藻株X6发酵动力学模型及肌醇对其生理影响研究
2022-10-08谢丽娜田璐毅桂媛媛陶俊恒李汉广
谢丽娜,田璐毅,桂媛媛,陶俊恒,李汉广
(江西农业大学 生物科学与工程学院/应用微生物研究所,江西 南昌 330045)
【研究意义】随着全球经济的快速发展,人类对能源的需求日益增加,由于传统化石燃料储量的逐年减少,寻找可再生、清洁环保型能源十分迫切[1]。生物柴油是目前较为成功的新型清洁能源,其生产原料主要来源于油菜、大豆等植物以及各种动物油脂[2]。与这些动植物原料相比,微藻具有许多较为明显的优势:光合效率高、生长周期短、油脂含量高及一年四季均可培养等优点[3-4]。因此加强微藻生物柴油的研究对加快发展新型清洁环保能源,减少人类对化石能源的依赖具有重要的现实意义。【前人研究进展】发酵动力学可以模拟发酵过程中的动态变化,可为提高相应微生物的生长性能提供一定的理论基础[5]。宋增光等[6]通过构建动力学模型来预测紫红曲菌发酵薏米产α-生育酚过程,其结果与实际测定值接近,在通过该动力学模型指导下进行代谢调控发酵可以缩短生产周期、促进产物α-生育酚的合成。另一方面,为提高微藻生物柴油的竞争性,提高其油脂含量是现实可行的一种方式,传统发酵过程中的代谢调控手段常应用于微藻的生产。肌醇广泛存在于各种动植物和微生物体内,参与生物素的运输以及细胞壁生物合成等过程,被认为是微藻等光合生物的生长促进剂[7]。Erbil 等[8]研究发现当向培养体系中添加500 mg/L 的肌醇时,海洋紫球藻的生物量相较于对照组提高了1.28倍。【本研究切入点】本实验室前期分离筛选获得了一株生长性能较为优良藻株X6,但其生长特性及如何提高其油脂含量并未进行深入研究,因此本试验拟通过构建藻株X6在不同培养条件下的动力学模型以及探究外源调控(添加肌醇)提高藻株油脂产量的内在生理机制,以期进一步提高该藻株的应用性。【拟解决的关键问题】通过Logistic、Luedeking-Piret 及Luedeking-Piret 修正方程构建藻株X6不同培养模式下的动力学模型,确定藻株X6最佳培养模式,并在此基础上探究外源添加肌醇对藻株X6生长及生理影响,以期通过本论文的开展为提高其它类似产油性能方面提供一定的理论及试验参考。
1 材料和方法
1.1 试验材料
本试验所用藻株X6[小球藻,Chlorellasp.(GenBank:MK951683.1)]是本课题组保藏藻种;肌醇购自索莱宝公司;培养基使用BG-11培养基,培养基成分参考胡文军等[9]。
1.2 试验设计
1.2.1 微藻培养 自养培养、异养培养和混养培养的培养方式参照黎秋玲等[10]建立的方法。
1.2.2 藻株X6动力学模型构建 葡萄糖底物消耗速率可以采用Luedeking-Piret修正模型[11],微藻细胞生长可采用Logistic 模型[12],产物合成采用Luedeking-Piret 方程[10]。对藻株X6生长过程中的生物量、油脂含量以及底物消耗等的数据进行分析,描述其生长过程中细胞生长、油脂合成及基质消耗随时间变化之间的关系。
1.2.3 不同浓度肌醇对藻株X6生长、油脂产量及生理影响 将藻液通过改良的BG-11培养基(外源添加15 g/L 葡萄糖的BG-11 培养基)培养至稳定期,以10%的接种量接种至已灭菌的改良BG-11 培养基中,添加已过滤除菌的肌醇,使得培养基中肌醇的最终浓度为0.5,1.0,1.5,2.5,5.0 mg/L,每组试验设置3 个重复,于120 r/min、27 ℃条件下摇床中避光培养6 d。
1.3 测定项目与方法
1.3.1 油脂含量的测定 采用胡文军等[9]建立的氯仿-甲醇提取法提取微藻细胞中的油脂,计算公式如下:

1.3.2 蒽酮比色法测定总糖含量(total sugar content,TSC)可溶性总糖含量采用蒽酮-硫酸法测定[13]。
1.3.3 葡萄糖含量的测定 培养基中葡萄糖含量采用二硝基酸鉴定法(DNS)测定[14]。
1.3.4 可溶性蛋白质含量的测定 蛋白质含量的测定采用考马斯亮蓝法[15]。
1.3.5 丙二醛(malondialdehyde,MDA)含量的测定[16]取藻泥0.5 g 加入5 mL 10%三氯乙酸及少量的石英砂,冰浴研磨成匀浆,4 000 r/min 离心10 min。向试管中加入2 mL 上清液和2 mL 0.6%硫代巴比妥酸溶液,混合均匀后沸水浴15 min,用分光光度计测其在450,532,600 nm 处的OD 值。计算公式如下:

式中Vt是提取液的总体积(mL),Vs是加入到反应体系的体积(mL),W是鲜重(g)。

式中V是藻液总体积。
1.4 数据统计及分析
采用Excel 2019进行数据整理,用Origin 9.0作图以及DPS 7.05进行差异显著性分析(P<0.05)。
2 结果与分析
2.1 不同培养条件下藻株X6生长动力学曲线
为获得藻株X6最佳培养方式及其对应的生长动力学曲线,对藻株X6进行自养、异养和混养培养,分析藻株X6生长过程中生物量、底物消耗和油脂产量随时间变化的关系,其结果如图1所示。由图1可知,在整个试验周期内,自养培养模式下藻株X6的生物量及油脂含量变化幅度小,生长缓慢,油脂含量没有明显增加。而异养和混养培养模式下,0~24 h,微藻处于延迟期,生物量和油脂含量没有显著增加,底物消耗较少;24~96 h,微藻处于对数生长期,生物量和油脂含量急速增加;在96 h 后,微藻进入平稳期,生物量变化不大,油脂含量继续增加,但增幅较对数生长期平缓,此时葡萄糖含量持续降低;当培养时间为144 h,混养培养条件下藻株X6的油脂含量达到最大值0.85 g/L;而对于异养培养而言,其油脂含量最大值出现在156 h 时,具体为0.82 g/L;混养培养和异养培养的最大生物量分别出现在168 h 和156 h,此刻它们的最大值分别为14.97 g/L和11.01 g/L。

图1 不同培养模式下藻株X6的生长特性(a)、油脂合成(b)和葡萄糖消耗(c)Fig.1 Growth(a),lipid content(b),glucose consumption(c)of microalga X6 for three culture modes
在整个微藻培养过程中,混养培养下藻株X6的生物量和油脂含量始终高于异养培养,且油脂合成与微藻生长和底物消耗趋势一致,表明藻株X6的生长与油脂合成的关系可能属于生长偶联型或部分生长偶联型模式[10],而自养培养条件下的生长特性在三者当中最差,因此后续仅探讨混养与异养模型的构建。
2.2 发酵动力学模型拟合求解与检验
将图1 的数据应用于Logistic、Leudeking-Piret 及Luedeking-Piret 修正模型,采用Origin 9.0 软件进行非线性拟合,结果如表1所示。由表1可知,不同培养条件下的最大比生长速率(μm)相近,混养培养条件下的μm略大于异养培养,说明混养培养更适应于X6生物量的积累。对不同培养条件下X6生长情况、产物合成以及底物消耗进行拟合时,其拟合相关系数R2的数值均在0.9 以上,表明拟合方程能够较好的描述X6不同培养条件下的生长情况。不同培养条件下的产物生成相关系数α和非产物生成相关系数β均不为零,且α远大于β。在产物合成过程中,异养培养条件下的最大油脂合成速率QPmax大于混养培养,0~84 h期间,异养培养条件下的油脂含量始终高于混养培养;而84~144 h混养培养的油脂含量更高。同时不同培养条件下,混养培养的底物最大比消耗速率QSmax高于异养培养,这一结果与图1不同培养条件的生长特性相符。
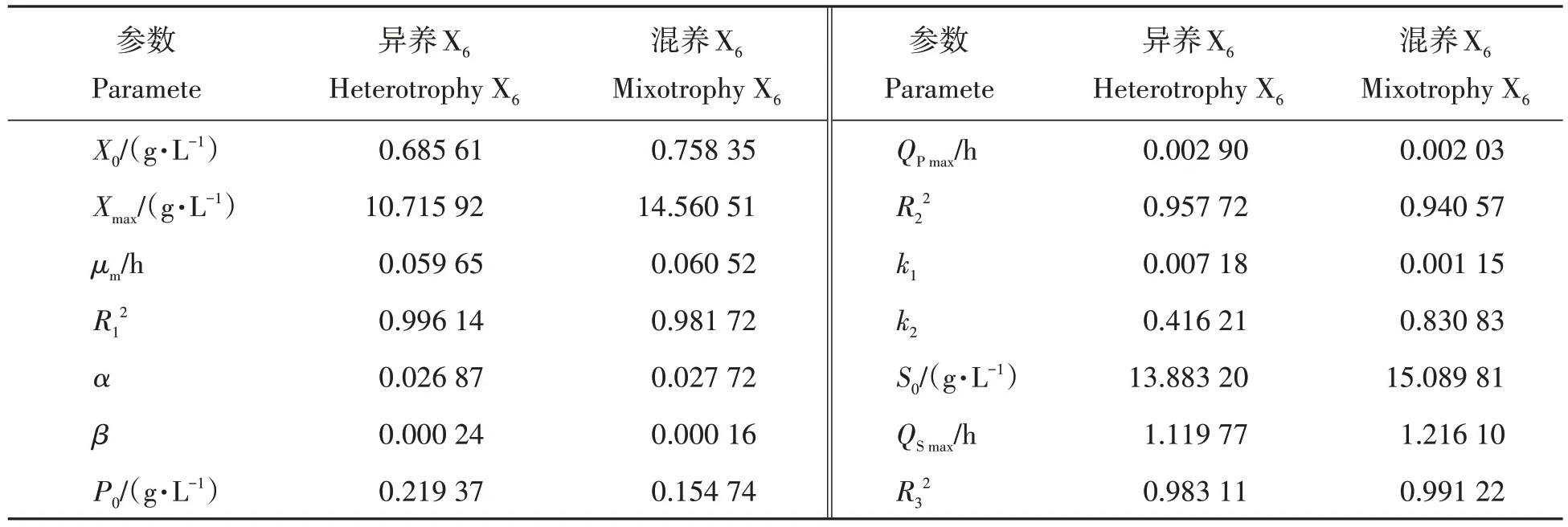
表1 动力学参数模拟值Tab.1 Simulation values of kinetic parameters
2.3 细胞生长模型
细胞生长动力学用Logistic方程描述:

式(4)中,dX/dt为藻细胞生长速度[g/(L·h)],μm为微藻最大比生长速率(h-1),X为微藻生物量(g/L),Xm为微藻的最大生物量(g/L),t为发酵时间(h)。
对Logistic方程积分,得:

用Logistic 方程对试验数据进行非线性拟合,结果见图2。藻株X6异养培养的拟合相关系数R2=0.996 14,混养培养的R2=0.981 72,说明该方程的实测值与预测值拟合良好,可以以此作为描述X6生长的模型。藻株X6生物量与发酵时间函数为:
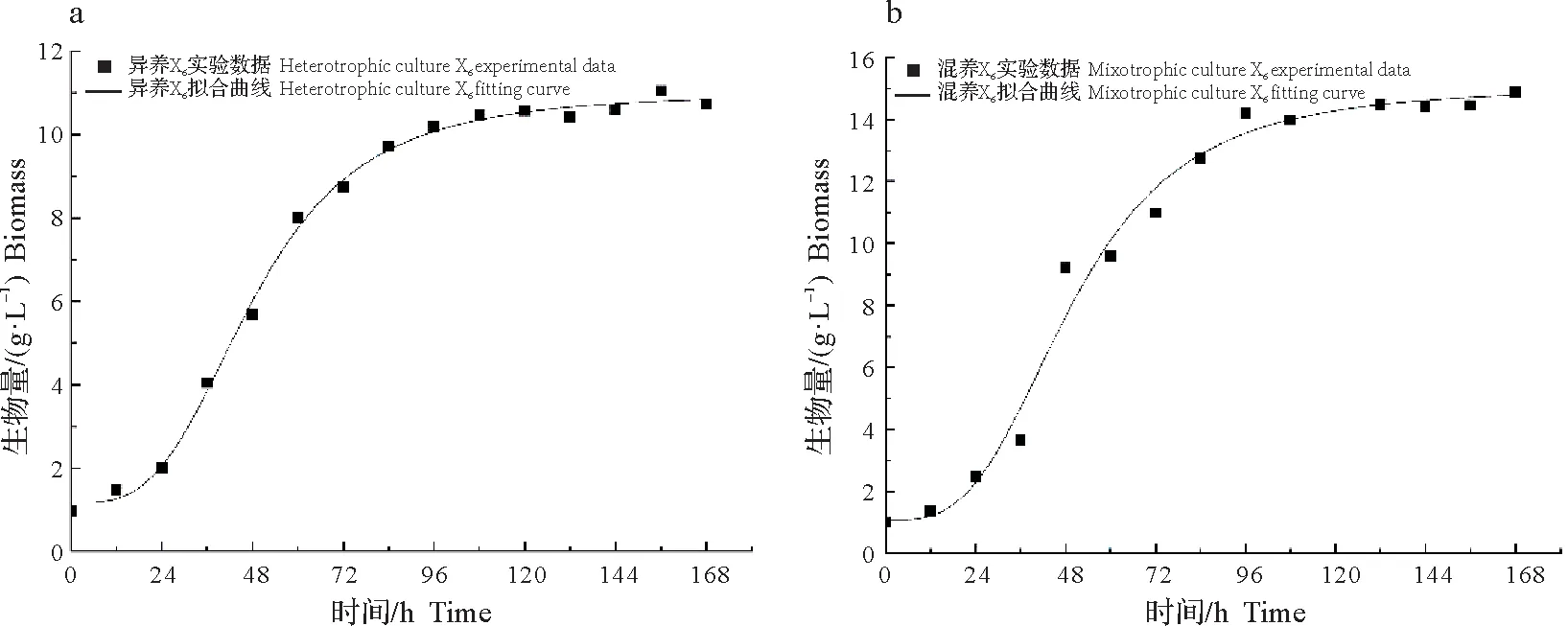
图2 不同培养条件下藻株X6细胞干重实验值与模型拟合结果的比较Fig.2 Comparison of the experimental cell dry weight of microalga X6with calculated values under different culture conditions
异养X6:X=7.345e0.05965t/(10.03+0.685 6e0.05965t)
混养X6:X=11.04e0.06052t/(13.80+0.758 3e0.06052t)
2.4 油脂合成动力学模型
微生物生长与产物合成的关系可分为生长偶联型(α≠0,β=0)、部分生长偶联型(α≠0,β≠0)和非生长偶联型(α=0,β≠0)。油脂是微藻细胞生长过程中的重要产物,根据Luedeking-Piret方程可将油脂合成动力学描述为:

式(6)中,P为油脂含量(g/L)。
从图1 可知,油脂的合成与细胞生长密切相关,当细胞进入对数生长期,油脂含量也迅速提高;当微藻细胞进入平稳期,油脂含量也逐渐趋于平稳。利用公式(4)、(5)和(6)建立油脂合成的数学模型,经换算得:

根据表1的数据代入可得:
异养X6:P=0.098 64+[0.197 41e0.05965t/(10.030 31+0.685 61e0.05965t)]+0.043 11 ln(10.030 31+0.685 61 e0.5965t)
混养X6:P=0.030 63+[0.306 08e0.06052t/(13.802 16+0.758 35e0.06052t)]+0.038 49 ln(13.802 16+0.758 35e0.5965t)
藻株X6的油脂产量拟合曲线如图3 所示,异养培养的拟合相关系数R2=0.957 72,混养培养的R2=0.940 57,结果表明实际数据与预测数据拟合良好,该方程可以较好的描述X6在不同培养条件下的产物合成情况。
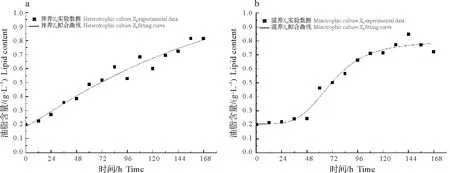
图3 不同培养条件下藻株X6油脂产量实验值与模型拟合结果的比较Fig.3 Comparison of the experimental lipid content of microalga X6 with calculated values under different culture conditions
2.5 基质消耗动力学模型
基质消耗速率可以采用Luedeking-Piret修正模型表示:

对方程进行换算可得:
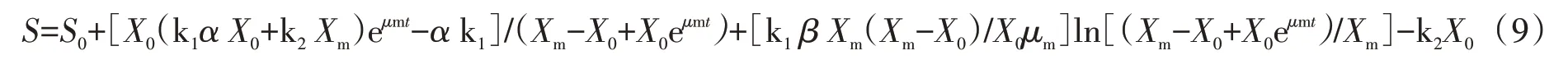
式(9)中S为葡萄糖浓度(g/L),S0为葡萄糖初始浓度(g/L),k1为微藻生长消耗系数,k2为产物消耗系数。
根据表1的数据代入可得:
异养X6:S=0.004 53ln(10.030 31+0.685 61e0.05965t)+(3.057 96e0.05965t-0.000 19)/(10.030 31+0.685 61e0.05965t)+13.587 10
混养X6:S=0.000 81ln(13.802 16+0.758 35e0.05965t)+(9.174 01e0.05965t-0.000 03)/(13.802 16+0.758 35e0.05965t)+14.457 59
藻株X6的底物消耗拟合曲线如图4 所示,异养培养的拟合相关系数R2=0.983 11,混养培养的R2=0.991 22,结果表明实际数据与预测数据拟合良好,可以以此方程描述X6在不同培养模式下的底物消耗情况。

图4 不同培养条件下藻株X6总糖含量的实验值与模型拟合结果的比较Fig.4 Comparison of the experimental total sugar content of microalga X6with calculated values under different culture conditions
结合动力学拟合曲线与试验数据可知,无论是异养培养还是混养培养,藻株X6的代谢变化趋势基本保持一致,混养培养结合了自养培养与异养培养的优点,其生物量显著高于异养和自养,且在混养条件下其油脂含量最高,更有利于微藻脂质的合成。
2.6 肌醇对藻株X6生长及油脂产量的影响
为探究不同浓度肌醇对藻株X6生长及油脂产量的影响,本试验将0.5,1.0,1.5,2.5,5.0 mg/L 的肌醇添加到改良的BG-11液体培养基(外加葡萄糖)中进行混养培养,发酵6 d后对藻株X6的生物量及油脂含量进行测定,结果如图5 所示。由图5 可知,X6的生物量随着肌醇浓度的增加而逐渐增长,并在5.0 mg/L时达到最大值12.18 g/L,相对于对照组提高了9.57%;同时,油脂含量也伴随肌醇浓度的增加而提高,在2.5 mg/L 时达到最大值1.70 g/L(此时油脂含量占藻细胞干重14.09%),相较于对照组提高了10.22%,虽然此时生物量与5.0 mg/L 添加条件下相比略有下降,但其油脂产率在肌醇浓度为2.5 mg/L 时达到最大值,故选择此浓度为最佳添加量。

图5 不同浓度的肌醇对藻株X6生物量及油脂产量的影响Fig.5 Effect of different concentrations of inositol on biomass and lipid production of microalga X6
2.7 肌醇对藻株X6生理生化性质的影响
2.7.1 肌醇对藻株X6胞内可溶性总糖含量的影响 肌醇对藻株X6胞内可溶性总糖含量的影响如图6所示。从图6 可知,可溶性总糖含量随着肌醇浓度的增加出现先增加后又降低的趋势,并在肌醇浓度为2.5 mg/L 时达到最大值140.11 mg/L,相对于对照组提高了87.54%,且外源添加肌醇的处理组胞内可溶性总糖含量始终高于对照组。

图6 不同浓度的肌醇对藻株X6胞内可溶性总糖含量的影响Fig.6 Effect of different concentrations of inositol on the intracellular total soluble sugar content of microalga X6
2.7.2 肌醇对胞内可溶性蛋白质含量的影响 可溶性蛋白质含量是微藻细胞一个重要的生理生化指标,它既可以反映细胞内部代谢的活跃程度,又能在渗透调节中起到重要作用,并且与生物体的生长发育、抗逆性等密切相关,因此其含量的变化可直接反应藻体内代谢情况[18]。由图7 可知,可溶性蛋白质的含量随着肌醇浓度的增加而逐渐降低,当肌醇浓度为2.5 mg/L 时,可溶性蛋白质含量最低,为0.58 mg/mL,且蛋白质含量始终低于对照组,以上结果说明,肌醇会降低藻株X6的胞内可溶性蛋白质含量。

图7 不同浓度的肌醇对藻株X6可溶性蛋白质含量的影响Fig.7 Effect of different concentrations of inositol on soluble protein content of microalga X6
2.7.3 肌醇对胞内丙二醛含量的影响 当微藻细胞受到外界不利因素胁迫时,机体会产生过量的活性氧(ROS),引起或加剧膜脂质过氧化作用,而丙二醛(MDA)是脂质过氧化反应的最终分解产物,因此丙二醛通常用于表示生物体在受到外界刺激后细胞的氧化损伤程度,丙二醛含量越高说明膜受损伤程度越大[19]。由图8 可知,随着肌醇浓度的增加,丙二醛的含量呈现先降低后又升高的趋势,在肌醇浓度为2.5 mg/L 时,丙二醛的含量达到最小值0.16 mmol/mL,与对照组相比较降低了64.85%,另一方面,肌醇的添加使得丙二醛的含量始终低于对照组。以上结果表明,外源肌醇的添加能够抑制微藻丙二醛的生成,降低微藻细胞膜系统受损伤的程度,对细胞膜有一定的保护作用。

图8 不同浓度的肌醇对藻株X6丙二醛含量的影响Fig.8 Effect of different concentrations of inositol on malondialdehyde content of microalga X6
2.7.4 肌醇对胞内超氧阴离子自由基含量的影响 超氧阴离子自由基是生物体内活性氧(ROS)的重要组成部分,当微藻细胞受到外界环境胁迫时,细胞内会产生大量ROS,而当机体自身清除自由基的能力不足时,就会导致氧化应激损伤,最终超氧阴离子自由基的含量也随之升高[20]。由图9可知,随着肌醇浓度的增加,超氧阴离子自由基的含量先降低后又升高,并在肌醇浓度为2.5 mg/L时达到最小值0.07µmol/mL,与对照组相比较降低了40.84%,而且肌醇的添加使得超氧阴离子自由基的含量始终低于对照组,说明肌醇能够抑制藻株X6胞内超氧阴离子自由基的生成,对微藻细胞具有一定的保护作用。
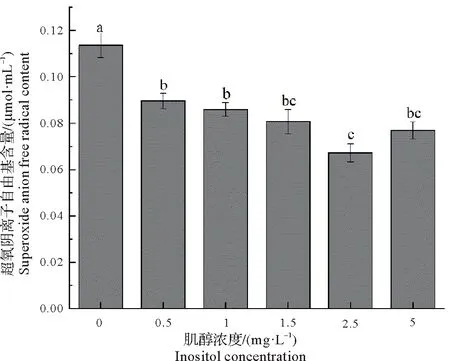
图9 不同浓度的肌醇对藻株X6超氧化物阴离子自由基含量的影响Fig.9 Effect of different concentrations of inositol on superoxide anion content of microalga X6
3 结论和讨论
微生物发酵过程中构建发酵动力学模型是一项成熟的工作,但关于探究不同培养模式下微藻生长、产物合成、基质消耗等动力学的研究不多。本文研究了藻株X6在不同培养条件下的生长动力学,对自养培养、异养培养和混养培养三种培养模式下藻株X6的生长、产物合成和底物消耗进行了实时观察,同时采用动力学模型(Logistic、Luedeking-Piret以及Luedeking-Piret修正方程)对异养培养和混养培养进行建模,结果表明藻株X6的油脂合成是部分生长偶联型且混养培养更有利于其生物量及脂质的积累。藻株X6进行自养培养时,其油脂含量和生物量随培养时间的延长变化不显著;而在混养培养和异养培养条件下,藻株X6的油脂含量及生物量急速增加,且混养培养条件下能获得最高的生物量和油脂含量,分别为14.97 g/L 和0.85 g/L。黎小廷[21]研究了混养培养和自养培养对顿顶螺旋藻生长的影响,结果表明混养培养更有利于藻株的生长与油脂的累积,这与本试验结果一致,但在该研究中混养培养条件下所获得的最大生物量为3.87 g/L,仅为本试验混养培养条件的25.85%,这也表明本论文所获藻株X6具有较为优良的生长特性,有进一步研究的价值。
肌醇化学结构类似葡萄糖,有研究表明其可作为促进微藻生长、提高代谢产物的碳源,在培养过程中加入如肌醇等多元醇可以使微生物在受到环境胁迫时提高其生长性能[22]。本试验在探究肌醇对藻株X6生长及生理影响时发现,当肌醇浓度为2.5 mg/L时,藻株X6的油脂含量最高,占细胞干重的14.09%,且在此条件下其总糖含量达到最大值140.11 mg/L,相对于对照组提高了87.54%,而此时蛋白质含量达到最小值0.58 mg/mL。有研究表明,微藻能够在光合碳划分方面发生程序性变化,因此细胞生化成分,特别是脂质、碳水化合物和蛋白质的含量会随环境和培养条件的变化而产生相应改变[23]。Qian等[24]也得到了类似的试验结果,小球藻添加肌醇后更多的碳和能量将用于碳水化合物和脂质的生物合成,这与试验结果中蛋白质含量降低而总糖含量与脂质含量提高的情况是一致的。丙二醛与超氧阴离子自由基含量可以作为判断外源物质对微藻毒害程度的指标,当微藻细胞受到胁迫等不利条件时,其含量会上升。有研究表明[25-26],外源添加肌醇可以增强抗氧化酶活性,抑制ROS的积累,保护细胞免受氧化应激损伤。而试验结果表明随着肌醇浓度的增加,丙二醛含量和超氧阴离子自由基含量逐渐降低,且始终低于对照组,说明肌醇在一定程度上降低了丙二醛及超氧阴离子自由基的含量,对微藻生物膜系统起到了一定的保护作用。
综上所述,通过不同培养条件下发酵动力学模型的构建,可知藻株X6是部分生长偶联型,在混养培养模式下有助于其生物量及脂质的积累。肌醇对藻株X6的生物量无显著促进作用,但能提高其油脂生产强度,添加肌醇的处理组与对照组相比较X6的胞内可溶性蛋白质、超氧阴离子自由基和丙二醛含量均出现降低趋势,且始终低于对照组,总糖含量升高,以上变化表明肌醇对微藻细胞具有一定的保护作用。
致谢:江西省教育厅科学研究项目(GJJ160388)和江西农业大学博士启动基金(9232305387)同时对本研究给予了资助,谨致谢意!
