2株野生木耳液体培养方法优化及其胞内多糖的抗氧化活性分析
2022-10-17冯小飞
冯小飞 朗 丹
1 寸孟人1 胡珊苑1 余 浪1 杨 斌2
(1. 西南林业大学生命科学学院,云南 昆明 650233;2. 西南林业大学生物多样性保护学院,云南 昆明 650233)
木耳属(Auricularia)真菌是我国常见的野生食用菌,常生长于林间阔叶树倒木及枯死木桩,由于其特殊的生物学特性,木耳属真菌能够分解多种材料中的纤维素、半纤维素、木质素等物质提供生长所需的营养[1-3]。木耳在我国栽培历史悠久,从初期的椴木“砍花”栽培到如今的袋料菌棒培育已有1 000多年历史[4],经过食用菌工作者的努力和研究,木耳人工栽培技术目前已经趋于成熟,多篇文献报道了菌种驯化、栽培种生产、替代基质筛选等方面的内容,为食用菌产业生产提供了较好的指导和借鉴[5-6]。黑木耳(Auriculoria auricular)和毛木耳(Auriculoria polytricho)是木耳属真菌中常见的2个种,也是我国目前主要的木耳栽培品种,据吴芳等[2]的报道,早在2011年,我国黑木耳年产鲜质量就达到了346.1万t,毛木耳年产鲜质量达到了143.5万t。随着木耳栽种规模的不断扩大,每年对菌种的需求也越来越大,菌种的质量将直接影响着木耳产品的质量和产量。液体发酵培养技术是目前食用菌产业中菌种生产的主要方法,合适的发酵培养条件是菌种生产、真菌代谢产物开发过程中的重要保障[7-8]。盛思远等[9]的研究发现,向液体发酵培养基质中添加维生素B1和磷酸二氢钾能够显著提高菌丝体中的抗氧化酶活性,从而缓解菌丝体的老化现象。于海洋等[10]利用正交实验优化了黑木耳的液体培养条件,通过条件优化提高了黑木耳菌丝体的生物量和多糖含量。柴新义等[11]通过液体培养条件的优化,提高了黑木耳菌丝体对硒元素的富集效果。所以,开展木耳菌株液体培养条件的筛选和优化是促进木耳栽培及发酵产品开发的重要渠道。
多糖是木耳属真菌中一类重要的化合物,广泛存在于木耳子实体、菌丝体等组织中,近代研究表明,包括木耳在内的多种食用菌多糖是一类具有开发价值和利用潜力的天然化合物,对调节机体免疫力、抗氧化、抗肿瘤等方面表现出了良好的活性[12-15]。例如,黑木耳中的水溶性多糖通过调节小鼠肠道微生物群落来降低由葡聚糖硫酸钠诱导的小鼠结肠炎[16-17]。角质木耳、毛木耳、黑木耳胞内多糖能够显著促进细胞增殖,对抗炎和抑菌表现出了较好的活性,同时,黑木耳子实体中的吡喃糖多糖还具有调节血脂保护肝脏的作用[18]。孙学明等[19]以心脑血管病人为研究对象,进行不同阶段的黑木耳多糖摄入治疗,通过病人的康复自愈能力分析结果发现黑木耳多糖能够促进心脑血管人体的康复。庄伟等[20]、许海林等[21]对黑木耳多糖进行了提取及结构解析并发现黑木耳多糖可以通过促进巨噬细胞的增殖和吞噬能力,该作用在人体免疫调节方面具有广阔的应用前景。
本研究以云南地区的野生黑木耳及毛木耳菌株为研究材料,通过液体摇瓶培养,研究不同碳源、氮源、无机盐、pH值等因素对2株野生木耳菌丝体生长情况的影响,筛选出适合2株野生木耳液体发酵培养条件的营养添加物。同时对2株野生木耳菌丝体胞内多糖抗氧化活性进行了研究,为该地区野生木耳的人工栽培及产品开发提供理论基础。
1 材料与方法
1.1 实验材料
1.1.1 材料
野生金江木耳菌株采自云南丽江市金江县(99°75′12″E,27°22′42″N)栓皮栎树桩上,毛木耳采自云南昭通市大关县三江口自然保护区(103°58′43″E,28°7′43″N)。“组 织分离法”对上述2种野生木耳子实体进行分离,菌丝萌发后经纯化培养获得纯菌落,菌落及子实体经形态鉴定后,菌种保存于西南林业大学生命科学学院生物化学教研室,子实体及菌落如图1所示。

图 1 野生金江木耳、毛木耳子实体及菌丝培养照Fig. 1 The picture of subentity and mycelia cultured of wild Jingjiang's black fungus and A. polytricha
1.2 试验方法
1.2.1 菌种活化和培养
将2种野生木耳菌种进行活化培养,无菌条件下接种于PDA培养基(直径9 cm)中央,于(25 ± 2) ℃条件下培养8 d后备用。
1.2.2 基础液体发酵培养基
葡萄糖2.0%,土豆20.0%,酵母浸膏0.3%,KH2PO40.1%,水1L,pH自然。
1.2.3 液体培养条件的优化
1)培养时间、温度、pH值、碳源、氮源、无机盐的筛选: 无菌条件下分别将2种野生木耳菌种打孔为大小相同的“菌饼”(直径0.5 cm),接种至不同pH值(pH值为5、6、7、8、9)、不同碳源(蔗糖、麦芽糖、果糖、淀粉、葡萄糖)、不同氮源(蛋白胨、牛肉膏、尿素、(NH4)2SO4、酵母膏)、不同无机盐(MgSO4、CaCl2、CuSO4、NaCl、KH2PO4)的基础培养基中,于25 ℃,160 r/min条件下摇床培养10 d;培养时间及温度实验将菌种接种至基础培养基中,分别于25 ℃,160 r/min条件下培养不同天数(2、4、6、8、10 d)、以及不同温度(20、25、30、35、40 ℃)条件下,160 r/min摇床培养10 d。每瓶发酵培养液接种6个菌饼(150 mL/瓶),每个单因素设置3个重复,按时取出不同培养条件的发酵培养物,过滤、洗涤,85 ℃烘4 h后称重,记录不同培养条件下2种木耳菌丝干质量。
2)正交实验 在6个单因素实验的基础上,采用了L4(23)正交表对碳源、氮源、无机盐、进行实验设计,以获得2种野生木耳最佳的液体发酵培养条件。
1.2.4 菌丝体胞内多糖的提取
参照安雪菲等[22]方法,略有改动。分别取5.0 g粉碎处理的2种野生木耳菌丝体于50 mL离心管,加入20.0 mL水于80 ℃,800 W条件下超声波提取10 min,离心收集上清液,重复提取3次后合并提取液。粗多糖提取液经过乙醇过夜沉淀,除蛋白后乙醇过夜二次沉淀,离心弃上清液,少量蒸馏水溶解,低温处理后冷冻干燥仪冻干备用。
1.2.5 菌丝体胞内多糖抗氧化活性的测定
总还原力测定采用铁氰化钾还原法,以700 nm处样品吸光值的大小评价还原力的强弱;羟基自由基(·OH-)清除能力采用水杨酸法测定;超氧阴离子()清除能力采用邻苯三酚自氧法测定;DPPH自由基(DPPH·)清除能力参考王倩朝等[23]、郭磊等[24]的方法测定。上述指标测定过程中以相同浓度抗坏血酸(Vc)为阳性对照,每组实验重复3次。

式中:A0为空白对照液的吸光值,Ax为加入样品溶液后的吸光值,Ax0为加入提取液后的吸光值。
1.3 数据处理
以SPSS Statistics 19.0统计软件对测定数据进行处理和分析,结果以平均值 ± 标准差表示;以Excel软件进行柱状图绘制。
2 结果与分析
2.1 单因素筛选结果
2.1.1 时间对发酵培养的影响
如图2所示,2种野生木耳菌丝体干质量与培养时间均正相关的趋势,野生金江木耳菌丝体生长较快,液体摇瓶培养6 d时菌丝体干质量达到了0.98 g,且在后续培养的3个时段中,菌丝体生物量变化不显著,于10 d时菌丝体干质量最大为1.0 g。野生毛木耳菌丝体生物量随着培养时间的增加而逐渐升高,10 d时,菌丝体干质量最大为0.89 g,显著(P< 0.05)高于其他时段的质量。

图 2 不同发酵培养时间对2株野生黑木耳菌丝体干质量的影响Fig. 2 Effects of different fermentation time on dry weight of 2 wild A. auricular mycelia
2.1.2 温度对发酵培养的影响
如图3所示,2株野生木耳在20~35 ℃的温度下都能够正常生长,且菌丝体生物量随着温度的增加而呈现出先增加后降低的趋势,温度为30 ℃时,野生金江木耳和毛木耳菌丝体生物量达到了最大,干质量分别为0.98 g和1.00 g。35 ℃后二者菌丝体干质量急剧下降,显著低于20~30 ℃培养条件下的菌丝体质量(P<0.05),40 ℃时供试木耳菌株不能正常萌发和生长。30 ℃是野生金江木耳和毛木耳液体摇瓶培养较为合适的培养温度,当温度超过35 ℃后,高温影响了2株野生木耳菌丝体的正常生长和发育,造成菌丝体生物量降低,甚至菌种不能萌发和生长。
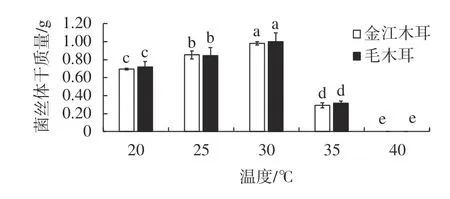
图 3 不同发酵培养温度对2株野生黑木耳菌丝体干质量的影响Fig. 3 Effects of different fermentation temperature on dry weight of 2 wild A. auricular mycelia
2.1.3 pH对发酵培养的影响
如图4所示,2株野生木耳在pH值5~9的范围内都能够正常生产,且菌丝体生物量随着发酵液pH值的变化呈现出先增加后降低的趋势。当pH为7时,野生金江木耳和毛木耳菌丝体干质量最大,分别为0.86 g和1.04 g,显著大于其他pH培养条件下的菌丝体干质量(P<0.05)。pH值大于8以后供试木耳菌株菌丝体生物量显著降低(P<0.05),当pH为9时,2种木耳菌丝体干质量最小,仅为0.06 g和0.08 g。

图 4 不同发酵培养pH值对2株野生黑木耳菌丝体干质量影响Fig. 4 Effects of different pH values on dry weight of 2 wild A. auricular mycelia
2.1.4 碳源对发酵培养的影响:
如图5 所示,2株野生木耳菌株都能够在5种不同的发酵培养基中正常生长,其中,葡萄糖为主要添加物时,二者菌丝体生物量均较大,分别为0.92 g(毛木耳)和0.89 g(金江木耳),其次为麦芽糖,且麦芽糖、果糖、蔗糖对毛木耳菌丝体干质量影响差异不显著,以淀粉为碳源发酵培养时,供试的22株木耳菌株菌丝体生长状况均较弱,菌丝体干质量仅为0.72 g(毛木耳)和0.61 g(金江木耳),显著低于葡萄糖和麦芽糖的培养结果(P<0.05)。葡萄糖作为生命活动过程中最重要的单糖,具有溶解性好,能够直接参与生物氧化反应提供能量,因此在本次单因素实验中,葡萄糖为碳源进行发酵培养后2株木耳菌丝体生物量均较大。

图 5 不同碳源对2株野生黑木耳菌丝体干质量的影响Fig. 5 Effects of different carbon sources on dry weight of 2 wild A. auricular mycelia
2.1.5 氮源对发酵培养的影响
如图6所示,2株野生木耳在生长过程中既能够利用有机氮元素,同时也能够利用无机氮源,添加蛋白胨时,2株野生木耳发酵后菌丝体干质量最大,分别为1.24 g(毛木耳)和0.87 g(金江木耳),牛肉膏的培养效果次之,供试菌株菌丝体干质量分别为 1.02 g(毛木耳)和0.84 g(金江木耳),虽然蛋白胨对供试野生木耳菌丝体的培养效果略优于牛肉膏,但是二者对供试木耳菌丝体干质量影响差异不显著。添加尿素为氮源时,2株野生木耳菌丝体生长情况较弱,菌丝体干质量分别为0.78 g(毛木耳)和0.68 g(金江木耳),与(NH4)2SO4相比,菌丝体干质量差异不显著。蛋白胨、牛肉膏含有种类更为丰富的氨基酸、多肽、蛋白质等营养物质,所以添加蛋白胨进行液体发酵培养后能够促进菌丝体生物量的积累,培养效果明显比无机氮源好。

图 6 不同氮源对22株野生黑木耳菌丝体干质量影响Fig. 6 Effects of different nitrogen sources on dry weight of 2 wild A. auricular mycelia
2.1.6 无机盐对发酵培养的影响
如图7所示,2株野生木耳在供试的5种无机盐中都能够正常生长,添加KH2PO4后,2种野生木耳菌丝体生长状况均较好,菌丝体干质量分别达到了0.75 g(金江木耳)和0.65 g(毛木耳),显著高于供试的其他无机盐(P<0.05)。同时,添加CuSO4后,金江木耳菌丝体干质量最小,仅为0.14 g,添加NaCl后,毛木耳菌丝体干质量最小仅为0.22 g。比较图中数据可知,在发酵培养基中添加H2PO4能够促进2株野生木耳菌株菌丝体的生长,提高菌丝生物量,同时,CuSO4和NaCl不利于2株木耳菌丝体的生长。

图 7 不同无机盐对2株野生黑木耳菌丝体干质量影响Fig. 7 Effects of different inorganic salts on dry weight of 2 wild A. auricular mycelia
2.1.7 正交实验优化结果
以碳源、氮源、无机盐为主要考查因素,开展三因素二水平正交实验,结果如表1和2所示。从表中可以看出,野生金江木耳菌丝体干质量最大为1.04 g/150 mL,野生毛木耳菌丝体干质量最大为1.26 g/150 mL。从极值(R)比较发现3个因素对供试木耳菌丝体干质量的影响顺序为:氮源 > 碳源 > 无机盐。所以,2株野生木耳优化后液体发酵培养配方均为葡萄糖(2.0%)、蛋白胨(0.3%)、KH2PO4(0.1%)、土豆20.0%、初始pH值为7.0。

表 1 野生金江木耳液体发酵培养条件优化结果Table 1 The optimization result of liquid fermentation culture conditions of wild Auricularia jingjiang fungus
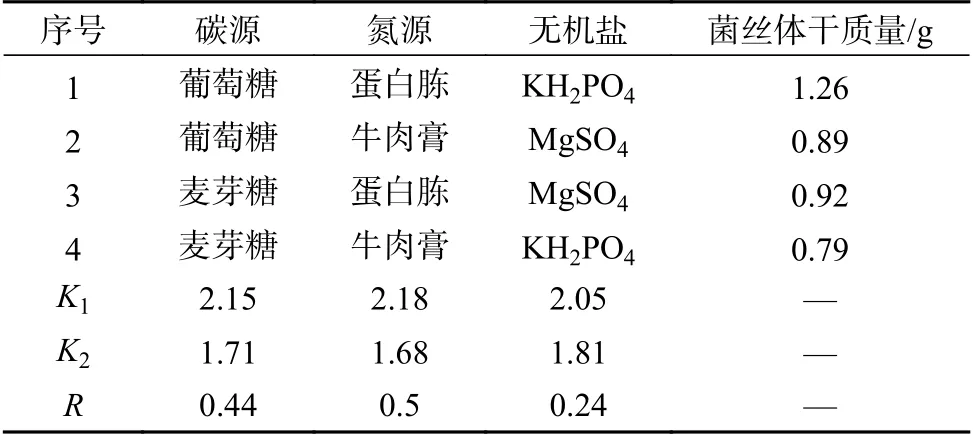
表 2 野生毛木耳液体发酵培养条件优化结果Table 2 The optimization result of liquid fermentation culture conditions of wild A. polytricha
2.2 2株野生木耳菌丝体胞内多糖抗氧化活性测定
2株野生木耳菌丝体胞内多糖抗氧化活性测定结果如表3所示,供试木耳菌丝体胞内多糖对·OH-、、DPPH·都表现出了一定的清除能力,且其抗氧化能力随着浓度的增加而呈现逐渐增强的趋势。相同浓度条件下野生金江木耳菌丝体胞内多糖还原力的测定结果显著高于野生毛木耳,但是二者均显著低于阳性对照Vc的测定值(P<0.05),浓度为0.50 mg/mL时,还原力测定值分别达到了0.47(金江木耳)和0.27(毛木耳)、1.62(Vc)。·OH-清除率结果显示,低浓度(0.10 mg/mL)时,2种野生木耳胞内多糖对·OH-的清除率差异不显著,随着浓度的增加,二者对·OH-的清除率逐渐增大,其中野生金江木耳菌丝体多糖的清除率显著高于相同浓度下野生毛木耳的测定值(P<0.05),浓度为0.50 mg/mL时,对·OH-的清除率分别为27.89%(金江木耳)和15.71%(毛木耳)、94.17%(Vc)。同时,2种野生木耳菌丝体胞内多糖对的清除效果差异不显著,相同浓度下二者的清除率均显著小于Vc的测定值(P<0.05),浓 度 为0.50 mg/mL时,对的清除率最强,分别达到了70.60%(金江木耳)、71.58%(毛木耳)、87.21%(Vc)。由DPPH·清除率测定结果可知,低浓度时(0.10 mg/mL)22种野生木耳菌丝体胞内多糖的清除率均较小,且差异不显著,随着多糖浓度的升高,二者对DPPH·的清除效果逐渐升高,其中金江木耳样品的清除效果显著强于毛木耳(P<0.05),清除率最高时分别达到了70.50%(金江木耳)和65.19%(毛木耳)。本次测定的4个抗氧化指标中,除了对清除率差异不显著外,野生金江木耳菌丝体胞内多糖对其他3种抗氧化活性指标的测定值均显著高于毛木耳,表现出了较好的抗氧化能力。

表 3 2株野生木耳菌丝体胞内多糖抗氧化能力测定结果Table 3 The antioxidant activity of intracellular polysaccharide in mycelium of 2 wild Auricularia strains
2.2.1 3种自由基半清除率(IC50值)的计算
由图8可知,2株野生木耳菌丝体胞内多糖对DPPH·清除率效果最好,IC50值分别为0.40、0.39 mg/mL,的IC50值分别为0.43 mg/mL(野生金江木耳),0.42 mg/mL(野生毛木耳),二者对和DPPH·的IC50值差异不显著。野生金江木耳菌丝体胞内多糖对·OH-的清除能力显著强于毛木耳(P<0.05),二者IC50值分别为0.67 mg/mL(金江木耳)和0.75 mg/mL(毛木耳)。综合比较图中数据可知,2株野生木耳菌丝体胞内多糖对DPPH·和的清除能力强于·OH-的清除能力。

图 8 2株野生木耳菌丝体胞内多糖对三种自由基的IC50值Fig. 8 The IC50 values of intracellular polysaccharides of 2 wild Auricularia mycelia for 3 kinds of free radicals
3 结论与讨论
菌丝体是真菌重要的营养组织,其生长状况影响着真菌后期的生长发育和子实体的分化,培养条件,基质营养成分往往对菌丝体的形态和代谢产物产生重要的影响,所以培养条件优化是菌种培育,发酵生产等多个领域中的重要环节[25-26]。本次实验结果显示,培养温度30 ℃、pH值为7.0、葡萄糖为碳源、蛋白胨为氮源、添加KH2PO4条件下供试木耳菌株培养效果较好,其中野生金江木耳菌丝体生长速度较快,上述条件培养6 d时菌丝体干质量达到了1.04 g/150 mL(菌丝体生物量增长率为0.17 g/d),野生毛木耳生长相对较慢,发酵培养10 d后菌丝体干质量达到了1.26 g/150 mL(菌丝体生物量增长率为0.13 g/d)。碳源、氮源、无机盐是微生物生长过程中不可或缺的营养物质,作为自然界中重要的单糖之一,葡萄糖直接参与糖酵解及三羧酸循环提供能量,与其他多糖或者二糖相比,葡萄糖无需水解,具有较高的利用率。蛋白胨为肉类、蛋白质类物质的酶水解物,含有丰富的小分子氨基酸和多肽,是一类重要的有机氮源,二者是微生物发酵培养过程中常用的营养添加物。例如:刘敏等[26]、李超等[27]、郭月仙等[28]分别对白背毛木耳(Auricularia polytricha)、黑木耳(Auricularia auricula)、玉木耳(Auricularia cornea)开展了培养条件研究,以葡萄糖为碳源进行发酵培养后,3种木耳菌丝体均获得了较好的生物转化率。车星星等[29]对黑木耳液体培养条件进行优化,以葡萄糖为碳源、蛋白胨为氮源、添加KH2PO4后黑木耳菌丝体生物量达到了1.63 g/100 mL,证实了以葡萄糖和蛋白胨为主要营养的发酵培养基配方能够提高木耳菌丝体的生物转化率,但是由于不同品种之间生物学特性的差异,本次研究中2株野生木耳菌丝体生物转化率略低于相关文献报道。抗氧化测定指标显示,2种野生木耳菌丝体胞内多糖对铁离子还原力、·OH-、、DPPH·都表现出了一定的抗氧化活性,且二者抗氧化效果与浓度呈现正相关趋势。金江木耳对3种自由基的IC50值分别为0.67 mg/mL(·OH-)、0.40 mg/mL(DPPH·)、0.43 mg/mL();毛木耳对3种自由基的IC50值分别为0.75 mg/mL(·OH-)、0.39 mg/mL(DPPH·)、0.42 mg/mL(·),且 二者 对DPPH·和的 清除能力差异不显著。以IC50值作为评价指标,金江木耳和毛木耳菌丝体孢内多糖对DPPH·的清除能力最强,二者IC50值均较小。木耳多糖是一类由醛糖或者酮糖通过糖苷键链接起来的多聚物,其抗氧化能力主要通过自由基清除及抗氧化酶活性的诱导实现[30]。罗敬文等[31]对玉木耳、黑木耳和毛木耳子实体多糖的抗氧化活性进行了研究,3种 木 耳 对·OH-的IC50值 在2.36~3.33 mg/mL之 间,的IC50值 在1.11~3.20 mg/mL之 间,DPPH·清除率的IC50值在1.32~10.34 mg/mL之间。孔沛筠等[23]研究结果也证实了黑木耳多糖具有抗氧化活性,从黑木耳子实体获得的多糖对、DPPH·的IC50值分别为0.65、1.47 mg/mL,金江木耳及毛木耳菌丝体胞内多糖对三种自由基的IC50值均较小,表现出了较强的自由基清除能力。本次研究开展了2株野生木耳液体发酵培养条件的优化及菌丝体胞内多糖抗氧化活性的测定,研究结果对云南地区野生木耳资源的利用和开发提供理论依据及参考价值。
