江西官山亚热带次生常绿阔叶林物种组成与群落结构特征
2022-10-15易伶俐彭巧华欧阳园兰曾小霞杨清培宋庆妮
王 倩,易伶俐,陈 琳,彭巧华,欧阳园兰,龚 超,曾小霞,杨清培,宋庆妮,刘 骏*
江西官山亚热带次生常绿阔叶林物种组成与群落结构特征
王倩1,易伶俐1,陈琳1,彭巧华1,欧阳园兰1,龚超2,曾小霞2,杨清培2,宋庆妮2,刘骏2*
(1. 江西官山国家级自然保护区管理局,江西 宜春 336000;2. 江西农业大学 江西省竹子种质资源与利用重点实验室,江西 南昌 330045)
【目的】常绿阔叶林是我国亚热带地区的地带性植被,但由于长期的人为干扰,导致地许多原始林变为次生林,为了更好地研究亚热带地区次生常绿阔叶林的恢复过程和多样性维持机制。【方法】按照美国史密森研究院热带森林科学研究中心(Center for Tropical Forest Science, CTFS)大型森林动态样地建设标准。在官山自然保护区建立12 hm2的次生常绿阔叶林动态监测样地,调查样地内DBH≥1 cm的木本植物,分析其物种组成、区系特征、径级结构等群落特征。【结果】1)样地内木本植物种类丰富且稀有种较多,共有65科139属312种,其中稀有种148种,约占总物种数的1/2;2)植物区系地理成分多样,有热带向温带过渡的性质,属水平上有28个类型(包含变型),热带区系成分占总属数的59.71%,温带区系成分占37.41%;3)群落径级分布呈“金字塔”型,属于增长型群落,优势种中大部分物种径级结构也呈“金字塔”型,处于增长状态。【结论】官山次生常绿阔叶林物种组成丰富,稀有种比例高,群落更新良好。
物种丰富度;种-多度格局;种-面积曲线;区系特征;径级结构
【研究意义】常绿阔叶林是亚热带陆地生态系统的重要组成,为稳定全球气候变化、维持全球碳循环平衡等方面起到关键作用[1]。但由于人类活动频繁导致中国常绿阔叶林主要为干扰后形成的次生林[2]。选择有代表性的地段进行群落特征的研究,对了解次生常绿阔叶林的恢复过程和物种多样性的维持机制具有重要的意义[3]。群落特征是森林生态系统功能的基础,为揭示群落演替、物种多样性维持机制等提供重要信息[4],主要特征包括物种组成、区系成分和年龄结构等,其中物种组成是形成群落的基础,在此基础上研究植物区系组成不仅有利于我们追溯区域内植物的变迁与进化体系,还可以为生物多样性的保护提供参考[5-6],种群年龄结构反映了植物的更新和生长现状,是判断种群动态和群落演替的重要依据。【前人研究进展】众多研究[7-8]表明传统的生态学研究方法具有尺度依赖性,近年来国内外兴起以样地为研究平台来开展植物群落组成与结构特征等方面的研究[9-13],如南亚热带地区的鼎湖山样地、中亚热带地区的古田山和天童山样地等,这些研究增强了人们对常绿阔叶林群落基础生态学机理的认识,提升了我国生物多样性研究水平。【本研究切入点】然而,这些研究基本是以保存较好的原始常绿阔叶林为研究对象,中国的常绿阔叶林主要为干扰后形成的次生林[2],其在物种组成、结构和外貌特征等方面与原始林存在明显的差异[14]。关于次生常绿阔叶林群落特征的关注较少,仅集中在小尺度[15-17],而小面积的样地调查研究,存在一定的局限性,不足以反映森林的树种组成与结构特征等,这不利于人们对次生常绿阔叶林恢复过程的了解及物种多样性维持机制的认识。因此,有必要对次生常绿阔叶林开展大样地建设工作,完善人们对其群落特征的认识。江西官山位于中亚热带地区,在成为自然保护区之前,有炼山造林的传统,大部分森林都曾遭受过烧毁,是开展次生常绿阔叶林相关研究的理想地点。【拟解决的关键问题】基于此,本研究参照CTFS大样地的建设方法,于2014年在江西官山国家级自然保护区中建立了12 hm2次生常绿阔叶林动态监测样地,调查并鉴定样地内DBH≥1 cm的所有木本植物,全面分析其物种组成、区系成分和径级结构等群落特征。以期探明以下问题:(1)官山次生常绿阔叶林由哪些物种组成?区系成分如何?(2)哪些物种占据优势地位?它们更新趋势如何?本研究补充了亚热带次生常绿阔叶林的数据库,使人们对亚热带地区森林群落的特征认识更加完善。
1 材料和方法
1.1 研究区域概况
江西官山自然保护区位于赣西北九岭山脉西段,该地年均气温16.2 ℃,年均降水量1 950~2 100 mm,四季分明、光照充足,属中亚热带暖湿东南季风气候区,地带性植被为亚热带常绿阔叶林[18]。森林覆盖率高达93.8%,高等植物2 344种,被子植物1 896种,国家重点保护野生植物21种和国家级珍稀濒危保护植物28种,分别占江西省的45.8%、46.4%、38.1%和51.8%[19]。
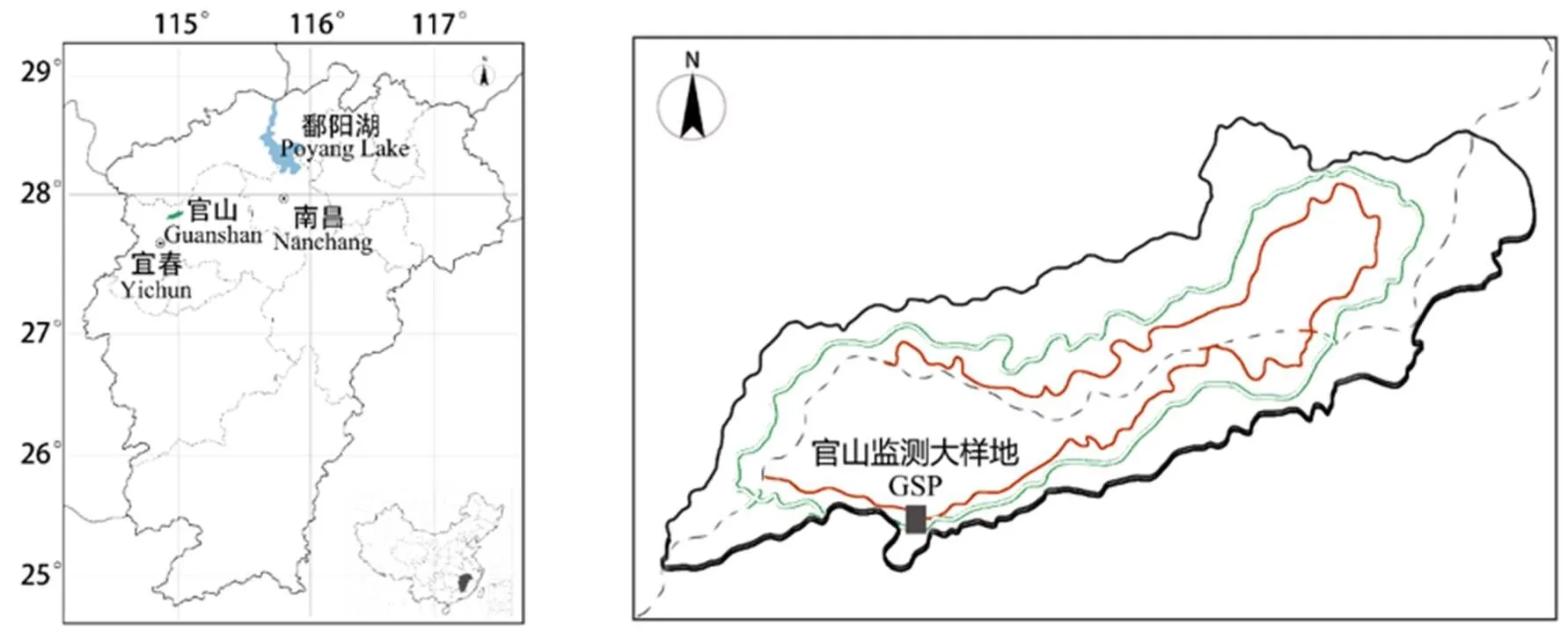
图1 官山常绿阔叶林动态监测样地地理位置
1.2 样地设置与数据采集
参照国际森林大型监测样地建设方法,采用全站仪等工具在宜春官山自然保护区建立了12 hm2大型森林监测样地(GSP),图1所示,样地西南角为原点(28°33′25″N,114°34′40″E),整个样地地形复杂,样地东北高,西南低,最高海拔645.0 m,最低海拔444.1 m,相对高差200.9 m,局部地段坡度在60 °以上。将整个样地划分300个20 m×20 m的样方,为方便物种坐标的读取,每个样方又分成16个5 m×5 m的工作样方[20]。采用相邻格子法进行每木调查,调查对象为胸径(DBH)≥1 cm的木本植物,在高度1.3 m处涂上红漆,并给每株木本植物编号挂牌。调查内容包括植物的编号、树种名称、胸径、样区位置(大样方号、小样方号、坐标)、光照情况、生长状况等,对于DBH≥5 cm的个体用胸径尺测量其胸径,1 cm≤ DBH<5 cm的个体则用数显游标卡尺测量。如果植株1.3 m以下有分枝,且分支大于1 cm则按照同样的方法挂牌测量(图2)。
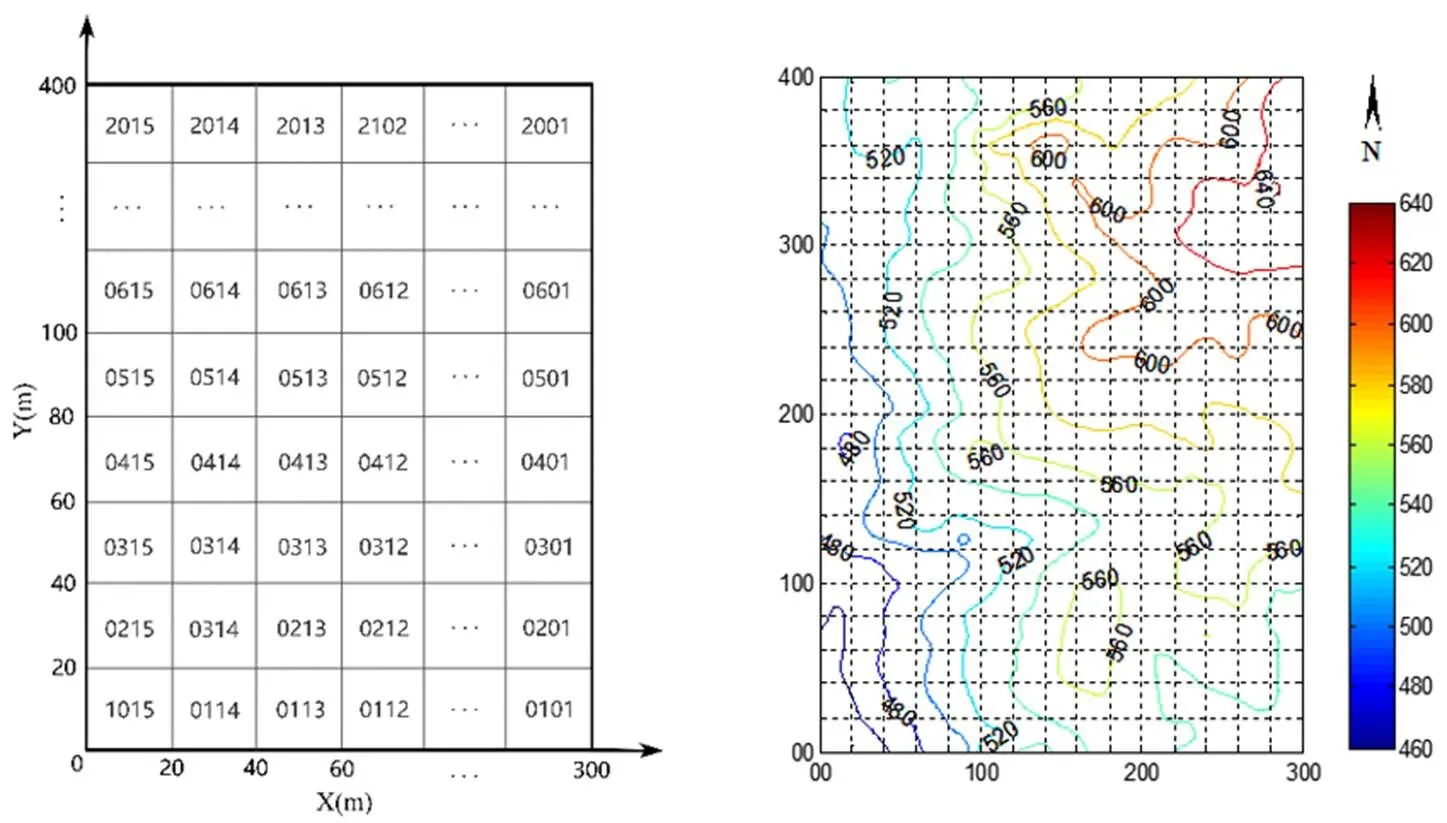
图2 官山常绿阔叶林动态监测样地分布与等高线图
1.3 数据处理
1.3.1种-多度格局及稀有种的统计物种-多度分布格局是群落结构的重要格局,一般指一定区域的群落内不同物种分布的数量。物种-多度曲线可以直观地表示所研究群落中稀有种和常见种所占比例等群落结构特征。根据物种个体数从高到低排序,分别统计个体数≥1 000株的植物种类、500~1 000株的植物种类、100~500株的植物种类和<100株的植物种类,从而绘制物种-多度曲线。按照Hubbell和Foster(1986)的定义[9],以个体多度≤1株/hm2的物种为稀有种,2~10 株/hm2的物种定义为偶见种。
1.3.2种-面积曲线种-面积曲线的绘制,以整个样地为计算单位,在样地内任意选取30个中心点,然后按1 m×1 m、2 m×2 m、3 m×3 m、4 m×4 m、5 m×5 m、10 m×10 m、20 m×20 m、30 m×30 m、40 m×40 m、50 m×50 m、60 m×60 m、70 m×70 m、80 m×80 m、90 m×90 m、100 m×100 m、200 m×200 m、300 m×300 m、300 m×400 m,18个面积梯度依次进行树种统计,并求出每种取样面积在30个中心点下的平均树种数,从而绘制种-面积曲线。本文选择了4条曲线作为种-面积曲线的拟合模型,其表达式如下[21]。
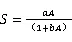
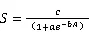
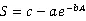
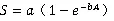
式中为面积,为物种数,、、为待定参数。
1.3.3科重要值与种重要值的计算重要值是一个综合性的数量指标,反映种群的生存状况及其对资源的利用能力[22],科的重要值综合了群落的丰富度、密度、优势度;物种的重要值综合了种群频度、密度、优势度及其在群落中的相对位置。它们的计算公式如下:
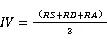
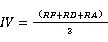
式(5)为科重要值的计算公式,式(6)[23]为种重要值的计算公式。式中为相对丰富度,是指一个科的物种数占总物种数的百分比;为相对多度,某个物种的株数与所有种的总株数的比;表示相对频度,为某个种(科)出现的次数与所有种出现的总次数的比;表示相对胸高断面积,为某个种(科)的胸高断面积与所有种的胸高断面积之和的比。计算时仅统计了独立个体的数量,不包括分枝和萌枝的断面积。
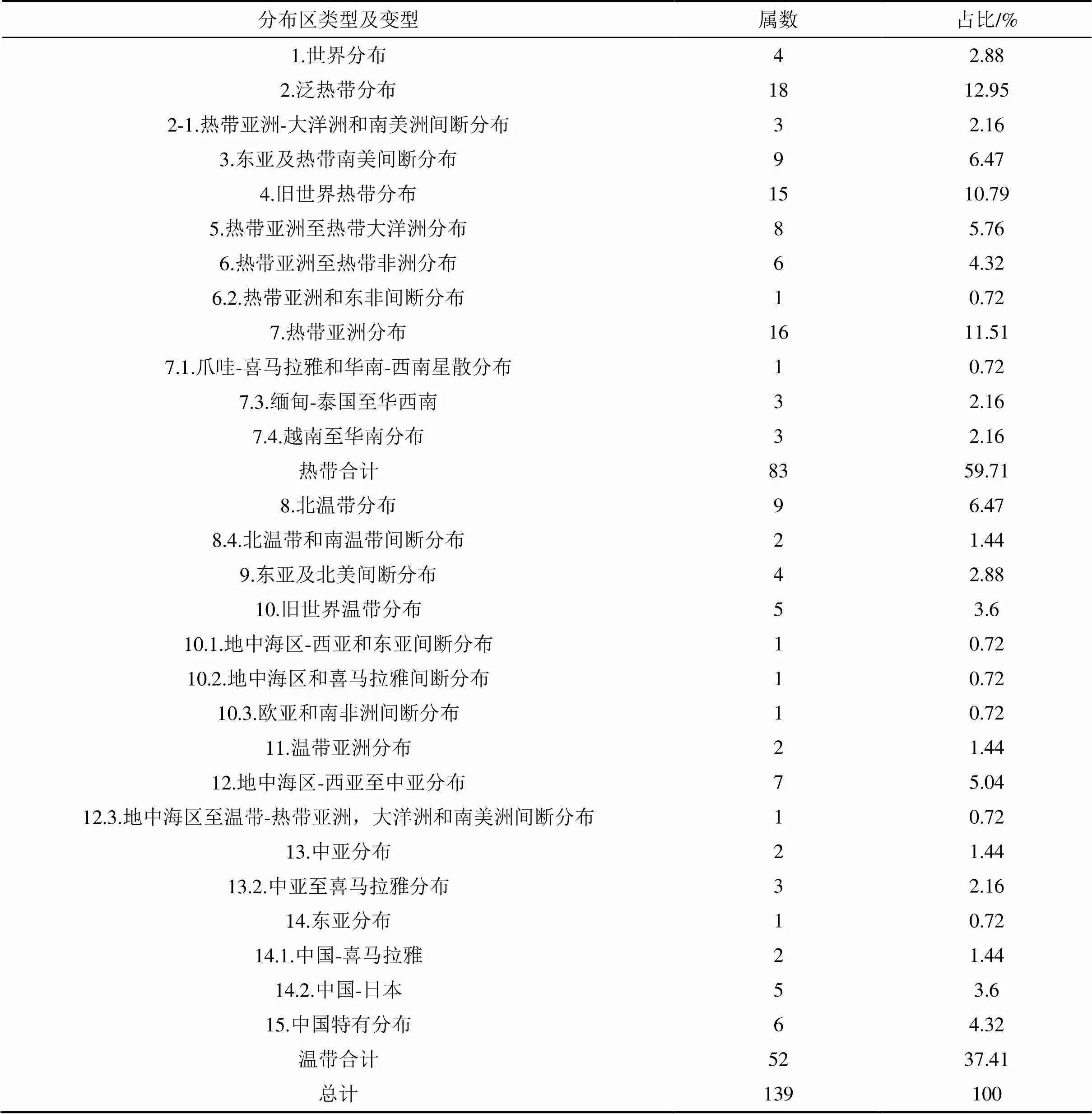
1.3.4植物区系按照《世界种子植物科的分布区类型系统》、《中国种子植物属的分布区类型》[24-25]对官山大型森林监测样地进行科、属区系分析。
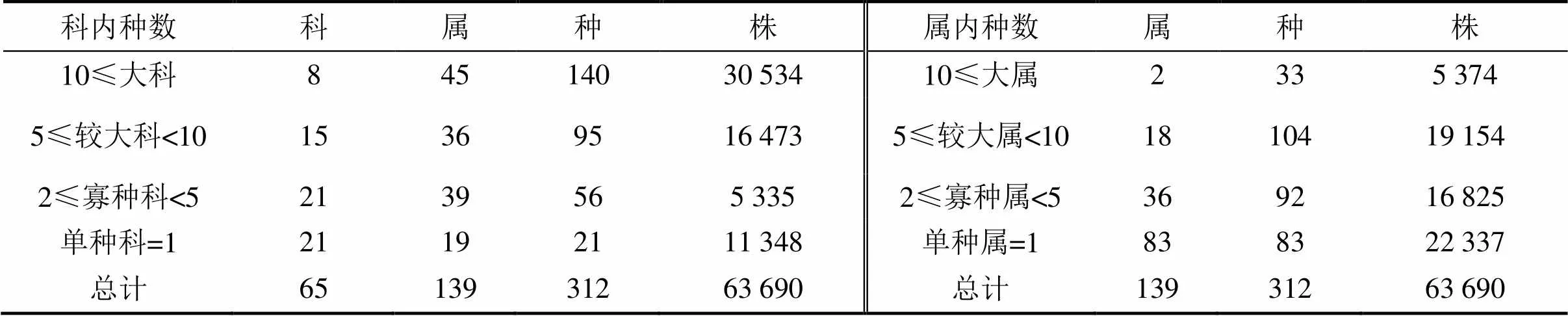
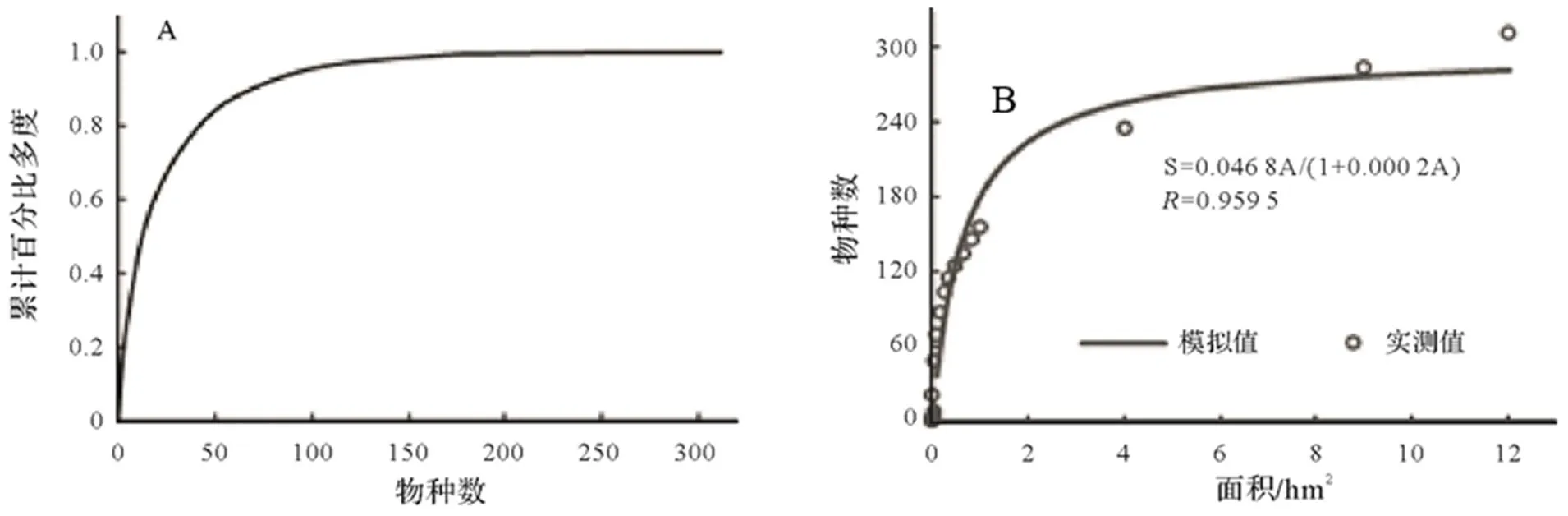
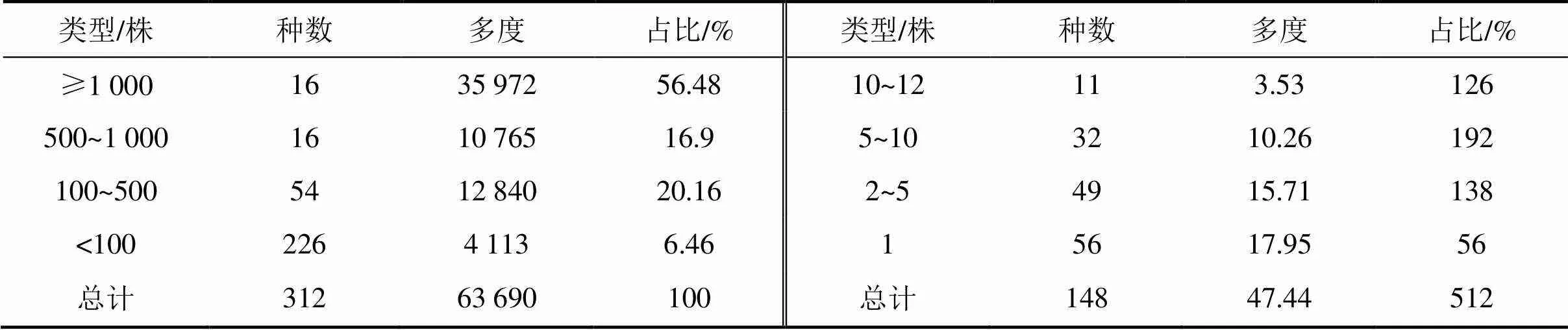
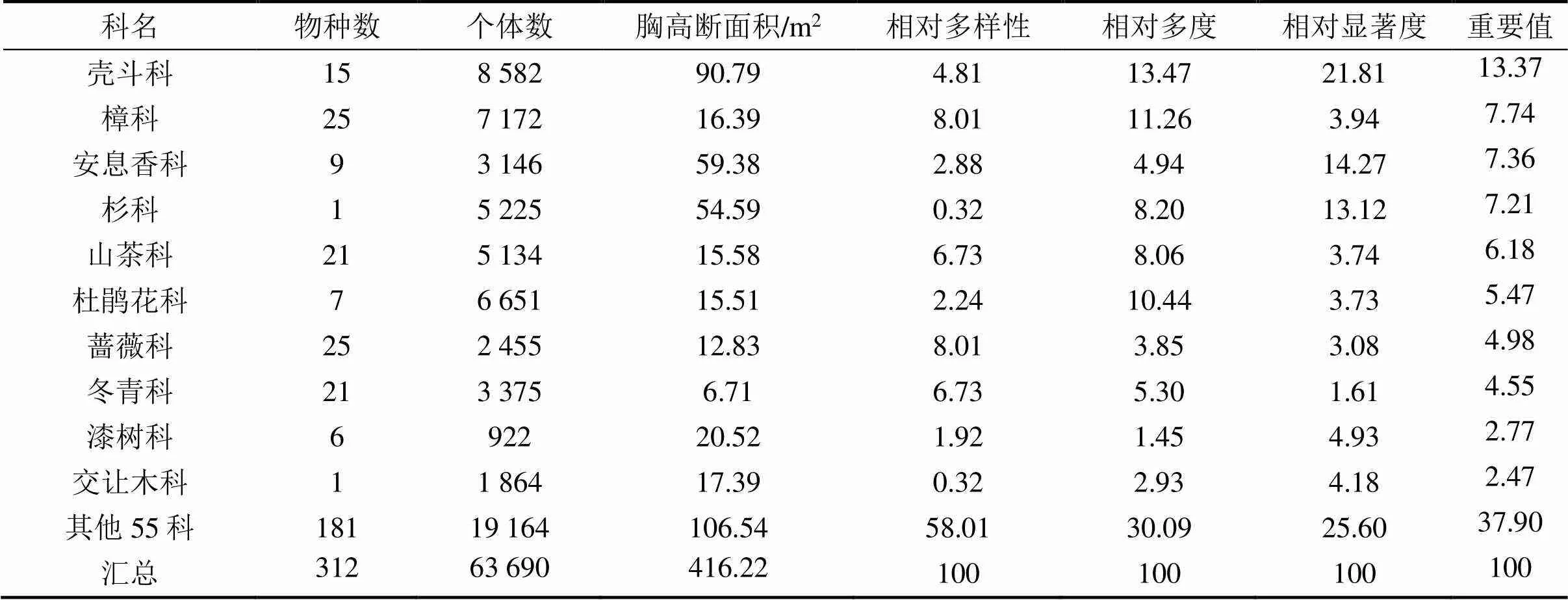
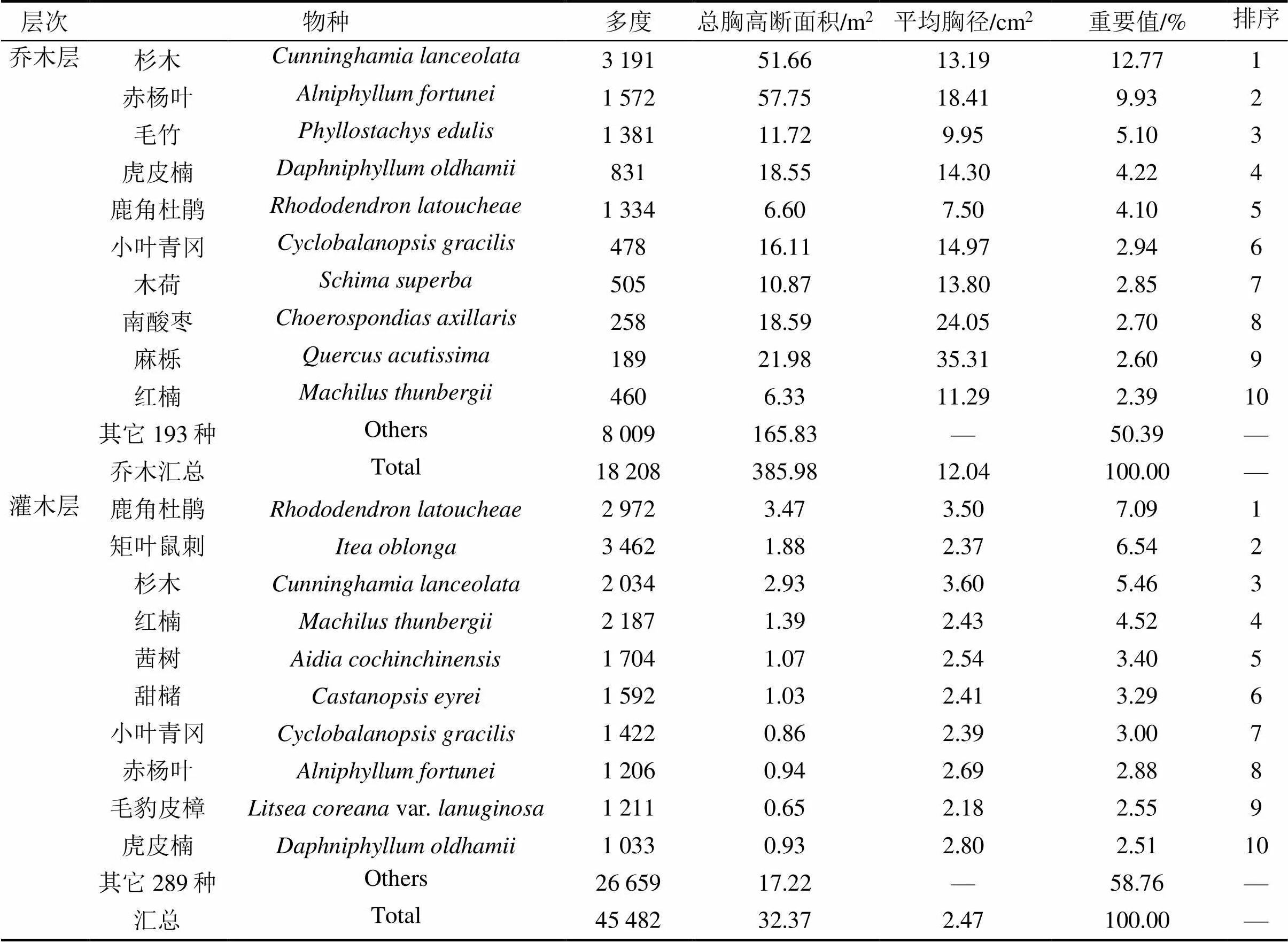
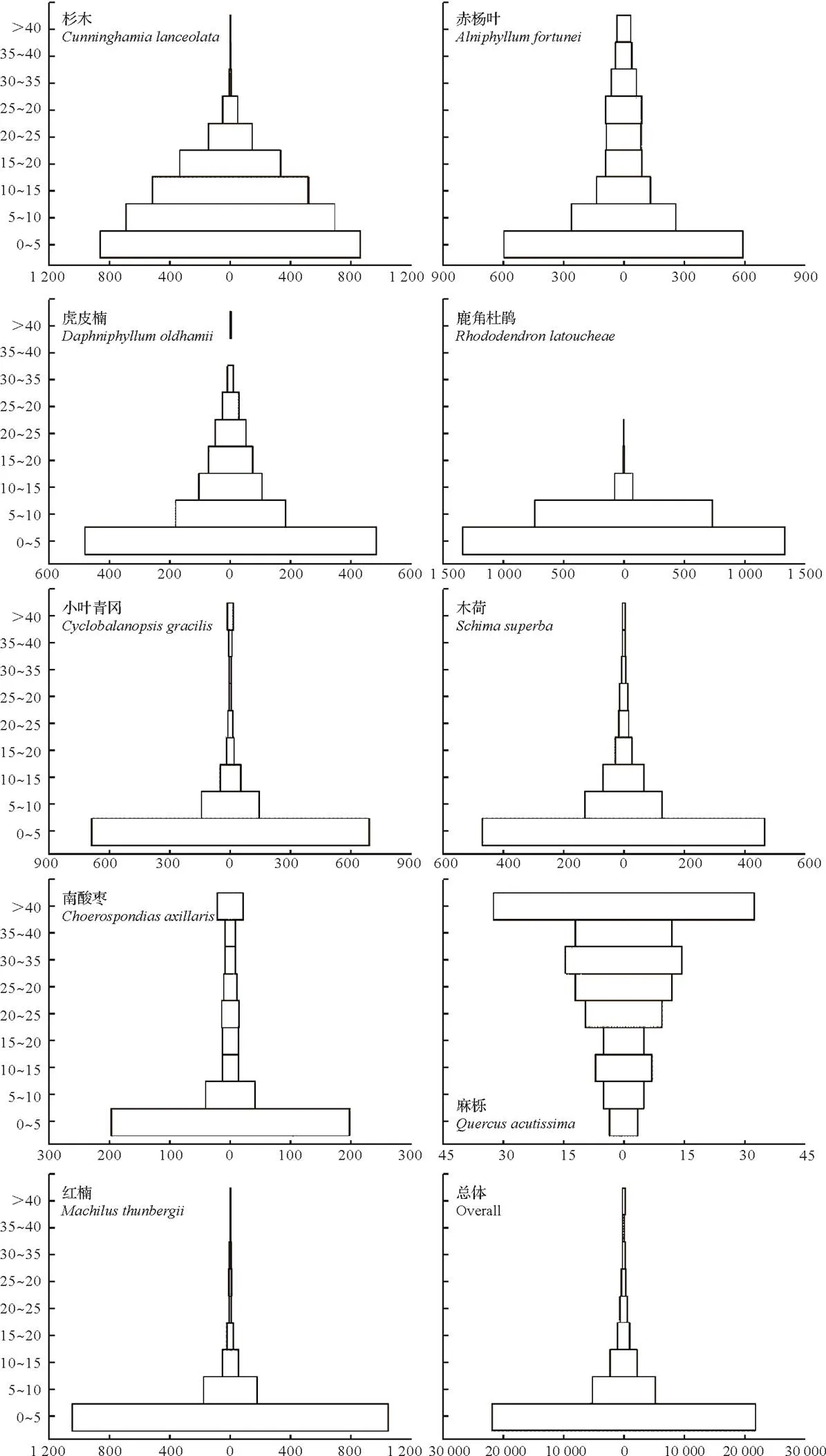
1.3.5径级结构种群的年龄结构是判断种群动态和群落演替的重要依据,但测量木本植物个体的年龄具有破坏性,因而在实际操作中往往采用胸径代替年龄的方法,来分析年龄结构[26]。本文将乔木层重要值位于前10并且年龄结构较为完整的9个物种[杉木、赤杨叶()、虎皮楠()、鹿角杜鹃()、小叶青冈()、木荷()、南酸枣()、麻栎()、红楠()]以及样地内所有个体划分为9个径级,第I径级DBH为0 cm 样地中木本植物种类丰富(表1),共有63 690株312种,隶属于65科139属(不包括未鉴定种)。其中,裸子植物较少,仅4科5属5种5 513株;被子植物有61科134属307种58 177株。从科的分布水平来看,样地内木本植物单种科、寡种科(2≤含种数<5)比例较高,共42科,占64.62%。科内种数大于等于10种的科只有8科,但种数高达140种,约占样地植物总种数的45%,其中蔷薇科(Rosaceae)和樟科(Lauraceae)的植物种类最丰富(25种);冬青科(Aquifoliaceae)和山茶科(Theaceae)次之(21种);壳斗科(Fagaceae)有15种。从属的分布水平来看,单种属和寡种属占据主体,其中,单种属共有83属,占样地木本植物总属的59.71%;寡种属(2≤含种数<5)有36属,占该区域木本植物总属数的25.90%;属内物种数超过10种的只有冬青属(21种)和柃木属(12种)。 表1 动态监测样地木本植物科内、属内种的组成 从种-多度格局来看:随着物种数的增加,个体数急剧增加,但当物种数达到32种时个体数增加的趋势减缓,达到86种时,个体数趋于稳定(图3)。其中,个体数<100株的物种数最多,占总种数的72.44%;株数≥1 000的有16种(35 972株,占56.48%),如:杉木(,5 225株)、鹿角杜鹃(,4 306株)、矩叶鼠刺(,3 674株)等。以个体数≤12株的物种为稀有种,统计结果显示样地中稀有种的比例很高,有148个物种,占总物种数的47.44%。其中,单株种的比例最高56种,占总物种数的17.95%,如巴东木莲()、白花泡桐()、厚皮香()等(表2)。 从种-面积曲线来看:当取样面积小于1 hm2,物种数随面积的增加急剧增加;当取样面积为4 hm2时,包含了官山样地中75%的物种(236种);当取样面积为9 hm2时,90%的物种(284种)出现。曲线(1)的拟合方程最佳,其模型:=0.046 8/(1+0.002),(=0.959 5)。 图A种-多度格局;图B种-面积曲线 表2 种内多度分布格局 通过对科重要值的分析发现,样地内科的优势现象明显(表3)。其中,壳斗科(Fagaceae)是样地内重要值(13.37%)最大的科,且远高于其他科,其个体数(8 582株)和胸高断面积(90.79 m2)均最大;樟科(Lauraceae)是样地内物种最丰富的科,有25个种,占总物种数的8.01%,其重要值仅次于壳斗科;排名第3的是安息香科(Styracaceae),其胸高断面积为59.38 m2,仅次于壳斗科;蔷薇科(Rosaceae)是样地中物种最丰富的科,重要值排名第7。重要值排名前10的科包含了样地中近40%的物种和70%的个体。 表3 官山动态监测样地重要值位于前10的科 样地中优势种明显(表4)。乔木层重要值排名前10的物种,株数占总株数的56.01%,胸高断面积占总数的57.04%,这些物种的重要值之和将近50.00%,重要值排名前3的物种为杉木(12.77)、赤杨叶(9.93)、毛竹(5.10);平均胸径最大的是南酸枣(24.05 cm)、麻栎(35.31 cm),但它们的个体数约占总株数的1%,重要值分别位于第8、9位。灌木层重要值排名前10的物种重要值占总物种的41.24%,鹿角杜鹃(7.09)、矩叶鼠刺(6.54)和杉木(5.46)分别位于重要值排名前三;鹿角杜鹃(3.50 cm)和杉木(3.60 cm)是平均胸径最大的物种;个体数最多的是矩叶鼠刺(3 462株);南酸枣和麻栎在灌木层的重要值则退出前10。 表4 官山动态监测样地内主要木本植物重要值 依照吴征镒先生对世界种子植物的分布区类型系统划分方法,官山大型森林监测样地木本植物139属可归并为15个分布区类型和13个变型,热带成分(2~7型)83属,温带成分(8~15型)52属,分别占总属数的59.71%、37.41%,其中中国特有分布型6属:山茶属()、红淡比属()、山茱萸属()、虎皮楠属()、木姜子属()、李属()。由此可以看出,官山样地植物区系具有热带和温带相互交融的特征,属于典型的亚热带地区(表5)。 表5 官山常绿阔叶林动态监测样地木本植物属分布型统计(含变型) 随着植物胸径的增大,样地内植物株数呈迅速减少的趋势,林木总体的径级结构遵从典型的金字塔型(图4)。其中胸径1~5 cm的个体数达43 747株,占总个体数的68.69%,占据主导地位;平均胸径为5.65 cm,胸径超过40 cm的个体数较少,共414株,占0.65%,其中胸径最大是南酸枣100 cm。可见低龄级的个体补给充足,老树个体较少,表明很长一段时间整个样地的物种将处于增长状态。 图4 主要物种的年龄结构 大部分优势种的径级结构呈金字塔型。其中虎皮楠、鹿角杜鹃、小叶青冈、木荷、红楠的径级分布属于典型的“金字塔”型,胸径(DBH)<5 cm的个体数分别占总个体数的79.26%、72.67%、63.22%、61.95%、51.88%,种群更新较好。杉木的径级分布虽然也遵从“金字塔”型,但每两个相邻径级间的个体数量相差不大(I径级33.33%,II径级)。赤杨叶、南酸枣虽然第I径级的个体数也是最多的,分别占总个体数的42.82%、40.53%,但当DBH>10 cm时赤杨叶和南酸枣,每两个相邻径级间的个体数相差不大,赤杨叶的径级分布属于“倒T”型,南酸枣属于种间小两端大的“沙漏”型,种群增长不稳定。麻栎的径级结构则呈“倒金字塔”型,其中DBH>40 cm的个体占总个体数达65株,占32.18%,DBH<5 cm的个体仅7株,占3.47%,该种群更新困难,已处于严重衰退状态。表明随着群落的演替赤杨叶、南酸枣、杉木也将渐渐退出群落。 江西官山常绿阔叶林动态监测样地物种丰富度高,共有木本植物63 690株312种139属65科,高于同处于亚热带地区其它样地[7,9-11,28-30],如:鼎湖山(20 hm2)210种119属56科、古田山(24 hm2)159种103属49科、天童山(20 hm2)152种94属51科。种-面积曲线分析结果进一步表明官山样地树木种类丰富、类型多样,当取样面积仅4 hm2时,就包含了236种木本植物。 官山样地的物种丰度高于同纬度的其它样地,其原因与人为干扰和群落演替阶段有关。本研究区域在1956—1957年间遭受人为砍伐和火烧,导致成体死亡形成林窗,为物种的生长和更新提供了空余生态位,有利于更多的物种迁入与定居,增加物种丰富度;根据中度干扰假说[31],干扰是调节植物群落多样性的主要因子,适度的人为干扰不仅能够提供更多的空余生态位,还能增加植物对资源利用的有效性,减弱种间的竞争作用,进而有利于物种多样性的维持。从群落的演替阶段来看,占据优势的树种既有演替先锋树种杉木、赤杨叶,又有相对稳定的顶级树种小叶青冈、红楠等,说明本研究样地植物群落处于干扰后自然恢复的中期阶段,而此阶段通常是物种多样性最高的阶段[32]。 此外,样地内物种丰富度高还与其稀有种比例有关,约占总物种数的一半(148种),远高于同处于亚热带地区的鼎湖山样地(110种)[9]、古田山样地(59种)[10]、天童山样地(55种)[33]。造成群落中稀有种较多的原因主要为:(1)受物种本身的种群特性和分布特征影响,样地中有些物种在自然界中种群数量稀少或分布区域较为零散,如:陀螺果()、榧树()、青钱柳()等。(2)由区系的交汇分布造成的,官山自然保护区处于华东植物区系与华中植物区系的交汇带,同时又是华南植物区系与华东植物区系的交汇带,相邻气候带的物种在分布上相互渗透,分布范围超出主要分布区域后,其个体数量将明显下降,如:穗花杉()、水青冈()、巴东木莲()等。 植物区系起源古老,单种属及寡种属占据优势,共有119属,占总属数的85.61%(表1)。寡种属、单种属的高百分比现象,一方面说明其自然环境复杂,另一方面也体现了植物区系起源古老[19],如:木兰科(Magnoliaceae)、胡桃科(Juglandaceae)、桑科(Moraceae)都是原始的科,穗花杉()、榧树()是第三纪孑遗植物。 样地植物区系具有从热带向温带过渡的特征,热带成分最多(59.71%),温带成分次之(37.41%),这与古田山样地(热带成分占53.9%,温带成分占43.1%)、天童山样地(热带成分占52.1%,温带成分占42.6%)研究结果相似[10-11],区别与鼎湖山样地以热带成分为主(89.90%),木林子样地[34]温带成分居多(62.50%)。这可能与研究区域的纬度和海拔高度有关,官山样地(444.1~645.0 m)与天童山样地(304.0~602.0 m)、古田山样地(446.3~714.3 m)的海拔较为接近且均属于中亚热带地区,相比之下鼎湖山样地的纬度更低属于南亚热带,且海拔更低(230~470 m),随着纬度和海拔的降低,热带成分比例增加;而木林子样地[35]的纬度更高属于北亚热带,且海拔也更高(1 588~1 780 m),随着纬度和海拔的升高,温带成分比例增加[36]。 径级结构是衡量植物群落稳定性和生长发育状况的重要指标[9]。从群落尺度来看,样地内所有个体的径级分布呈明显的金字塔型,其中胸径1~5 cm的个体数占总个体数的68.69%,说明样地中幼苗充足,林分结构合理,更新状态良好,群落处于稳定生长状态。 虽然群落尺度上物种更新良好,但部分先锋树种的种群已经处于衰退状态,如麻栎径级结构呈倒金字塔型,I、II径级(1~10 cm)个体比例低,III~IX径级(DBH>10 cm)的个体数多。可能受到了生物学特性和生态环境的综合影响,麻栎的种子属于坚果类,只能靠动物传播,传播的距离受到了限制,大部分种子留在了母株附近,而麻栎属于喜阳的先锋树种,幼苗的生长过程需要充足的光照,在母株的遮挡降下低了它的存活率[37];同时,随着群落的发展小叶青冈、红楠等常绿耐阴树种渐渐进入,林内郁闭度逐步增加,进一步抑制了喜阳物种麻栎的生长与更新,从而导致麻栎渐渐退出群落。这说明在群落的演替过程中,赤杨叶、南酸枣等阳性喜光树种也会被常绿耐阴树种逐步取代,从其径级分布可以得到证明,虽然目前赤杨叶和南酸枣的幼树个体充足,但壮树个体较少,在逐渐郁闭的林分下,幼树生长困难且补充量少,种群增长不稳定。 综上所述,官山常绿阔叶林动态监测样地木本植物种类丰富,稀有种比例高;植物区系地理成分多样,具有热带向温带过渡的特征,属于典型的亚热带气候区;且群落结构稳定,更新正常。但对植物组成、区系成分、径级结构等群落特征的分析仅为生物多样性的保护提供了基础数据,后期还需要进一步对其植物与植物之间、植物与群落环境之间的关系进行研究,阐明物种多样性的维持机制,从而更好地保护物种多样性。 [1] WANG X, GAO Q, WANG C, et al. Spatiotemporal patterns of vegetation phenology change and relationships with climate in the two transects of East China[J]. Global ecology and conservation, 2017, 10: 206-219. [2] 仲磊, 张杨家豪, 卢品, 等. 次生常绿阔叶林的群落结构与物种组成:基于浙江乌岩岭9 ha森林动态样地[J]. 生物多样性, 2015, 23(5): 619-629. [3] TILMAN, DAVID, REICH, et al. Biodiversity and ecosystem stability in a decade-long grassland experiment.[J]. Nature, 2006, 441(7093): 629-632. [4] SNOW A A, ANDOW D A, GEPTS P, et al. Genetically engineered organisms and the environment: current status and recommendations[J]. Ecological applications, 2005, 15(2): 377-404. [5] PEAT H J, CLARKE A, CONVEY P. ORIGINAL ARTICLE: Diversity and biogeography of the Antarctic flora[J]. Journal of biogeography, 2007, 34(1): 132-146. [6] DENG H B, JIANG M X, WU J W. Flora and ecological characteristics of rare plant communities on the southern slope of Shennongjia Mountain[J]. Journal of forestry research, 2002, 13(1): 21-24. [7] 温韩东, 林露湘, 杨洁, 等. 云南哀牢山中山湿性常绿阔叶林20 hm2动态样地的物种组成与群落结构[J]. 植物生态学报, 2018, 42(4): 419-429. [8] RICKLEFS R E. A comprehensive framework for global patterns in biodiversity[J]. Ecology letters, 2010, 7(1): 1-15. [9] 叶万辉, 曹洪麟, 黄忠良, 等. 鼎湖山南亚热带常绿阔叶林20公顷样地群落特征研究[J]. 植物生态学报, 2008(2): 274-286. [10] 祝燕, 赵谷风, 张俪文, 等. 古田山中亚热带常绿阔叶林动态监测样地: 群落组成与结构[J]. 植物生态学报, 2008(2): 262-273. [11] 杨庆松, 马遵平, 谢玉彬, 等. 浙江天童20 ha常绿阔叶林动态监测样地的群落特征[J]. 生物多样性, 2011, 19(2): 215-223. [12] Condit R, Ashton P, Bunyavejchewin S, et al. The importance of demographic niches to tree diversity[J]. Science, 2006, 313(5783): 98-101. [13] HOOPER E R, LEGENDRE P, CONDIT R. Factors affecting community composition of forest regeneration in deforested, abandoned land in Panama [J]. Ecology, 2008, 85(12): 3313-3326. [14] 朱教君, 刘世荣. 次生林概念与生态干扰度[J]. 生态学杂志, 2007(7): 1085-1093. [15] 姚良锦, 姚兰, 易咏梅, 等. 湖北七姊妹山亚热带常绿落叶阔叶混交林的物种组成和群落结构[J]. 生物多样性, 2017, 25(3): 275-284. [16] 卢刚, 沈爱华, 张健, 等. 浙南次生阔叶林群落结构特征研究[J]. 浙江林业科技, 2010, 30(1): 36-40. [17] 曹洪麟, 蔡锡安, 彭少麟, 等. 鹤山龙口村边次生常绿阔叶林群落分析[J]. 热带地理, 1999(4): 312-317. [18] 曹岚, 梁芳, 邹红, 等. 江西官山国家级自然保护区珍稀植物多样性研究[J]. 安徽农业科学, 2012, 40(3): 1696-1698. [19] 刘信中. 江西官山自然保护区科学考察与研究[M]. 北京: 中国林业出版社, 2005. [20] 兰国玉. 世界热带森林生态系统大样地定位研究进展[J]. 西北植物学报, 2007(10): 2140-2145. [21] 刘灿然, 马克平, 于顺利, 等. 北京东灵山地区植物群落多样性研究: 种-面积曲线的拟合与评价[J]. 植物生态学报, 1999(6): 490-500. [22] 李帅锋, 刘万德, 苏建荣, 等. 季风常绿阔叶林不同恢复阶段乔木优势种群生态位和种间联结[J]. 生态学杂志, 2011, 30(3): 508-515. [23] LINARES-PALOMINO R, ALVAREZ S. Tree community patterns in seasonally dry tropical forests in the Cerros de Amotape Cordillera, Tumbes, Peru[J]. Forest ecology & management, 2005, 209(3): 261-272. [24] 吴征镒, 周浙昆, 李德铢, 等. 世界种子植物科的分布区类型系统[J]. 云南植物研究, 2003(3): 245-257. [25] 吴征镒. 中国种子植物属的分布区类型[J]. 植物资源与环境学报, 1991(S4): 1-139. [26] 达良俊, 杨永川, 宋永昌. 浙江天童国家森林公园常绿阔叶林主要组成种的种群结构及更新类型[J]. 植物生态学报, 2004(3): 376-384. [27] 韩路, 王家强, 王海珍, 等. 塔里木河上游胡杨种群结构与动态[J]. 生态学报, 2014, 34(16): 4640-4651. [28] 姚兰, 崔国发, 易咏梅, 等. 湖北木林子保护区大样地的木本植物多样性[J]. 林业科学, 2016, 52(1): 1-9. [29] 秦运芝, 张佳鑫, 刘检明, 等. 湖南八大公山25 ha常绿落叶阔叶混交林动态监测样地群落组成与空间结构[J]. 生物多样性, 2018, 26(9): 1016-1022. [30] 孔令建, 游济顺, 俞社保, 等. 阳际峰自然保护区大样地群落结构特征[J]. 南方林业科学, 2016, 44(4): 5-8. [31] 文陇英, 李仲芳. 干扰对物种多样性维持机制的影响[J]. 西北师范大学学报(自然科学版), 2006(4): 87-91. [32] Pulsford S A, Lindenmayer D B, Driscoll D A. A succession of theories: purging redundancy from disturbance theory[J]. Biological reviews, 2016, 91(1): 148-167. [33] 孙小伟, 杨庆松, 刘何铭, 等. 基于浙江天童20 hm2常绿阔叶林动态监测样地的群丛划分[J]. 植物生态学报, 2018, 42(5): 550-561. [34] 姚兰. 湖北木林子保护区15 hm2大样地森林群落结构及多样性[D]. 北京: 北京林业大学, 2016. [35] 黄小, 姚兰, 艾训儒, 等. 木林子大样地两个木姜子属物种空间分布格局及关联性[J]. 广西植物, 2021, 41(8): 1333-1345. [36] 徐成东, 冯建孟, 王襄平, 等. 云南高黎贡山北段植物物种多样性的垂直分布格局[J]. 生态学杂志, 2008(3): 323-327. [37] 王霞, 胡海波, 张世豪, 等. 不同林龄麻栎林地下部分生物量与碳储量研究[J]. 生态学报, 2019, 39(22): 8556-8564. Species Composition and Community Structures of Subtropical Evergreen Broad-leaved Forests in Guanshan, Jiangxi Province, China WANG Qian1, YI Lingli1, CHEN Lin1, PENG Qiaohua1, OUYANG Yuanlan1, GONG Chao2, ZENG Xiaoxia2, YANG Qingpei2, SONG Qingni2, LIU Jun2* (1. Administration of Jiangxi Guanshan National Nature Reserve, Yichun, Jiangxi 336000, China; 2. Jiangxi Provincial Key Laboratory for Bamboo Germplasm Resources and Utilization, Jiangxi Agricultural University, Nanchang 330045, China) Evergreen broad-leaved forest is a zonal vegetation in subtropical areas of China. However, many primitive forests have become secondary forests,due to long-term human disturbance. In order to better monitor the restoration process of secondary evergreen broad-leaved forest in subtropical areas and reveal the mechanism of biodiversity maintenance.According to the construction standard of large-scale forest dynamic sample plot of Center for Tropical Forest Science (CTFS) of Smithsonian Institution, a 12 hm2secondary evergreen broad-leaved forest dynamic monitoring sample plot was established in Guanshan National Nature Reserve to investigate woody plants with DBH≥1 cm, and analyze their species composition, flora characteristics, diameter class structures and other community features.(1) The dynamic plot had rich species and rare species. There were 312 species in 65 families and 139 genera, of which 148 species were rare species, accounting for about half of the total number of species; (2) The flora and geographic components were diverse, with the nature of transition from tropical to temperate. There were 28 types (including variants) at the genus level. The tropical flora accounted for 59.71% of the total genera and the temperate flora accounted for 37.41%; (3) The diameter class distribution of the whole plant in the community was a “pyramid” type, and the forests were in a growing stage. The diameter class structure of most dominant species had the same trend.In summary, Guanshan secondary evergreen broad-leaved forest was rich in species composition, with a high proportion of rare species and good community regeneration. species richness; species-abundance pattern; species-area curve; flora characteristics; diameter class structure S718.54+2 A 2095-3704(2022)02-199-11 王倩, 易伶俐, 陈琳, 等. 江西官山亚热带次生常绿阔叶林物种组成与群落结构特征[J]. 生物灾害科学, 2022, 45(2): 199-209. 10.3969/j.issn.2095-3704.2022.02.36 2022-05-13 2022-06-06 财政部、环保部国家生物多样性野外监测示范基地改造项目(9131205274) 王倩(1974—),女,工程师,主要从事自然保护区管理工作,1575749864@qq.com;*通信作者:刘骏,副教授,博士,ljaim99@163.com。2 结果与分析
2.1 物种组成
2.2 种-多度格局与种-面积曲线
2.3 优势科与优势种
2.4 区系地理成分
2.5 径级结构
3 结论与讨论
3.1 物种组成
3.2 区系成分
3.3 径级结构
