松材线虫HAP基因家族鉴定及生物信息学分析1)
2022-10-08曹景鑫郝昕李洋李璐张月梁立巍杨显良马玲
曹景鑫 郝昕 李洋 李璐 张月 梁立巍杨显良马玲
(东北林业大学,哈尔滨,150040)(赤塔农业科技研究所,俄罗斯)(东北林业大学)
松材线虫(Bursaphelenchusxylophilus)作为世界范围针叶树病害松材线虫病的病原,是严重危险森林生态系统的病原物。松材线虫不仅对许多国家的林业造成巨大损失,而且对森林生态系统造成极大的破坏[1-2]。2021年松材线虫病在我国已传播扩散至19个省(自治区、直辖市)742个县级行政区,发生面积达1.72×106hm2,致死树木1.41×107株,造成经济和生态价值的巨大损失[3-4]。
松材线虫属于迁移型植物内寄生线虫,具有与其它植物寄生线虫不同的取食策略,并通过这种独特的取食策略实现在寄主体内繁殖与生存[5]。松材线虫在侵染寄主初期,通过口针释放多种效应因子到寄主体内,破坏寄主的防御体系,达到对寄主的入侵的目的[6]。效应因子(如β-1,4-内切葡聚糖酶基因[7]、果胶酸裂解酶[8]、磷酸酶等)与寄主间存在较为复杂的相互关系[9-10],并且在降解寄主细胞壁、调控寄主植物细胞周期和抵抗寄主防御反应等方面具有重要作用。
磷酸酶作为病原物侵入寄主后分泌的效应因子,能够改变寄主体内信号转导物质的磷酸化状态,增强病原致病性。如绿僵菌(Metarhiziumanisopliae)侵染寄主昆虫后,会分泌酸性磷酸酶来降解昆虫血液中的有机磷,产生无机磷为自身繁殖和生长提供营养[11-13];许多侵染动植物的革兰氏阴性菌也会通过向寄主体内分泌磷酸酶,干扰寄主免疫反应并增强自身致病性[14];秀丽线虫(Caenorhabditiselegans)分泌的酸性磷酸酶也与胚胎发育和抗逆态的形成密切相关[15-16];寄生在植物体内的线虫分泌的磷酸酶在侵染寄主过程中发挥着重要作用(如南方根结线虫(Meloidogyneincognita)侵染寄主植物后,会分泌磷酸酶到寄主体内发挥消化作用,从而促进线虫寄生[17];禾谷孢囊线虫(Heteroderaavenae)侵染寄主植物后会分泌酸性磷酸酶以促进对寄主的早期侵染[18])。
松材线虫在对寄主的侵染过程中,实践证明松材线虫寄生基因和效应因子发挥着重要作用[19-23],但是松材线虫分泌的蛋白磷酸酶功能和作用尚未见报道。因此,本文选取松材线虫酸性磷酸酶基因家族中成员较多的组氨酸酸性磷酸酶(HAP)基因家族开展研究。从生物信息学角度分析松材线虫HAP基因家族,利用松材线虫的基因组数据筛选松材线虫HAP基因家族成员,通过生物信息工具对其进行理化性质、信号肽、跨膜结构域、系统发育、基因定位、基因结构与保守基序分析,以及分析松材线虫接种黑松不同时期和被松树代谢产物β-蒎烯[24]胁迫后的转录组数据,获得松材线虫HAP基因家族成员在不用处理下的基因表达谱,为进一步揭示HAP基因家族在松材线虫侵染寄主过程中所发挥的功能提供理论基础。
1 材料与方法
1.1 松材线虫HAP基因家族成员鉴定及理化性质
通过NCBI(http://www.ncbi.nlm.nih.gov/)数据库获取松材线虫最新全基因组SAMEA7282713的相关数据。再通过NCBI蛋白数据库查询并下载秀丽线虫的HAP蛋白序列,利用该蛋白序列本地blast比对检索松材线虫蛋白数据库,获得松材线虫HAP候选蛋白序列。登陆Pfam数据库查询并下载HAP所具有的保守结构His_Phos_1(PF00300)和His_Phos_2(PF00328)的隐马尔可夫模型,利用HMMER3.0软件在松材线虫蛋白数据库中检索具有以上两个结构域的候选蛋白序列。在NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)和Pfam(http://pfam.xfam.org/)上鉴定两种方法所得候选蛋白序列是否具有His_Phos_1和His_Phos_2两种保守结构域,删除冗余序列,获得松材线虫HAP基因家族成员蛋白序列。利用在线软件Expasy(https://web.expasy.org/cgi-bin/protparam)[25],SignalP4.1(http://www.cbs.dtu.dk/services/SignalP/)和TMHMM2.0(http://www.cbs.dtu.dk/services/TMHMM/)分析松材线虫HAP基因家族成员的理化性质,信号肽和跨膜结构域。
1.2 松材线虫HAP基因家族系统发育分析
为进一步了解松材线虫HAP基因家族成员的进化关系,使用MEGA11.0软件对36个秀丽线虫和34个松材线虫HAP蛋白序列进行CLUSTAL W多序列比对,依据多序列比对结果使用MEGA软件计算最适模型(LG+G),构建最大似然进化树,Bootstrap检验1 000次。随后通过iTOL在线网站(https://itol.embl.de/)对系统进化树美化。
1.3 松材线虫HAP基因家族染色体定位
根据松材线虫基因结构注释文件,使用TBtools软件[26]的Gene Density Profile功能提取线虫基因密度信息。再依据线虫基因结构注释文件和基因密度信息,使用TBtools的Gene Location Visualize from GTF/GFF功能绘制BxHAP基因在松材线虫染色体上的分布情况。
1.4 松材线虫HAP基因家族基因结构及保守基序
通过松材线虫基因结构注释文件提取松材线虫HAP基因的外显子/内含子结构,应用在线工具GSDS2.0(http://gsds.gao-lab.org/)对基因结构进行可视化。使用模因(MEME)(https://meme-suite.org/meme/tools/meme/)分析松材线虫HAP基因家族成员的保守基序分布,基序参数值设为19,其它参数默认。
1.5 松材线虫HAP基因家族在不同处理下的基因表达谱
根据松材线虫接种3年生黑松幼苗的转录组数据(PRJNA397001)和被不同浓度β-蒎烯胁迫后的转录组数据(PRJNA640733),获得34个松材线虫HAP基因在不同处理下的表达值(FPKMs),使用R语言pheatmap软件包对所得数据进行表达量标准化处理及热图绘制。
1.6 松材线虫被不同松树代谢产物胁迫后体内BxHAP基因表达定量分析
为验证松材线虫HAP基因家族成员与松树代谢产物互作的能力,选用东北林业大学森林保护学科生物农药与分子生物学实验室的松材线虫材料,使用质量浓度为0、200、400、600、800、1 000 mg/L的松树代谢产物α-蒎烯(麦克林,上海)和β-蒎烯(麦克林,上海)处理50 条混合虫龄的松材线虫,每处理重复三组。统计代谢产物处理松材线虫24 h后线虫存活率变化情况,使用SPSS 26.0软件计算致死中间浓度(LC50)。再使用LC50的α-蒎烯和β-蒎烯处理松材线虫,提取松材线虫总RNA。Primer Premier 6.0设计4个上调基因的荧光定量引物(见表1)。荧光定量使用2×SYBR Green qPCR Master Mix试剂盒,反应体系为94 ℃预变性30 s;94 ℃变性15 s,58 ℃退火30 s,72 ℃延伸60 s,40个循环;95 ℃变性1 min,55 ℃延伸30 s,95 ℃变性30 s。反应所得基因相对表达量采用2-ΔΔCt法计算,每个基因重复定量3次,使用GraphPad Prism 9软件绘制松材线虫基因表达量变化图。

表1 实时荧光定量PCR引物
2 结果与分析
2.1 松材线虫HAP基因家族成员鉴定及理化性质
由表2可知,经过两次检索比对,以及NCBI-CDD和Pfam验证保守结构域后,从松材线虫基因组中鉴定出34个HAP基因家族成员,依据成员的保守结构域与染色体位置分布信息,将其命名为BxHAP1~BxHAP34。BxHAP基因所编码蛋白大小的跨度较大,其中最小的是BxHAP6(212个氨基酸),而最大的是BxHAP11(1447个氨基酸)。BxHAP蛋白的相对分子量介于24 322.84~165 183.69,相对分子量最低的是BxHAP7(24 322.84),最高的仍是BxHAP11(165 183.69)。BxHAP蛋白的理论等电点为5.12~9.91,其中等电点最低的是BxHAP12(5.12),最高的为BxHAP1(9.91),大部分BxHAP蛋白富含酸性氨基酸,主要在酸性亚细胞环境发挥功能。因此,说明松材线虫HAP基因家族的不同成员间理化性质差异较大,各成员在生物体中可能具有不同的功能。信号肽和跨膜结构域预测分析表明,具有信号肽和跨膜结构域的有BxHAP12、BxHAP13、BxHAP14、BxHAP15、BxHAP20、BxHAP22、BxHAP23、BxHAP26、BxHAP27、BxHAP29、BxHAP31和BxHAP33等12个成员,这些成员符合典型效应因子的条件[27],而其它成员可能属于非典型效应因子[28-29]。
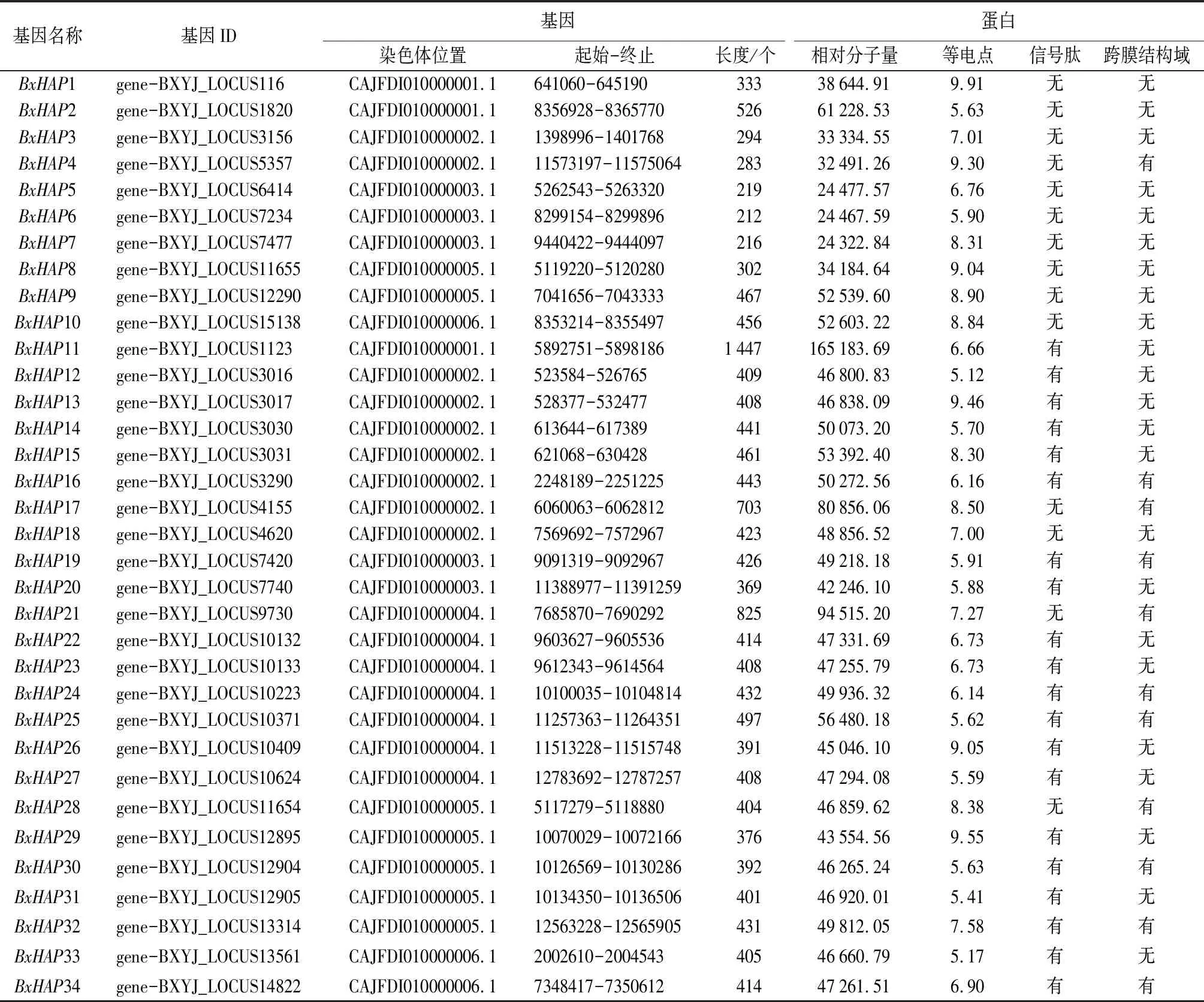
表2 松材线虫HAP基因家族成员理化性质
2.2 松材线虫HAP基因家族系统发育分析
由图1可知,通过松材线虫和秀丽线虫HAP蛋白序列构建的系统进化树,明确了松材线虫HAP基因家族成员间的进化关系。根据进化树聚类分析,将松材线虫HAP和秀丽线虫HAP基因家族共70个成员划分为6个亚家族:第Ⅰ、Ⅳ亚族为具有His_Phos_1保守结构域的亚族,因两者间亲缘关系较远,故分成两个亚族。其中松材线虫HAP基因家族第Ⅰ亚族有BxHAP1、BxHAP4和BxHAP9等3个成员,第Ⅳ亚族有BxHAP3、BxHAP5、BxHAP6、BxHAP7和BxHAP10等5个成员;秀丽线虫HAP基因家族第Ⅰ亚族有CeHAP1和CeHAP2两个成员,第Ⅳ亚族有CeHAP5、CeHAP6、CeHAP9和CeHAP10等4个成员。第Ⅱ亚族成员含有His_Phos_2、PPIP5K2_N和RimK等3个保守结构域,其中BxHAP11属于松材线虫HAP基因家族第Ⅱ亚族;CeHAP31和CeHAP32属于秀丽线虫HAP基因家族第Ⅱ亚族。第Ⅲ亚族成员含有6PF2K和His_Phos_1保守结构域,包含一个松材线虫HAP基因家族成员BxHAP2和三个秀丽线虫HAP基因家族成员CeHAP4、CeHAP7和CeHAP8。第Ⅴ亚族成员含有His_Phos_1或His_Phos_2保守结构域,其中BxHAP8、BxHAP17属于松材线虫HAP基因家族第Ⅴ亚族;CeHAP3、CeHAP11、CeHAP20、CeHAP34、CeHAP35和CeHAP36等属于秀丽线虫HAP基因家族第Ⅴ亚族。第Ⅵ亚族是最大的亚族,含有His_Phos_2保守结构域,其中松材线虫HAP基因家族第Ⅵ亚族有BxHAP12、BxHAP13、BxHAP14、BxHAP15、BxHAP16、BxHAP18、BxHAP19、BxHAP20、BxHAP21、BxHAP22、BxHAP23、BxHAP24、BxHAP25、BxHAP26、BxHAP27、BxHAP28、BxHAP29、BxHAP30、BxHAP31、BxHAP32、BxHAP33、BxHAP34等22个成员;秀丽线虫HAP基因家族第Ⅵ亚族有CeHAP12、CeHAP13、CeHAP14、CeHAP15、CeHAP16、CeHAP17、CeHAP18、CeHAP19、CeHAP21、CeHAP22、CeHAP23、CeHAP24、CeHAP25、CeHAP26、CeHAP27、CeHAP28、CeHAP29、CeHAP30、CeHAP33等18个成员。
2.3 松材线虫HAP基因家族染色体定位
由图2可知,松材线虫HAP基因染色体定位图阐述了BxHAP基因在染色体上的分布情况。根据BxHAP在松材线虫染色体上的分布信息,34个BxHAP基因均匀分布在前6条染色体上。BxHAP1、BxHAP2和BxHAP11等3个BxHAP基因均匀分布第1条染色体;BxHAP3、BxHAP4、BxHAP12、BxHAP13、BxHAP14、BxHAP15、BxHAP16、BxHAP17和BxHAP18等9个BxHAP基因分布在第2条染色体上,其中BxHAP12~BxHAP15集中分布在染色体的顶端。BxHAP5、BxHAP6、BxHAP7、BxHAP19和BxHAP20等5个BxHAP基因集中分布在第3条染色体的中下部;BxHAP21、BxHAP22、BxHAP23、BxHAP24、BxHAP25、BxHAP26、BxHAP27等7个BxHAP基因集中分布在第4条染色体的下部;BxHAP8、BxHAP9、BxHAP28、BxHAP29、BxHAP30、BxHAP31和BxHAP32主要分布在第5条染色体的中下部,其中BxHAP29、BxHAP30和BxHAP31集中在染色体下部;BxHAP10、BxHAP3和BxHAP34等3个BxHAP基因均匀分布在第6条染色体上。表明34个BxHAP基因均匀分布在松材线虫的6条染色体上,属于松材线虫的关键基因家族。

图1 松材线虫与秀丽线虫HAP基因家族系统进化树
2.4 松材线虫HAP基因家族的基因结构及保守基序
由图3可知,根据松材线虫HAP基因家族基因结构,明确了松材线虫HAP基因家族的内含子和外显子的组成情况。34个BxHAP基因中,BxHAP6、BxHAP7、BxHAP20含有的外显子数量最少,每基因仅含有3个外显子,BxHAP11含有最多,每基因含有15个外显子。BxHAP基因中拥有5个外显子的最多,共有8个基因(BxHAP1、BxHAP3、BxHAP4、BxHAP8、BxHAP19、BxHAP25、BxHAP33和BxHAP34);拥有7个外显子的基因有BxHAP13、BxHAP22、BxHAP23、BxHAP26和BxHAP31;拥有6个外显子的基因有BxHAP9、BxHAP12、BxHAP24和BxHAP29;拥有9个外显子的基因由BxHAP2、BxHAP10、BxHAP14和BxHAP28。然后为拥有8个外显子的基因有BxHAP18、BxHAP27与BxHAP30;拥有10个外显子的基因有BxHAP15、BxHAP16和BxHAP3;拥有4、11、13个外显子的基因分别是BxHAP5、BxHAP17和BxHAP21。表明松材线虫HAP基因家族成员的功能在家族进化过程中具有较大分化。

图2 松材线虫HAP基因染色体定位图

图3 松材线虫HAP基因家族基因结构
由图4可知,34个松材线虫HAP基因家族成员共含有19种保守基序,分别命名为Motif1~Motif19。34个成员均具有Motif2,以及除BxHAP10外都具有Motif3。BxHAP1~BxHAP10中,除BxHAP4外,仅具有Motif13,而没有Motif1;BxHAP11~BxHAP34均仅具有Motif1,而没有Motif13。其余Motif随机分布于各成员中,且相同Motif在不同成员上排列顺序近乎一致。利用NCBI-CDD和Pfam软件对19种Motif进一步分析,发现不同成员的Motif2和Motif3均定位于His_Phos_1或His_Phos_2保守结构域中,Motif1定位于His_Phos_2保守结构域中,Motif13定位于His_Phos_1保守结构域中。结果进一步证实34个基因成员均属于松材线虫HAP基因家族,说明系统进化树对松材线虫HAP基因家族亚家族的划分准确。

图4 松材线虫HAP基因家族保守基序结构图
2.5 松材线虫HAP基因家族在不同处理下的基因表达谱
由图5可知。有4个BxHAP基因在接种黑松不同时期和被β-蒎烯胁迫后的松材线虫体内均表现为上调表达,分别为BxHAP1、BxHAP5、BxHAP17和BxHAP20;有2个基因在接种黑松不同时期和被β-蒎烯胁迫后的松材线虫体内均表现为下调表达,分别为BxHAP2和BxHAP21;有且仅有BxHAP8在接种黑松不同时期后的松材线虫体内表达上调,却在被β-蒎烯胁迫后的松材线虫体内表达下调;有6个BxHAP基因在接种黑松不同时期后的松材线虫体内表达下调,却在被β-蒎烯胁迫后的松材线虫体内表达上调,分别为BxHAP3、BxHAP18、BxHAP27、BxHAP30、BxHAP33和BxHAP34;其余21个BxHAP基因在两种不同处理条件下,各阶段表达趋势各异。因此,大多数松材线虫HAP基因家族成员在不同处理下表达趋势均发生明显改变,证明BxHAP基因在松材线虫侵染松树过程中发挥重要功能,同时证明该家族成员具有响应松树代谢产物的能力,尤其是两种处理条件下基因表达量均表现为上调表达的BxHAP1、BxHAP5、BxHAP17和BxHAP20等4个基因。
2.6 松材线虫被不同松树代谢产物胁迫后体内BxHAP基因表达定量分析
为进一步确认4个基因与松树代谢产物之间的互作方式,选取不同质量浓度松树代谢产物α-蒎烯和β-蒎烯胁迫松材线虫。根据松材线虫24 h后的存活率变化情况,统计得出α-蒎烯和β-蒎烯对松材线虫的24 h致死的中间浓度(LC50)分别为854.284、797.919 mg/L。因此,β-蒎烯比α-蒎烯对松材线虫有更高的杀线活性。
由表3可知,β-蒎烯处理松材线虫后,线虫体内4个BxHAP基因表达量均表现为显著上调表达(p<0.05);而α-蒎烯处理松材线虫后,线虫体内4个BxHAP基因出现不同的变化趋势,BxHAP1表现为不显著下调表达,BxHAP5表现为显著上调表达(p<0.01),BxHAP17表现为显著下调表达(p<0.01),BxHAP20表现为不显著上调表达。因此,BxHAP基因确实具有响应松树代谢产物的能力,且对不同松树代谢产物具有不同的功能,其中,BxHAP5基因表达量在两种处理条件下,松材线虫体内均表现为显著上调表达(p<0.01),表明BxHAP5基因在松材线虫抵抗松树代谢产物的过程中发挥重要作用。

图5 松材线虫HAP基因在不同处理下的表达谱
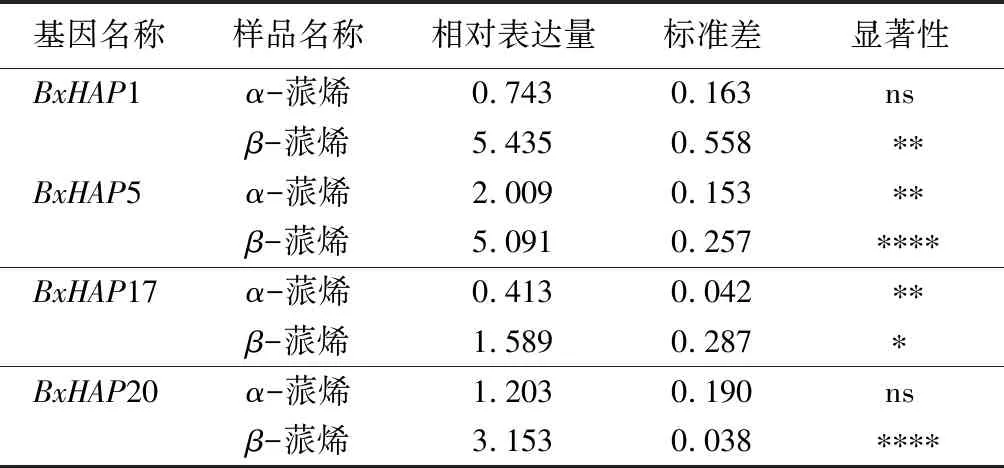
表3 松材线虫被不同松树代谢产物胁迫后体内BxHAP基因表达情况
3 结论与讨论
松材线虫病自1982年传入我国,对我国林业生态系统造成了极大的危害。松材线虫与寄主间的互作关系因其所具有的隐蔽性特征而极为复杂[30-32]。因此,分析松材线虫入侵寄主松树并定殖的分子机制对于防治松材线虫具有极为重要的意义。根结线虫(Meloidogyne)[33]和孢囊线虫[18]在侵染植物过程中会分泌磷酸酶破坏寄主防御体系,从而进入寄主体内建立寄生关系。
本文对松材线虫HAP基因家族进行了鉴定和功能分析,共鉴定出34个松材线虫HAP基因家族成员。松材线虫HAP基因家族成员的氨基酸长度在212~1 447个氨基酸残基间分布,相对分子量在24 322.84~165 183.69,多数成员为酸性蛋白;12个松材线虫HAP基因家族成员为典型效应因子,其余成员可能为非典型效应因子;34个松材线虫HAP基因家族成员分属于6个不同的亚家族;松材线虫HAP基因家族成员均匀分布在6条染色体上;各亚家族成员之间差异较大,但同亚家族的成员间具有相似的基因结构及保守基序,说明在线虫进化过程中BxHAP基因发生了保留和丢失[34]。BxHAP基因在松材线虫接种黑松不同时期和被松树代谢产物β-蒎烯胁迫后的基因表达谱表明,松材线虫HAP基因家族成员在松材线虫侵染寄主过程中发挥重要功能,同时证明该家族成员能够响应松树代谢产物,尤其是被两种处理后基因表达量均上调表达的BxHAP1、BxHAP5、BxHAP17和BxHAP20等4个基因。通过松材线虫被不同松树代谢产物胁迫后体内BxHAP基因表达定量分析,验证了转录组数据分析的准确性,发现BxHAP基因针对不同松树代谢产物时具有不同的功能,尤其是BxHAP5在两种处理条件下均显著上调表达,表明其是松材线虫抵抗松树代谢产物过程中的关键基因。综上所述,本研究证实了松材线虫HAP基因家族成员在松材线虫侵染松树过程中确实发挥着重要功能,能够与松树代谢产物相互作用,且针对不同的松树代谢产物的作用方式不尽相同。同时发现并鉴定了松材线虫抵抗松树代谢产物过程中的关键基因BxHAP5,但其对松树代谢产物的具体作用机理仍需要进一步探索与发现。
