不同种源高山栲光合日变化及光合潜能分析
2022-10-10王顺利张春华戚建华毛晓佩孙永玉
王顺利,张春华,戚建华,毛晓佩,孙永玉
(1.西南林业大学,云南 昆明 650233;2.中国林业科学研究院高原林业研究所,云南 昆明 650233,3.国家林业局云南元谋荒漠生态系统定位研究站,云南 昆明 650233)
高山栲(Castanopsisdelavayi)又名滇锥栗,为壳斗科(Fagaceae)栲属(Castanopsis)常绿乔木树种,主要分布在我国云南、四川和贵州海拔1500~2800 m的山地[1]。其木材坚久耐用,可供薪碳、建筑、枕木、车辆等用材;其果可食,根、茎皮、果实等可入药[2]。高山栲是我国西南部半湿润常绿阔叶林的主要优势树种之一,也是滇中高原和川西南山地区域的地带性顶级群落,作为亚热带壳斗科栲属中重要建群种,形成了中国特有的森林类型[3-5]。同时,高山栲也是生态公益林建设的优良树种,在水土保持、水源涵养和调节气候等生态服务功能方面具有重要作用。但由于受人类活动的干扰,现存林分多为残留的斑块状森林,林下自然更新状况较差。因此,开展不同种源高山栲的光合特性研究,筛选出光合效率高、光合潜力大的高山栲种源,对高山栲植树造林及生态恢复和生态保护等具有重要意义。
1 材料与方法
1.1 研究材料
试验点位于云南省昆明市中国林业科学研究院高原林业研究所(25.04°N、102.45′E),海拔1891 m,年均降水量约为1011.8 mm,年均气温14.7 ℃,年均日照时间2200 h。依据高山栲实际地理分布,参考中国数字植物标本馆中相关标本采集的地理信息,确定高山栲核心分布区在滇中高原附近。本次供试的高山栲幼苗种源来自于核心分布区域内的5个县域,分别为:楚雄双柏(SB)、大理宾川(BC)、临沧永德(YD)、普洱镇沅(ZY)、保山龙陵(LL)。各种源地基本信息详见表1。
于2020年7月,按照随机原则,选取5个种源的1年生高山栲实生苗(幼苗枝叶健康,无明显的损伤和病虫害痕迹)各30株,带回试验地,使用规格为26 cm×20 cm×17 cm的容器进行盆栽种植。盆土统一使用试验地内基础养分一致的红土生土(pH值7.0、有机质含量4.77 g·kg-1、全N含量1.87 g·kg-1、全磷含量1.06 g·kg-1、全钾含量8.34 g·kg-1)。种植后置于露天环境下,每2 d浇水1次,定期除草(不施肥)。培育至2020年9月,幼苗生长状况稳定开始测定各类指标。

表1 高山栲种源采集地基本信息
1.2 研究方法
1.2.1 光合日变化测定 于2020年9月,选择晴朗天气,利用Li-6400便携式光合仪(Li-Cor,Lincolin,NE,USA)测定5个种源高山栲幼苗的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、蒸腾速率(Tr)、光合有效辐射(PAR)等指标,并计算光能利用效率(SUE=Pn/PAR) 和水分利用效率(WUE=Pn/Tr)。从每个高山栲种源中选择长势基本一致的苗木5株,每株随机选择2~3片叶进行光合测定,测定时间为8∶00—18∶00,每隔2 h测定1次,每片叶重复3次,连续测定2 d,结果取平均值。测定时CO2浓度、湿度和温度均为自然状态。
1.2.2 光响应曲线测定 于2020年9月,选择晴朗天气,利用Li-6400便携式光合仪光响应曲线测定程序测定5个种源高山栲幼苗在不同光强下的净光合速率(Pn)。为避免发生中午强光抑制现象,测定时段控制在8∶30—11∶30,光合有效辐射梯度依次设定为1600、1200、1000、800、600、400、300、200、150、100、50、0 μmol·m-2·s-1。每个光强下测定120 s,每片叶重复3次,每个时段测定2 d,结果取平均值。
1.3 数据处理
数据采用Excel 2010、SPSS 25.0(IBM SPSS,USA,NY)进行统计分析及相关性分析,采用Origin作图。
2 结果与分析
2.1 不同种源高山栲幼苗光合日变化特性
研究结果表明,高山栲5个地理种源的叶片净光合速率日变化曲线呈双峰型(图1),峰值出现在10∶00和16∶00,在2个峰值之间,即14∶00时的净光合速率出现谷值,说明高山栲幼苗存在光合午休现象。5个种源的净光合速率都在10∶00时达到一天中的最大值,此时净光合速率从大到小依次为ZY(5.87 μmol·m-2·s-1)>LL(5.24 μmol·m-2·s-1)>SB(4.61 μmol·m-2·s-1)>YD(4.47 μmol·m-2·s-1)>BC(4.38 μmol·m-2·s-1)。5个种源的净光合速率都在18∶00时达到一天中的最小值,此时净光合速率从大到小依次为LL(1.52 μmol·m-2·s-1)>ZY(1.46 μmol·m-2·s-1)>BC(1.02 μmol·m-2·s-1)>SB(0.86 μmol·m-2·s-1)>YD(0.54 μmol·m-2·s-1)。而日均光合速率从大到小排序为LL(3.66 μmol·m-2·s-1)>ZY(3.36 μmol·m-2·s-1)>SB(2.85 μmol·m-2·s-1)>YD(2.84 μmol·m-2·s-1)>BC(2.56 μmol·m-2·s-1)。
高山栲气孔导度与光合速率变化趋势基本一致,在正午前较高,午后一直维持在较低水平。不同种源高山栲叶片气孔导度呈单峰型变化趋势,5个种源气孔导度均在10∶00达到峰值,随后呈逐渐降低的趋势,最终于18∶00达到最小值。在12∶00—14∶00间,SB种源气孔导度及净光合速率下降幅度最大,同时其胞间CO2浓度上升与其它种源表现出明显差异,这从侧面说明不同种源高山栲经过引种培育后在同一环境条件下,为适应环境而做出了不同的生理响应。不同种源高山栲胞间CO2浓度变化过程有所不同,但都在18∶00时达到最大值,LL、ZY、YD最低值出现在14∶00,BC、SB最低值分别出现在12∶00、16∶00。蒸腾速率变化趋势与气孔导度变化相似,即呈单峰变化,除YD种源在14∶00时最高,其余种源均在10∶00达到最大值,随后逐渐降低,18∶00时达到最小值。总体来看,10∶00时高山栲呼吸作用较强,水分消耗较多,蒸腾速率较快,净光合速率较高,此时段可能是高山栲根系生长的重要时段。
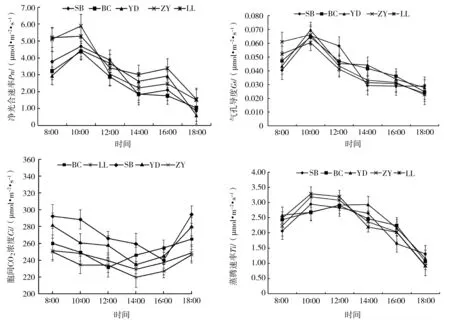
图1 不同种源高山栲光合参数日变化
2.2 不同种源高山栲幼苗光响应特性
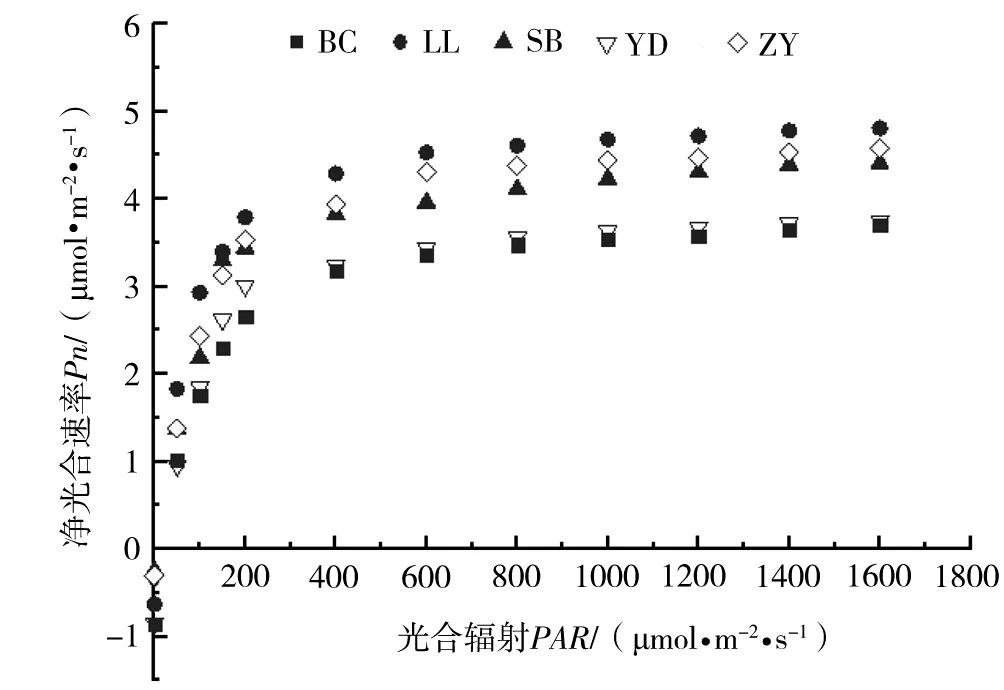
图2 不同种源高山栲光响应曲线(Pn-PAR)
随着光合有效辐射的增加,各地理种源的净光合速率都呈现出上升趋势(图2)。各种源在光强低于800 μmol·m-2·s-1时,随着光合有效辐射的增加,苗木的净光合速率急剧增加;当光强增加至800 μmol·m-2·s-1以上时,随着光合有效辐射的增加,苗木的净光合速率变化不大。这说明不同地理种源高山栲苗木的光饱和点为800 μmol·m-2·s-1,且为阳性树种。光强800 μmol·m-2·s-1时,净光合速率LL种源最大,为4.61 μmol·m-2·s-1;其次为ZY种源(4.38 μmol·m-2·s-1)、SB种源(4.11 μmol·m-2·s-1)、YD种源(3.56 μmol·m-2·s-1);BC种源最小,仅3.48 μmol·m-2·s-1。由此得出,高山栲能应用采伐迹地、裸地环境造林,对强日照有较好的适应能力,其中LL种源的适应性最强。
2.3 不同种源高山栲幼苗光能利用率日变化
不同种源高山栲苗木的光能利用率日变化总体呈先下降后上升的趋势。8∶00—14∶00,不同种源苗木的光能利用率逐渐下降,在12∶00时5个种源的光能利用效率较为接近;14∶00后不同种源高山栲苗木的光能利用率开始上升,并在18∶00时达到最高(图3)。在5个种源中,LL种源的光能利用率最大,为0.022 mol·mol-1,且光能利用率日平均值为0.0056 mol·mol-1,二者显著高于其它4个种源。这说明在相同自然条件下,LL种源对光照强度反应的敏感程度高于其它几个种源,对强光的适应能力较好,有较宽的适应性,既能充分利用弱光进行光合作用,也能对强光有较高的利用能力,光照生态幅较宽,相比而言其潜在的光合作用较强。

图3 不同种源高山栲光能利用率日变化图4 不同种源高山栲水分利用率日变化
2.4 不同种源高山栲幼苗水分利用率日变化
不同种源高山栲苗木的水分利用率日变化复杂,但总体呈先下降后上升的趋势。10∶00—12∶00各种源水分利用率逐渐下降,14∶00—16∶00不同种源高山栲苗木的水分利用率开始上升(图4),其中LL种源的水分利用率在8∶00及18∶00时都处在较高水平,分别为2.03、1.68 mol·mol-1。这表明在弱光条件下LL种源的水分利用率更高,且在5个种源中,LL种源日均水分利用率也最高,为1.55 mol·mol-1,显著高于其它4个种源。这说明在相同自然条件下,LL种源在水分利用方面最为优异。
2.5 高山栲光合指标的相关性

表2 高山栲幼苗光合指标相关分析
由表2可知,高山栲幼苗叶片的Pn与Gs呈极显著正相关(R=0.990),与Ci、Ta呈现负相关但不显著,相关系数分别为-0.532、-0.180,而与Tr、PAR的相关系数分别为0.646、0.369。这说明气孔导度增加能显著提高叶片净光合速率,而Ci、Ta、Tr、PAR对高山栲Pn影响不大。另外,Tr与PAR间的相关系数为0.925,呈现出极显著正相关,说明有效光合辐射强弱是高山栲叶片蒸腾速率的决定因子,且蒸腾速率随有效光合辐射的增加而显著增强。其它因子间的相关系数介于-0.538~0.704之间,无显著相关关系。
3 讨论
幼苗时期是植物对环境变化反应最为敏感的阶段,其生长规律由遗传特性在环境条件下的适应性所决定[6-7],光作为植物生长、发育最重要的环境因子之一,在一定程度上能够对植物的功能性状产生影响,与生长发育阶段和环境因子密切相关[8-11]。许大全[12]认为,当植物“午休”现象严重时,光合日进程中没有下午的第2个峰值出现,其峰值不是在中午,而是在上午的早些时候。本研究中各种源高山栲光合日进程变化为“双峰”型,不同于典型的“单峰”型,这表明高山栲在该时期“午休”现象不严重。Farquhar等[13]将植物“午休”原因归结为气孔限制型和非气孔限制型。气孔限制型主要表现为气孔导度和胞间CO2浓度同步降低。本试验中,10∶00—14∶00高山栲气孔导度、胞间CO2浓度和光合速率同步下降,说明在此时间段高山栲的光合“午休”主要是气孔限制型。此时段由于受外界条件影响,太阳光照强烈,高山栲体内水分供应紧张,空气湿度低,引起气孔部分关闭,植物体内CO2含量降低造成了光合作用的减弱,因此在夏季高温时,可通过适时喷灌,促进气孔开放和蒸腾作用,以维持高山栲较强的光合性能。
5个种源高山栲的光饱和点为800 μmol·m-2·s-1,处于一般木本植物阳生叶片的光饱和点范围内(600~1500 μmol·m-2s-1)[14]这表明高山栲在高光和有效辐射下光合作用能力较强,该结果与其光合日变化进程中净光合速率在上午10:00出现最大值的表现一致,这与闭鸿雁等[15]研究结果一致,可以从侧面反映出高山栲属阳性树种,能够适应较强的光。
李理渊等[16]研究表明,7—9月在低光强下,栓皮栎林内叶片对弱光的利用能力较强,本研究结果与之相似,9月各种源高山栲对弱光利用能力较强,因此在高山栲种植过程中可使用遮荫网遮阴,或适当增加栽培密度使其获得更大的光合积累。谢会成等[17]研究表明,栓皮栎在生长盛期(7—10月)时水分利用效率在上午和下午时较高,中午时较低。本试验中,各种源高山栲水分利用效率在8∶00—10∶00和16∶00—18∶00时段较高,这与前人研究结果一致,可能是由于栓皮栎与高山栲为同属植物,在生理上具有一定相似性。
本试验中,LL种源高山栲在试验地生境条件下经过光照适应,具有较快的净光合速率。为减少强光照辐射带来的高温影响,高山栲叶片通过调整气孔开闭来调控蒸腾速率的大小和胞间CO2浓度的高低,以平衡叶片温度,从而减少强光辐射带来的高温伤害和生理干旱,实现净光合速率的最大化,这反映出LL种源较高的环境适应力。不同种源高山栲的光响应参数变化趋于一致,但在相同光饱和点参数下,LL种源Pn值显著高于其它几个种源,表明LL种源能够更好地利用弱光积累更多的有机物质,这在一定程度上能够促进LL种源对环境适应的调节能力,从侧面反映出LL种源具有更强的光合潜能。对比分析发现,在相同生境条件下,LL种源对光照强度反应的敏感程度及水分利用效率均高于其它几个种源,对强光的适应能力较好,有较宽的适应性,既能充分利用弱光进行光合作用,也能对强光有较高的利用能力,光照生态幅较宽,相比而言其潜在的光合作用较强。
4 结论
1)不同种源高山栲光合日变化趋势较为一致,随着光照辐射的增加,净光合速率上升,均呈双峰型曲线,表现出光合午休现象。光合速率总体表现为LL>ZY>SB>YD>BC。不同种源高山栲叶片对光合有效辐射的响应与Pn变化趋势基本一致,即随着光合有效辐射的增加Pn逐渐升高。当光合有效辐射达到800 μmol·m-2·s-1时,各种源光合速率排序为LL>ZY>SB>YD>BC。
2)各种源高山栲光合利用率均在18:00时到达最大值;不同种源间水分利用效率变化复杂,各种源均在8∶00时水分利用效率更高,相比而言LL种源日均光能利用率、日均水分利用率都较其它几个种源要高。
3)综合比较来看,在相同光照条件下LL种源净光合速率更快,能够适应更广泛的光,并且有较强的光能利用率和水分利用率,光合效率高,光合潜能大,可作为高山栲繁育的优良种源。
