干旱胁迫对夹竹桃光合特性的影响*
2022-10-08陈慧英孙元敏李和阳丁伟伦汤坤贤
张 飞 陈慧英 陈 珊,3,4,5 孙元敏 李和阳 马 勇 丁伟伦 汤坤贤**
(1.自然资源部第三海洋研究所,福建 厦门 361005;2.福建省海洋生态保护与修复重点实验室,福建 厦门 361005;3.自然资源部海洋生态保护与修复重点实验室,福建 厦门 361005;4.漳州海岛海岸带福建省野外科学观测与研究站,福建 厦门 361005;5.自然资源部海峡西岸海岛海岸带生态系统野外科学观测与研究站,福建 厦门 361005)
在植物生长过程中,干旱是其经常面临的胁迫之一[1],尤其在淡水资源缺乏的海岛环境中,植被的修复很大程度上取决于水分的获取[2]。在干旱胁迫下,植物一般会通过降低气孔开度或关闭气孔来减少蒸腾作用带来的水分流失[3]。进一步的干旱会导致植物叶肉细胞损伤、光合酶的活性下降[4],同时还会损害叶绿体的结构,导致叶绿素含量降低[5],进而影响植物对光能的吸收和利用。夹竹桃(Nerium indicum)又名笔桃、柳叶桃,属夹竹桃属,为常绿灌木或小乔木,常被用作园林绿化和观赏[6]。夹竹桃适应性强,具有良好的抗旱及护坡能力,可作为海岛植被修复的备选植物[7,8]。干旱胁迫可造成夹竹桃叶片卷曲、枝叶干枯,生理指标上则表现为细胞通透性增加、丙二醛和脯氨酸含量升高[9]。目前干旱胁迫对于夹竹桃光合作用的影响尚未见报道,研究干旱胁迫对夹竹桃光合特性的影响,对于揭示该植物对于干旱环境的响应规律及适应机制具有重要意义,并可为海岛、海岸等区域的植被修复及后续管理提供参考。
1 材料与方法
1.1 实验材料
夹竹桃采用长势良好的2年生苗,来自于福建塘屿附近园林苗圃。
1.2 实验方法
1.2.1 干旱实验处理
参考汤坤贤等的方法[9],于2015年11月至2016年4月,在自然资源部第三海洋研究所漳州古雷基地可通风透明的塑料大棚实验区内开展干旱实验。相比外部环境,大棚内光照强度约为2/3,温度略高,湿度差异不大;夹竹桃种植于直径为30 cm,高35 cm的塑料花盆中;盆栽基质采用沙土比例约为1∶1的土壤,花盆内基质填装高度约为30 cm高。选取长势良好且较一致的夹竹桃植株20株,平均分为对照组和干旱组,每7天为1周期,共持续17周期。在实验期间,对照组每周期浇1次水,每次1 L;干旱组前11个周期保持不浇水操作,后6个周期每次复水1次(充分浇水使土壤水分达到饱和)。
1.2.2 植物叶片光和参数的测定
在晴朗天气的同一时间段,选取对照组和干旱组中各3株植株的中段叶片,使用Li-6400XT便携式光合系统测定仪进行光和参数的测定。每组测定3个叶片,每片3个重复。在设定光强500 μmol/m2·s下测定所取叶片的净光合速率(Pn)、蒸腾速率(Tr)、水分利用率(WUE)、气孔导度(Gs)和胞间CO2浓度(Ci)。
1.3 数据收集及分析
各指标使用Excel和SPSS13.0对数据处理,并对所获数据进行单因素方差分析以及pearson相关性分析,用Origin 8.0作图。
2 结果与分析
2.1 干旱胁迫对夹竹桃净光合速率(Pn)的影响
由图1所示,对照组夹竹桃的净光合速率为(1.39±1.58)μmol CO2/m2·s,干旱胁迫下,夹竹桃的净光合速率为(-1.57±0.57)μmol CO2/m2·s。水分亏缺条件下的植株净光合速率下降,与对照组的变化差异极显著(P<0.01)。当净光合速率降为负值,这表明植株的光合作用强度小于呼吸作用强度;进行复水操作给予植株充足水分之后,对照组夹竹桃的净光合速率为(13.25±0.87)μmol CO2/m2·s,干旱胁迫下夹竹桃的净光合速率为(11.02±3.11)μmol CO2/m2·s,此时对照组与干旱组植株的净光合速率都得到了提升,组间差异不显著(P>0.05),说明复水后夹竹桃的光合系统得到了较好的恢复。

图1 复水前后夹竹桃叶片净光合速率
2.2 干旱胁迫对夹竹桃蒸腾速率(Tr)的影响
在干旱胁迫条件下,植株水分严重亏缺,为防止水分进一步流失,植株通常会通过降低蒸腾强度来实现(图2)。对照组夹竹桃的蒸腾速率为(0.89±0.37)mmol H2O/m2·s,干旱组夹竹桃的蒸腾速率降为(0.35±0.08)mmol H2O/m2·s,但是与对照组的蒸腾速率变化差异极显著(P<0.01)。复水后,对照组夹竹桃的蒸腾速率为(1.29±0.25)mmol H2O/m2·s,干旱胁迫下夹竹桃的蒸腾速率为(1.31±0.39)mmol H2O/m2·s,两组植株的蒸腾速率都有升高,且组间差异不显著(P>0.05),这表明干旱组夹竹桃在复水得到充足水分后,其蒸腾速率恢复到正常水平。
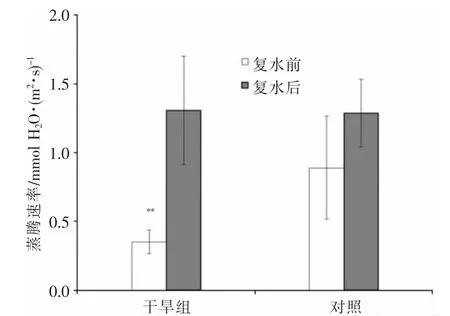
图2 复水前后夹竹桃叶片蒸腾速率
2.3 干旱胁迫对夹竹桃水分利用率(WUE)的影响
随着干旱情况的不同,植物对水分的利用率也会发生变化(图3)。干旱组的夹竹桃植株在经历11个周期的干旱胁迫后,植株叶片干卷,对水分的利用率出现负值,为(-11.42±4.81)g/kg,而同一时间对照组夹竹桃对水分的利用率为(3.05±0.54)g/kg,两组植株的水分利用率差异极大,达到了极显著水平(P<0.01);复水后,两组植株的水分利用率都得以提高,但是组间差异仍然显著(P<0.05),这表明在获取充足水分后干旱组植株的水分利用率还不能够达到对照组水平。
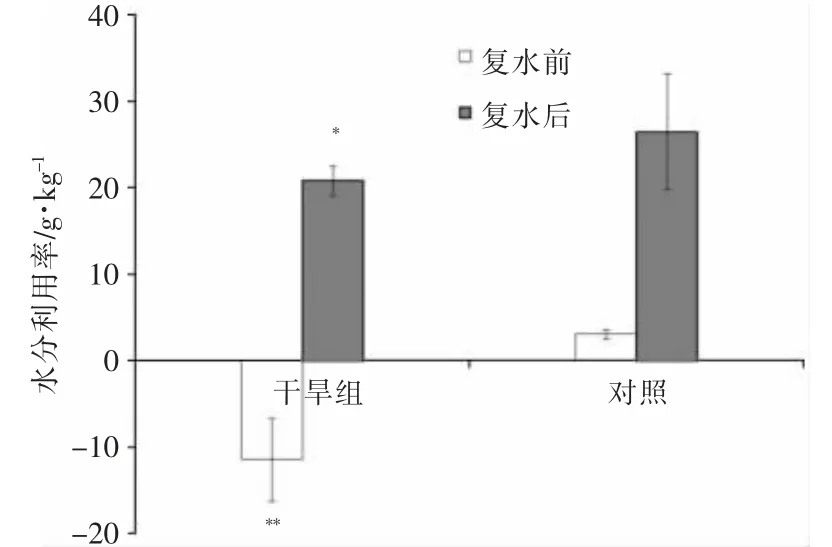
图3 复水前后夹竹桃叶片水分利用率
2.4 干旱胁迫对夹竹桃气孔导度(Gs)的影响
由图4可见,夹竹桃干旱组植株在11个周期的干旱胁迫后,叶片气孔导度为(0.02±0.01)mol/m2·s,而同一时间对照组夹竹桃叶片气孔导度为(0.13±0.06)mol/m2·s,相比对照组,干旱组植株的气孔导度有所降低,且变化差异极显著(P<0.01)。干旱组与对照组植株的气孔导度复水后,两组植株的气孔导度都有所提高,分别是干旱组为(0.10±0.04)mol/m2·s,对照组为(0.12±0.02)mol/m2·s,但组间差异不显著(P>0.05),这表明有充足水分供应后干旱组植株的气孔导度恢复的正常水平。

图4 复水前后夹竹桃叶片气孔导度
2.5 干旱胁迫对夹竹桃胞间CO2浓度(Ci)的影响
通常用胞间CO2浓度来分析植株光合速率变化是否由气孔因素导致。如图5所示,夹竹桃干旱组植株在11个周期的干旱胁迫后,叶片胞间CO2浓度为(331.44±74.44)μmol/mol,而同一时间点对照组夹竹桃叶片胞间CO2浓度为(240.22±57.44)μmol/mol。相比对照组,干旱组植株的胞间CO2浓度有所增加,但变化不显著(P>0.05)。复水后,对照组与干旱组夹竹桃植株的胞间CO2浓度都有所降低,分别是干旱组为(187.00±18.24)μmol/mol,对照组为(179.50±46.14)μmol/mol,变化不显著(P>0.05)。这表明获得充足水分后,干旱组植株的胞间CO2浓度可以达到对照组水平。

图5 复水前后夹竹桃叶片胞间CO2浓度
通过pearson相关分析发现,所检测的夹竹桃光合特性指标中,净光合速率、蒸腾速率、水分利用率、气孔导度等均与干旱胁迫呈极显著负相关(表1)。夹竹桃植株的水分利用率和净光合速率、蒸腾速率呈极显著相关。

表1 复水前干旱胁迫下夹竹桃光合指标间相关性分析
3 讨论
在本实验中,干旱胁迫开始时栽培有夹竹桃植株的土壤,其表层含水率为21.21%,第1周期对照组就降到10.83%,干旱组降为10.24%。剩余实验期间,对照组由于周期性浇水,其表层土壤含水率在7.24%~21.37%之间呈现规律性变化;干旱组的表层土壤含水率第2周期降为5.16%,此后除了第3周期的4.73%外,均保持在1.10%~2.61%。最后一个干旱周期结束时,干旱组表层土壤含水率为1.30%,处于极度缺水的状态[9]。
3.1 干旱胁迫对净光合速率的影响
经历了11个周期干旱胁迫的夹竹桃,其净光合速率为(-1.57±0.57)μmol CO2/m2·s,这与刘建伟等研究干旱胁迫下杨树(Pterocarya stenoptera)净光合作用的变化趋势相类似[11]。表明随着干旱胁迫的持续,干旱组中的夹竹桃植株呼吸作用已超过光合作用的产能,进入净呼吸消耗代谢。复水后,随着干旱胁迫的消除,夹竹桃叶片的净光合速率迅速增加转为正值,且与对照植株相差不大(约2.23 μmol CO2/m2·s)。说明在干旱胁迫消除后,夹竹桃植株的光合系统得以较快恢复。
3.2 干旱胁迫对蒸腾速率的影响
在植株经历轻度及中度干旱胁迫时,气孔因素是影响其光合作用的主要原因。在重度干旱胁迫下,主要影响植株光合作用的是非气孔因素[12],伴随着气孔关闭,植株的蒸腾作用也会降低,而下降程度会因植物种类及受干旱胁迫程度存在一定差异[13,14]。研究发现,在干旱胁迫下,植物会很快通过降低蒸腾作用以减少对于水分的消耗,但是其光合作用水平则不会立即改变[15]。
3.3 干旱胁迫对水分利用率的影响
净光合速率和蒸腾速率的比值即为水分利用率[16],因为气孔的开闭程度影响着叶片的蒸腾速率和光合速率,从而也必将影响叶片的水分利用效率。通常情况会在植株遭受水分胁迫后有不同程度的提高[17],以高效利用水资源。但是在本研究中,相对于复水前夹竹桃的水分利用率,干旱组在经历11个干旱周期后,其水分利用率显著降低,甚至降为负值,可能原因为植株遭受极端干旱的情况下,植株呼吸作用消耗有机质的量超过光合作用合成的量,导致净光合速率为负值,因此水分利用率也为负值。这与赵瑜琦等[18]在群众杨面临干旱胁迫时的反应趋势相类似。
3.4 干旱胁迫对气孔导度的影响
在经历干旱胁迫时,植物可以通过降低气孔导度来减少自身水分散失,以增强对干旱胁迫的耐受[19]。复水前干旱组夹竹桃植株的气孔导度显著低于对照组,复水后干旱组夹竹桃的气孔导度大幅提升,达到与对照组接近的水平,这说明虽然经历长时间的严重干旱,夹竹桃通过降低气孔导度以缓解水分的快速流失,一旦水分得到补充,夹竹桃气孔导度会迅速恢复到接近正常水平,体现了其耐旱性强,适应性好的特征。
3.5 干旱胁迫对胞间CO2浓度的影响
目前认为干旱胁迫导致植物光合作用降低的主要原因包括气孔限制和非气孔限制:前者是指由于气孔关闭,切断了CO2向叶绿体的运输通道,进而使叶肉细胞光合活性下降,同时蒸腾速率降低;后者是指在水分胁迫下,植株叶绿体的结构和功能遭受破坏,叶肉同化CO2的能力从而减弱[12]。胞间CO2浓度的变化趋势可以作为气孔因素影响光合速率变化的主要依据[20]。实验中,复水前干旱组夹竹桃植株的胞间CO2浓度高于对照组,复水后干旱组夹竹桃胞间CO2浓度大幅下降,这说明在干旱胁迫下夹竹桃光合速率降低的原因除了气孔因素外,还有植株体内光合色素和光合酶的参与。干旱胁迫对植物的影响是多方面的,不但会降低植株的净光合速率,还会影响植株的形态和生理指标[9]。夹竹桃具有较强的耐旱性,可能与其叶片的特殊结构有关:该植物的叶片具有发达的角质层,表皮由多层细胞组成;气孔下陷,栅栏组织层数较多,海绵组织和胞间间隙不发达[21]。当前对于夹竹桃抗旱特性的研究还大多局限在细胞内溶物、光合特性等生理生化指标上,对于其背后的分子机制涉及的较少。为了更加深入了解该植物的抗逆机制和适应严苛环境的机理,需要我们从基因组层面进一步挖掘夹竹桃的特殊基因,丰富抗逆相关基因资源,推进我国植物生物工程建设。
4 结论
干旱胁迫导致夹竹桃植株净光合速率、蒸腾速率、水分利用率、气孔导度等极显著降低,胞间CO2浓度有所增高。复水后,植株的前4个光合特性指标都有较大的提升,胞间CO2浓度降低。复水后以上指标与对照无显著差异,这表明该种植物具有较强的耐旱性,在遭受严重干旱胁迫下,其光合系统未受到严重损伤,仍可在复水后及时恢复。此研究从光合特征角度揭示了夹竹桃对于重度干旱胁迫的响应,进一步说明该种植物优秀的耐旱特质,可为海岛等干旱环境植被修复过程中植物的选用提供参考。
