔬菜植物,药食同源,HCT及其提取物具有抗氧化[1]、抗病毒[2]、抗炎[3],抗衰老[4]、抗肿瘤[5]等作用。HCT提取物槲皮素能有效抑制Ⅱ型登革病毒[2]。HCT显著降低高糖诱导的人脐静脉内皮细胞衰老标准物的表达,发挥抗衰老作用[4],鱼腥草多糖(Houttuynia cordata polysaccharide,HCP)可减少脂多糖诱导慢性炎症大鼠肺组织炎症,改善肺水肿[2]。研究发现,HCT及其提取物的抗衰老、抗癌、抗炎作用与抗氧化有关[2,5-6],HCP能显著提高脂多糖诱导的慢性炎症大鼠细胞内抗氧化酶活性,发挥其抗氧化作用[2]。在以秀丽线虫为动物模型的抗衰老研究中发现,HCT提取物显著降低突变型线虫活性氧(reactive oxygen species,ROS),增强应激抵抗,延长其寿命[5],HCT能有效降低苯并(a)芘诱导的肺癌细胞内ROS的生成,保护细胞免受苯并(a)芘诱导的DNA损伤和炎症[6]。过氧化参与糖尿病、高血压、动脉粥样硬化等诸多临床疾病,针对抗过氧化相关反应机制的研究对治疗相应疾病具体重要意义。在概述HCT抗过氧化生理作用的基础上,综述近年来鱼腥草抗氧化应激作用机制,包括减轻体内外氧化应激反应及信号通路调控,为HCT治疗缓解氧化应激疾病的研究提供视角。
1 HCT抗氧化作用
机体依赖抗氧化系统产生超氧化物歧化酶(superoxide dismutase,SOD)、过氧化氢酶(CAtalase,CAT)、谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)、谷胱甘肽还原酶(glutathione reductase,GR),清除过量自由基,保持稳态。氧化系统和抗氧化系统失衡,引起细胞和组织内的病理变化[7]。HCT中分离鉴定出220种化合物,其主要活性成包括黄酮类、挥发油、HCP、生物碱[8],它们均可增强抗氧化酶活性,清除自由基,发挥抗氧化作用。
1.1 HCT黄酮抗氧化作用
研究发现HCT黄酮能够清除氧自由基,提高抗氧化酶活性,表现出较强的抗氧化性。细胞试验发现,HCT黄酮具有较好的抗氧化作用,HCT水提物总黄酮和多酚,能够提高小鼠成肌C2C12细胞SOD活性,逆转由肿瘤坏死因子(tumor necrosis factor-α,TNF-α)诱导的细胞活力下降[9]。动物试验发现,高剂量(160 mg/kg体重)HCT总黄酮灌胃能显著提高由四氧嘧啶诱导的Ⅰ型糖尿病小鼠血SOD含量,改善其糖耐量[10]。槲皮苷、槲皮素和金丝桃苷作为HCT黄酮的主要组成成分,能显著降低四氯甲烷诱导的急性肝中毒小鼠肝丙二醛(malonaldehyde,MDA)含量水平,提高血清谷胱甘肽转换酶(glutathione s-transferase,GST)、SOD、CAT水平[11]。此外,HCT分离出的槲皮素、槲皮素苷抑制脂质过氧化,具有明显的剂量依赖性,且糖基化的槲皮素苷比槲皮素表现出更强的抗氧化活性[12]。
1.2 HCT挥发油抗氧化应激作用
挥发油是HCT最重要的药效成分之一,动物试验中,HCT挥发油能够显著升高去卵巢骨质疏松小鼠血清SOD活性,显著降低血清MDA含量,并呈剂量依赖性[13]。活性氧清除方面,HCT挥发油能降低脂多糖诱导的巨噬细胞中NO的产生和NO合成酶(nitric oxide synthase,iNOS)蛋白的合成[14]。癸酰乙醛(鱼腥草素)是其发挥抗氧化的主要物质,鱼腥草素不稳定,常与亚硫酸氢钠化合形成鱼腥草素钠(sodium houttuyfonate,SH),病理学检查发现,SH能够降低脂多糖诱导的肺组织损伤小鼠肺细胞MAD水平,提高SOD水平,抑制氧化应激和炎症反应[15]。
1.3 HCP抗氧化应激作用
HCP通过清除自由基、提高抗氧化酶活性,起到抗氧化作用。从鱼腥草叶中所分离出的甘露糖、半乳糖、葡萄糖,具备DPPH·清除能力,但清除能力不及维生素C,同时能够螯合Fe2+,还原铁氰化钾,表现出抗氧化作用[16]。体内试验表明,通过每周1次持续6周灌服不同剂量(50,100和200 mg/kg体重)HCP溶液,能够显著提高脂多糖诱导的慢性炎症大鼠肝组织中SOD、CAT、GSH-Px含量,且除了50 mg/kg体重组,其余剂量组MDA含量显著降低,提升肝组织抗氧化能力[4]。HCP抑制脂多糖诱导的小鼠支气管肺NO的产生[17]。
1.4 HCT提取液抗氧化作用
HCT无论是水提物还是醇提取物也表现出抗氧化作用。体外动物试验发现,2%和5% HCT水提物能够增强煎炸油喂养诱导SD大鼠的氧化应激能力,提高血浆、肝脏总的抗氧化能力,肝脏低密度脂蛋白滞后时间延长[18]。在糖尿病大鼠试验模型上同样得到验证,2% HCT水提物降低糖尿病小鼠心脏和肾脏中ROS活性、TNF-α、IL-6水平,恢复心脏GSH含量[19];HCT水提物(1和2 g/L)的预摄取可维持对乙酰氨基酚诱导小鼠肝脏GSH含量,激活GSH-Px、CAT、SOD,减少氧化型谷胱甘肽(Glutathione Oxidized,GSSG)、MDA和ROS的形成[20]。细胞同样证实HCT水提物抗氧化作用,与其他植物(鬼针草、枸杞、薄荷、车前、积雪草)水提物相比较,同一水提物浓度(25 μg/mL)下,HCT水提物处理H2O2诱导的淋巴细胞DNA彗星状拖尾运动现象百分比最低(11.14%),经ABTS法测定,HCT甲醇提取物显示出较高TEAC值(231.16 mmol/L),提示HCT提取物表现出较高的抗氧化能力,降低H2O2诱导的淋巴细胞DNA凋亡[21]。另外,HCT水提物(8 μmol/L)预处理,抑制β淀粉样蛋白暴露大鼠初级皮层神经元ROS及Ca2+水平,保护线粒体膜[22]。
2 HCT抗氧化应激机制研究进展
平衡状态下的氧化应激参与细胞信号转导,过量氧化应激产生自由基,攻击组织细胞,引起细胞和组织内的病理变化。机体通过氧化还原信号传导来调节过氧化水平,鱼腥草作为植物抗生素,通过氧化应激信号传导通路调控,整合氧化还原途径,维持机体氧化应激平衡[7]。
2.1 PI3K/Akt信号通路介导的氧化应激
动物试验表明,芒果苷可激活胎盘中PI3K/Akt/mTOR通路,改善子痫前期小鼠胎盘氧化应激[23],磷脂酰肌醇3-激(phosphatidylinositol3-kinase,PI3K)阻滞剂LY29400能抑制姜黄素的抗氧化作用,恢复甘露醇诱导的心肌细胞内ROS活性[24]。提示ROS通过PI3K/Akt信号通路发挥抗氧化作用。另有学者发现细胞内氧化应激水平提高后,可引起PI3K/Akt信号通路的代偿性激活,进而调节血红素加氧酶(heine oxygenase-1,HO-1)的表达和细胞质内核因子E2相关因子2(nuclear factor-E2-related factor 2,Nrf2)的转录激活,起到抗氧化应激的作用[25]。细胞试验发现,HCT水提物通过激活PI3K/Akt信号通路、抑制Toll样受体4(Toll-like receptors4,TLR4)、核因子κB(nuclear factor kappa-B,NF-κB),进而保护丙泊酚诱导的大鼠海马神经元损伤[26]。PI3K/Akt信号通路级联叉头框蛋白O(Forked(frame protein O,FoxO),FoxO激活过氧化物酶体增殖物激活受体γ共激活因子1(peroxisome proliferator-activated receptorγcoactiva-tor-1α,PGC-1)PGC-1α与FoxO共同转录表达,促进锰超氧化物歧化酶(manganese superoxide dismutase,MnSOD)、CAT的表达,增加抗氧化系统的功能[27]。体外试验发现,饱和脂肪酸处理引起人血管内皮细胞发生氧化应激反应,而HCT水提液预处理通过增强FoxO1表达,激活PGC-1α调节细胞线粒体功能,降低氧化应激反应[28]。可以发现,HCT提取物通过PI3K/Akt信号通路级联下游的FoxO1转录表达,激活PGC-1α,促进抗氧化酶的生成,提高细胞抗氧化作用。
2.2 Nrf2/ARE信号通路介导的氧化应激
Nrf2/ARE信号通路细胞抗过氧化的关键通路,正常情况下,Nrf2维持较低水平表达量。在氧化应激源苯并(a)芘的刺激下,Nrf2、Kelch样环氧氯丙烷相关蛋白1(Kelch-like epichlorohydrin-associated protein 1,keap1)表达激活,RNA干扰发现抗氧化基因的表达与(Cu/Zn-SOD、CAT和GPx)基因的表达一致[29]。研究发现,Nrf2/keap1通路级联核内抗氧化反应元件(anti-oxidant response element,ARE),诱导抗氧化酶包括GSH、HO-1等表达增加[30],提示Nrf2/keap1/ARE信号通路是诱导抗氧化基因所必需的。在动物试验模型中,HO-1的表达水平升高,NF-κB的活化,降低下游炎症因子的分泌,维持细胞正常氧化应激水平[31]。体外细胞试验发现,HCT水提物抑制TNF-α诱导的人角质形成细胞ROS的生成,并激活芳香烃受体(Aryl hydrocarbon receptor,AHR)和Nrf2,调节NADPH氧化酶(NADH oxidase,Nox),发挥抗氧化应激作用[32]。另有试验证实,HCT抑制由苯并(a)芘诱导肺肿瘤细胞Nrf2、HO-1、醌氧化还原酶1(NADPH quinone dehydrogenase 1,NQO-1)蛋白表达下降,降低由苯并(a)芘诱导肺肿瘤细胞内ROS活性[1]。可以看出,HCT及其提取物通过Nrf2的激活,促进NQO-1、HO-1的表达,促进体内自由基的清除,发挥抗氧化作用。
2.3 TLRs信号通路介导的氧化应激
在氧化应激源刺激下,人单核细胞中TLR2/4 mRNA/蛋白表达水平升高,抗氧化剂罗布宁/N-乙酰半胱氨酸处理后,TLR2/4 mRNA/蛋白表达水平升下降,H2O2诱导促进应激源刺激下TLR2/4基因的表达[33],TLRs偶联下游炎症因子(mitogen-activated protein kinase,MAPK)、NF-κB,导致氧化应激损伤[34],下调Bcl-2(B cell lymphoma 2,Bcl-2),上调凋亡相关因子Bcl-2相关X蛋白(Bcl-2 Associated X Protein,BAX)、冬氨酸蛋白酶3(caspase-3)、caspase-9,诱导细胞凋亡[35]。细胞试验证实,HCT提取物通过抑制TLR4/NF-κB通路,保护由丙泊酚诱导大鼠海马神经元损伤[26]。更为重要的是,HCT黄酮苷金丝桃苷、槲皮苷能有效抑制被H1N1病毒感染大鼠细胞内TLRs信号转导,抑制病毒复制[36]。动物试验发现,HCT挥发油能通过抑制DOX诱导的慢性大鼠TLR4表达,降低氧化还原酶介导的NF-κB炎症,抑制心肌细胞晚期凋亡[3]。在吸烟联合气管内滴注脂多糖诱导慢性阻塞性肺疾病大鼠模型上,低、中、高浓度HCT提取液下调caspase-3、caspase-9、TLR4、接头蛋白髓样分化因(myeloiddifferentiationfactor88,MyD88)和NF-κB(p65)的蛋白表达,提示HCT提取液通过抑制TLR4/MyD88/NF-κB(p65)信号通路的激活,改善慢性阻塞肺疾病大鼠肺组织淤血、水肿、炎症和凋亡[37]。更有研究证实,HCP通过TLR4、p-NF-κB(p65)的表达,抑制H1N1感染小鼠肺、肠炎症[38]。另外动物试验发现,HCT水提物通过抑制MAPK(p38)改善高脂血症诱导的内皮损伤[31],细胞试验同样证实HCT水提物通过抑制MAPK(p38)降低脂多糖诱导的视网膜胶质细胞的激活[39]。可以看出,HCT及其提取物能够通过抑制TLRs/MyD88/NF-κB(p65)/MAPK(p38)通路的激活,可能通过下调caspase-3、caspase-9,保护氧化应激引发细胞凋亡。这些数据表明,HCT通过抑制p-p38-MAPK来抑制LPS诱导的视网膜小胶质细胞的激活。
2.4 线粒体介导的氧化应激
内源性ROS主要由线粒体呼吸链活动产生,线粒体功能受损,呼吸链电子传递的过程中出现电子漏出,与单分子氧进行单电子还原生成超氧阴离子,引起细胞内氧化应激反应[40]。过量的氧化应激线粒体通透性转换孔开放(mitochondrial permeability transition pore,MTPT),线粒体Ca2+外排,线粒体膜电位(mitochondrial membrane potentia,MMP)改变,胞内Ca2+浓度升高[41]。MTPT持续开导致线粒体膜去极化,细胞凋亡起始因子细胞色素C(Cytochrome C,Cyt-C)外泄,在胞质内激活caspase-9、caspase-3,与过氧化应激反应所激活的Bcl-2/Bax一起共同促进细胞凋亡[42]。另外,去极化的线粒体膜电位后,PTEN诱导推定激酶1(PTEN induced putative kinase 1,PINK1)募集并活化帕金基因表达引起线粒体自噬[43]。动物试验和细胞试验都表明,HCT及其提取物能够通过多种途径,调节线粒体依赖的氧化应激,在D-半乳糖胺诱导的肝损伤小鼠模型上,HCT 甲醇提取物中分离的黄酮醇糖苷,可通过Rev-Erb-α/磷酸-AMPK/SIRT1信号传导调节线粒体自噬,增强线粒体生物合成,降低线粒体自噬相关蛋白 PINK1和帕金基因表达的增加[44]。同时,HCT水提物通过稳定线粒体膜电位水平,降低细胞内Ca2+浓度,抑制caspase-3激活,保护由β淀粉样蛋白诱导大鼠原代皮层神经元[28]。由此可以看出,HCT及其提取物通过降低胞内Ca2+,恢复MMP水平,稳定线粒体功能,减轻过度氧化应激。另外,HCT及其提取物通过增加Bax/Bcl-2比值,抑制caspase-9、caspase-3,降低线粒体自噬相关蛋白PINK1、帕金表达,减轻因过度氧化应激而造成的细胞凋亡。
3 结语
体内外试验表明:HCT及其提取物通过增强抗氧化酶活性,清除自由基,表现出较强的抗氧化应激作用,HCT作为一种天然抗氧化物,通过调控PI3K/Akt信号通路、Nrf2/ARE信号通路、TLRs信号通路及线粒体介导的信号通路,发挥抗氧化作用(图1)。HCT及其提取物通过PI3K/Akt信号通路,级联下游FoxO1激活PGC-1α,降低细胞氧化应激,而氧化应激模型中PI3K/Akt信号通路激活,可调节Nrf2、HO-1转录[25],可以得到启示HCT提取物通过PI3K/Akt/FoxO/PGC-1α通路提高细胞抗氧化能力的同时,也激活Nrf2/HO-1通路启动抗氧化应激。近期的细胞试验发现,HCT提取物激活PI3K/Akt信号通路、抑制TLR4/NF-κB,调控Nrf2/HO-1发挥抗氧化应激作用[25],提示HCT提取物可能通过PI3K/Akt信号通路的激活、TLR4/NF-κB的抑制,促进Nrf2/HO-1的抗氧化作用。而近期的试验证实HCT提取物去甲头花千金藤二酮B,通过激活PI3K/Akt、提高HO-1,帮助海马神经元细胞抵抗H2O2诱导的氧化应激[45]。除此之外,HCT提取物抑制TLR4/MyD88/NF-κB(p65)通路[38],抑制MAPK(p38)[28-39]、下调caspase-3、caspase-9,防止细胞过氧化凋亡[38],而HCT及其提取物在线粒体途径依赖氧化应激通路中,HCT能增加Bax/Bcl-2比率,抑制caspase-9、caspase-3激活[45],防止细胞凋亡,提示HCT提取物可能通过TLR4/MyD88/NF-κB,同时抑制线粒体、胞浆凋亡蛋白的激活,共同抵御氧化应激损伤。
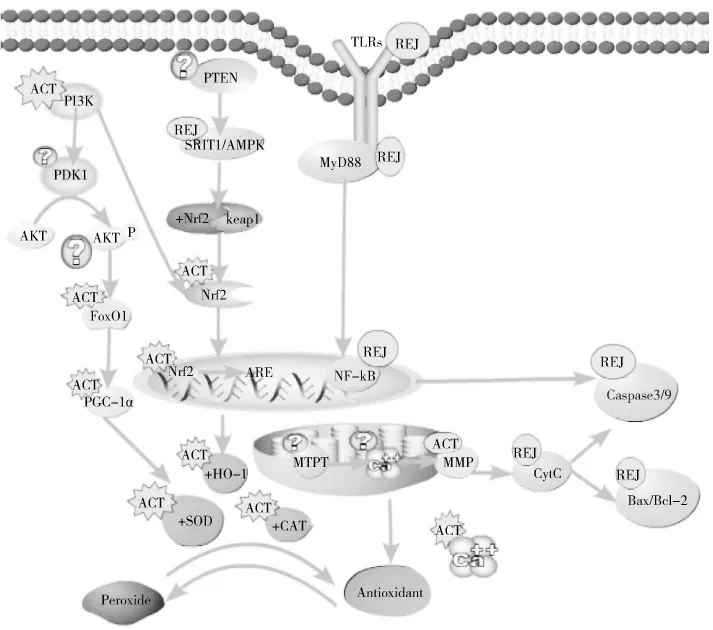
图1 HCT及其提取物抗氧化应激机制
HCT及其提取物抗氧化应激机制涉及多个信号通路、多个信号因子,各个信号通路、信号分子之间相互作用,氧化应激反应。未来可从多种途径、多种分子水平系统性地研究鱼腥草抗氧化应激机制,以及鱼腥草抗氧化应激不同信号通路,从而丰富鱼腥草抗氧化应激的理论研究内容和提供实践应用指导。
