不饱和脂肪酸瘤胃微生物氢化与调控奶牛泌乳性能的研究进展
2022-10-12普宣宣李秋爽谭支良
普宣宣,李秋爽,王 敏*,谭支良
(1.湖南农业大学动物科学技术学院,湖南长沙 410128;2.中国科学院亚热带农业生态研究所,湖南长沙 410125)
10 余年来,随着国家奶业振兴行动计划的实施,奶牛生产效率和单产水平显著提高。奶牛产后1~3周的能量摄入量通常低于产奶量的增长需求,易导致机体能量负平衡,产奶量越高,能量负平衡则越严重,产后4~8 周方可恢复能量正平衡,缓解能量负平衡对于维护奶牛健康和生产力至关重要。日粮中添加脂肪(酸)是缓解奶牛机体能量负平衡和维护奶牛健康的重要手段,并有助于提高泌乳性能。脂肪混合物中不饱和脂肪酸c9 C18:1 比例由10%增加到30%时,新产阶段奶牛体重和体况损失显著下降,干物质采食量呈上升趋势,并对奶牛后期的能量校正乳和乳脂有持续正向影响。日粮添加不饱和脂肪酸有助于提高乳脂不饱和脂肪酸含量,其经瘤胃生物氢化后有助于提高牛乳共轭亚油酸含量,进而有效提高牛乳营养价值。但部分瘤胃氢化中间产物对乳脂合成具有抑制作用。鉴于此,本文综述了不饱和脂肪酸瘤胃微生物氢化过程、氢化影响因子及不饱和脂肪酸对奶牛泌乳性能的影响等,旨在为利用不饱和脂肪酸提高奶牛生产效率和牛乳品质提供科学依据。
1 不饱和脂肪酸瘤胃微生物氢化
不饱和脂肪酸在瘤胃微生物作用下将碳链上的双键氢化成单键的过程,即为不饱和脂肪酸瘤胃微生物氢化,该现象于1951 年被首次发现。油酸、亚油酸和-亚麻酸的瘤胃生物氢化率分别为50%~80%、70%~95%和80%~100%。以油酸、亚油酸和-亚麻酸为例,其瘤胃生物氢化及其相关微生物作用过程分别见图1、图2 和图3。
1.1 不饱和脂肪酸瘤胃氢化的生物化学过程 油酸在瘤胃微生物的作用下加氢主要生成饱和脂肪酸C18:0,并伴随着反式脂肪酸t6、t7、t9、t10-t16 C18:1 等的生成(图1)。亚油酸在异构酶的作用下主要生成c9t11 C18:2,后在瘤胃微生物的作用下氢化为t11 C18:1,最后氢化为C18:0,并伴随着异构体t9t11、t9c12、c9c11、c10c12、t10c12 和t10t12 C18:2 等的生成,以及氢化中间产物t8-t10、t12、c10-c12 C18:1 等的生成(图2)。-亚麻酸在异构酶的作用下主要生成c9t11c15 C18:3,后在瘤胃微生物的作用下氢化为t11c15 C18:2,进一步氢化为t11 C18:1,最终氢化为C18:0,并伴随着异构体c9t13c15、t9t11c13、t10c12c15 和t9t11c15 C18:3 等的生成,以及氢化中间产物t11c13、t11t13、t13t15、t13c15、c12t12、t10c15、t11t14、c12c15、t12c15 C18:2 和t12-t16、c11、c12 及c15 C18:1 等的生成(图3)。另有研究发现,不饱和脂肪酸瘤胃生物氢化过程中有新的路径和中间产物生成,有待进一步挖掘和总结。

图1 油酸瘤胃生物氢化主要路径[12]

图2 亚油酸瘤胃生物氢化主要路径[12-14]
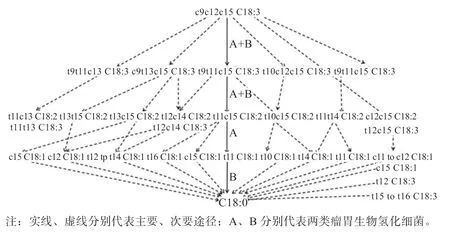
图3 α-亚麻酸瘤胃生物氢化主要路径[12-14]
1.2 不饱和脂肪酸瘤胃生物氢化的微生物过程 瘤胃细菌和原虫是参与不饱和脂肪酸生物氢化的主要微生物。根据氢化产物的不同,可将瘤胃氢化细菌分为A、B 两类。A 类细菌可氢化不饱和脂肪酸的最终产物为反式单不饱和脂肪酸,包括丁酸弧菌属、密螺旋体属、微球菌属、真细菌、真细菌F2/2、真细菌W461、白色瘤胃球菌F2/6、革兰氏阴性杆菌EC7/2、革兰氏阴性杆菌R8/3 及革兰氏阴性弧菌2/9/1 等。A 类细菌中溶纤维丁酸弧菌A38、密螺旋体属主要转化亚油酸为t11 C18:1、c9 C18:1、c11 C18:1 和c9t11 C18:2;溶纤维丁酸弧菌S2、微球菌属及白色瘤胃球菌F2/6 主要转化亚油酸和-亚麻酸为反式C18:1 和顺式C18:1;真细菌F2/2 主要氢化亚油酸为反式C18:1,转化-亚麻酸为t11 C18:1、t11c15 C18:2 和c9t11c15 C18:3;真 细菌W461 主要氢化亚油酸为C18:1,氢化-亚麻酸为顺式和反式C18:1;革兰氏阴性杆菌R8/3 主要氢化亚油酸和-亚麻酸分别为t11 C18:1 和t11c15 C18:2;革兰氏阴性杆菌EC7/2 主要氢化亚油酸为反式C18:1,氢 化-亚麻酸为t11c15 C18:2 和t11 C18:1。B类细菌可氢化不饱和脂肪酸的最终产物为饱和脂肪酸C18:0,包括、、革兰氏阴性杆菌R8/5、丁酸产生菌和等。B 类细菌中主要氢化油酸为C18:0,氢化亚油酸为t11 C18:1 和C18:0,氢化-亚麻酸为t11c15 C18:2 和c15 C18:1;Fusocillus T344 主要转化油酸为t11 C18:1、c9 C18:1 和C18:0,氢化亚油酸为t11 C18:1和C18:0,氢化-亚麻酸为c9t11 C18:2 和c15 C18:1;革兰氏阴性杆菌R8/5 主要转化油酸为c9 C18:1 和C18:0,氢化亚油酸为t11 C18:1 和C18:0,氢化-亚麻酸为t11c15 C18:2、c15 C18:1、t15 18:1 和C18:0。
原虫的不饱和脂肪酸含量显著高于细菌,因此原虫在脂肪酸生物氢化中可能起着一定作用,但尚未完全清楚。有研究表明,原虫丰度与瘤胃t11 C18:1 含量呈正相关,与t10 C18:1 及瘤胃生物氢化完整性呈负相关,原虫可能抑制了不饱和脂肪酸氢化为饱和脂肪酸的最后一步。Or-Rashid 等研究发现,原虫和细菌混合可以改变亚油酸的生物氢化路径,这也可能与原虫对细菌的吞噬作用相关。今后可通过进一步试验探究原虫与不饱和脂肪酸间的联系,为更好地调控脂肪酸瘤胃微生物氢化提供参考依据。
1.3 不饱和脂肪酸瘤胃生物氢化的影响因素
1.3.1 不饱和脂肪酸碳链长短、浓度和添加形式 瘤胃生物氢化受不饱和脂肪酸碳链长短、含量和添加形式等影响。William发现,不饱和脂肪酸碳链长短促进氢化效率的顺序为C18:3>C16:1>C18:2>反式C18:1>顺式C18:1。不饱和脂肪酸生物氢化效率与其浓度相关,低pH 发酵液中,亚油酸的生物氢化率随玉米油添加浓度的升高而下降。不饱和脂肪酸的添加形式对其瘤胃生物氢化率有显著影响,以钙盐或甲醛形式处理后,进入瘤胃的游离不饱和脂肪酸含量降低,进而降低了不饱和脂肪酸的瘤胃生物氢化率。与大豆油相比,添加整粒大豆显著降低了亚油酸的生物氢化率,提高了c9t11 C18:2 的生成量。与菜籽油相比,日粮中添加整粒菜籽可显著提高乳脂中不饱和脂肪酸含量。
1.3.2 日粮结构与组成 精粗比是影响不饱和脂肪酸瘤胃生物氢化的主要日粮因素。随精料比例的增加,不饱和脂肪酸瘤胃生物氢化率降低。当日粮精料比例大于70%时,亚油酸和-亚麻酸的生物氢化率分别降低至35%~60%和50%~80%,这与瘤胃发酵模式向丙酸型转变相关,促进了氢气的利用,进而抑制不饱和脂肪酸瘤胃生物氢化。不饱和脂肪酸和精料比例过高易导致氢化路径由t11 C18:1 向t10 C18:1 转变。另外,日粮精粗比增加也会引起瘤胃pH 下降。低pH 抑制脂肪酶和异构酶活性,进而降低生物氢化效率。体外法研究结果表明,低pH 易导致发酵液t10c12 C18:2 含量增加。日粮种类对脂肪酸瘤胃生物氢化也具有一定影响。新鲜牧草富含-亚麻酸,可降低瘤胃不饱和脂肪酸生物氢化率,进而提高放牧奶牛瘤胃中t11 C18:1含量。
除上述因素外,不饱和脂肪酸间的相互作用也可影响其瘤胃生物氢化率。超长链不饱和脂肪酸可抑制亚油酸和-亚麻酸生物氢化为饱和脂肪酸的最后一步,促进反式不饱和脂肪酸和共轭亚油酸的生成。
2 不饱和脂肪酸对奶牛泌乳性能的影响
2.1 不饱和脂肪酸对产奶量的影响 富含不饱和脂肪酸的籽实(油)可提高日粮能量水平,进而有助于提高奶牛产奶量。皮宇研究表明,日粮中添加富含-亚麻酸的橡胶籽油、亚麻籽油及两者混合物均显著提高了奶牛产奶量。每增加100 g 膨化亚麻籽可增加奶牛产奶量0.11~0.14 kg/d。6.5%膨化亚麻籽可使奶牛产奶量增加12.3%。DHA 可使头胎奶牛和多胎奶牛的产奶量分别增加1.2 kg/d 和0.5 kg/d。日粮添加菜籽和菜籽油可使奶牛产奶量分别增加0.8 kg/d 和1.0 kg/d。由此可知,富含不饱和脂肪酸籽实(油)可有效提高奶牛产奶量,作用效果与添加的不饱和脂肪酸种类和浓度及奶牛泌乳胎次等相关。
2.2 不饱和脂肪酸对乳脂合成的影响 不饱和脂肪酸对乳脂合成的影响效果不一。部分研究表明,添加整粒富含不饱和脂肪酸的籽实可提高乳脂率和乳脂含量,但进入瘤胃的游离不饱和脂肪酸含量较高时易抑制乳脂合成。乳脂抑制主要指乳脂肪含量降低10%~50%,同时未出现产奶量或其他乳成分含量降低的现象。不饱和脂肪酸对乳脂合成的抑制作用主要在于其对瘤胃发酵模式的影响及其生物氢化中间产物。与改变瘤胃发酵模式相比,瘤胃生物氢化中间产物(如t10c12 C18:2、t10 C18:1 和c10t12 C18:2 等)更易抑制乳脂合成。t10c12 C18:2 与乳脂含量呈负相关,对奶牛进行瘤胃灌注、静脉注射或十二指肠灌注t10c12 C18:2 时,乳脂浓度和产量均显著降低,该现象常伴随着乳腺中相关乳脂合成酶编码基因表达量的降低,如乳腺转录因子固醇调节元件结合蛋白()、脂肪酸结合蛋白基因4()、乙酰羧化酶编码基因()及脂肪合成酶编码基因()等。但Ventto 等研究表明,奶牛乳脂含量降低时,瘤胃中t10c12 C18:2含量并未增加。这可能与t10c12 C18:2 在乳脂中的含量较少相关,可能有其他瘤胃氢化中间产物共同参与了对乳脂合成的抑制作用。脂肪酸瘤胃生物氢化通路由t11 C18:1 向t10 C18:1 转变是产生乳脂抑制现象的特征之一。t10 C18:1 对乳脂合成酶编码基因和等具有抑制作用,这是其抑制乳脂合成的主要原因。而针对母羊乳脂降低现象的相关数据分析中,t10 C18:1 未起到主要作用,这可能主要与乳脂中t10 C18:1 含量较少相关。c10t12 C18:2 与乳脂抑制也具有一定联系。Saebø 等研究发现,c10t12 C18:2 具有抗脂肪生成作用。综上,不饱和脂肪酸瘤胃生物氢化中间产物对乳脂抑制现象的影响与其浓度相关,可通过调控其瘤胃生物氢化通路及上述中间产物生成量,进而减少对乳脂合成的抑制作用。除上述因素外,某些尚未明确的氢化中间产物也与乳脂降低相关,可进一步探究不饱和脂肪酸瘤胃氢化通路和氢化中间产物与乳腺合成乳脂间的联系,为更好地调控奶牛乳脂组成提供理论依据。
乳脂合成还受脂肪酸和日粮组合效应、脂肪酸添加形式和促乳脂生成物质等调控。在低纤维日粮中添加高剂量的不饱和脂肪酸易抑制乳脂合成,高纤维日粮则促进不饱和脂肪酸氢化为饱和脂肪酸,降低对乳脂合成的抑制。富含不饱和脂肪酸整粒籽实可降低不饱和脂肪酸对乳脂合成的抑制作用。促乳脂合成物质(如乙酸盐)可以显著提高乳脂含量,因此可通过添加乙酸盐和不饱和脂肪酸混合物来提高乳脂含量和改善乳脂品质。另外,实际生产中应注意不饱和脂肪酸种类、添加形式的选取,并根据实际情况添加适宜浓度不饱和脂肪酸以平衡乳脂和乳脂肪酸间的协同性。
2.3 不饱和脂肪酸对乳脂肪酸组成的影响 不饱和脂肪酸含有多种生物活性因子,有利于降低人体血液甘油三酯和胆固醇含量,提高机体免疫力。实际牛乳脂不饱和脂肪酸占比(23%~35%)显著低于“理想”不饱和脂肪酸(92%)。添加富含不饱和脂肪酸的籽实(油)则可提高进入乳腺的不饱和脂肪酸含量,进而提高牛乳中不饱和脂肪酸含量。不饱和脂肪酸及其氢化中间产物对乳腺乙酰羧化酶的转录和活性具有抑制作用,因此可抑制乳腺利用乙酸合成饱和脂肪酸,进而降低饱和脂肪酸含量。DHA(俗称脑黄金)为人体必需脂肪酸之一,可促进神经生长和发育,并具有抗癌作用。牛乳为人体DHA 主要来源之一,添加鱼油、微藻及DHA 等可有效提高乳脂中DHA 含量。共轭亚油酸具有抗癌、调节免疫功能、预防动脉硬化和促进人体新陈代谢等作用。c9t11 C18:2 占牛乳总共轭亚油酸含量的90%以上,主要由不饱和脂肪酸的瘤胃氢化中间产物t11 C18:1 经乳腺脱氢合成。因此,添加不饱和脂肪酸可有效提高牛乳共轭亚油酸含量。日粮中添加2%鱼油,乳脂中t11 C18:1 和c9t11 C18:2 的含量可分别增加90%和83%。富含不饱和脂肪酸的籽实(油)种类与其对乳脂共轭亚油酸含量的影响相关。添加等量的棉籽油、玉米油和大豆油时,乳脂共轭亚油酸含量分别由对照组的3.5 mg/g 分别增加到6.0、7.1、10.3 mg/g。添加等量的大豆油和亚麻籽油时,乳脂中共轭亚油酸含量分别增加1.51%和0.79%。综上,日粮中添加不饱和脂肪酸可有效改善乳脂肪酸组成,提高牛乳营养价值。
3 小结和展望
添加富含不饱和脂肪酸籽实(油)可提高日粮能量水平,对维护奶牛健康、提高产奶量和改善乳品质均具有正向作用,但部分瘤胃氢化中间产物t10c12 C18:2、t10 C18:1 及c10t12 C18:2 等对乳脂合成具有抑制作用。今后应加强对不饱和脂肪酸瘤胃微生物氢化的调控,提高氢化中间产物t11 C18:1 和c9t11 C18:2 的生成量,进而更有效地提高牛乳品质。另外,应加强不饱和脂肪酸与基础日粮组合效应、促乳脂生成物质与不饱和脂肪酸的组合效应等方面的研究,进而平衡乳脂合成和乳品质间的协同性,为不饱和脂肪酸更好地应用于奶牛生产中提供参考依据。
