NEAT1/miR-193a-3p通过TLR-4/NF-κB信号通路对脂多糖诱导的心肌H9c2细胞损伤的影响
2022-09-29曾羽霄庄少伟
曾羽霄,庄少伟
多种心肌疾病中均存在心肌细胞凋亡的现象。心肌细胞凋亡、炎症可造成急性心肌梗死(acute myocardial infarction,AMI)进一步加重并引发心肌功能障碍、心力衰竭等,导致病人死亡风险升高[1]。近年来的研究发现,长链非编码RNA(long noncoding RNA,lncRNAs)和短非编码RNA(miRNAs)在生物过程中发挥重要的作用。lncRNA是一类长度大于200多个核苷酸的转录RNA分子,参与调控细胞的增殖、凋亡、迁移和侵袭等生物学行为。NEAT1是一种长度约为3 700 nt的lncRNA,定位于11q13.1,与mRNA前体剪接区域 SC-35有密切联系,研究显示NEAT1参与调节细胞增殖、凋亡和迁移等多种癌症相关的细胞活动[2]。最近研究显示,NEAT1可通过产生肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)和白细胞介素-6(IL-6)促进炎症反应[3]。miRNA为18~25个核苷酸之间的内源性短链非编码RNA分子,主要结合 mRNA 的非翻译区,下调或者抑制mRNA的转录与翻译,进而调控下游靶基因的表达,参与各种疾病的病理过程[4]。miR-193a-3p来源于miR-193家族,其抗肿瘤活性已在肺癌和肾细胞癌等各种癌症研究中广泛报道[5]。竞争性内源RNA(ceRNA)假说的提出让人们对lncRNAs和miRNAs之间的作用机制有了新的认识。当前研究表明,lncRNA可作为ceRNA并竞争性结合到miRNA,从而调节mRNA的转录水平并调节基因表达[6]。已有研究显示,NEAT1作为ceRNA并结合miR-193a-3p以加速肺腺癌恶化[7]。但是NEAT1和miR-193a-3p在心肌细胞损伤是否存在某种调控关系及其作用机制尚不明确。本研究用脂多糖(LPS)处理心肌H9c2细胞建立心肌细胞损伤模型,探讨NEAT1/miR-193a-3p在心肌细胞损伤中的作用,并且分析其影响炎性损伤的潜在机制。
1 材料与方法
1.1 材料
1.1.1 细胞株 胚胎大鼠心肌H9c2细胞,购自上海匹拓生物科技有限公司。
1.1.2 主要试剂 脂多糖购自上海广锐生物科技有限公司;胎牛血清、DMEM培养液和胰蛋白酶购自美国Gibico公司;Trizol试剂和Lipofectamine 2000购自美国Invitrogen公司;细胞计数试剂盒-8(CCK-8)试剂盒、增强型化学发光(ECL)试剂盒和RIPA 蛋白裂解液购自中国上海碧云天生物技术有限公司;二喹啉甲酸(BCA)蛋白分析试剂盒购自中国南京碧云天生物技术有限公司;双萤光素酶报告基因检测试剂盒购自Promega;膜联蛋白V-异硫氰酸荧光素(annexin V-FITC)和碘化丙啶(propidium iodide,PI)试剂盒购自北京宝赛生物技术有限公司;实时荧光定量聚合酶链式反应(RT-qPCR)试剂盒购自日本TaKaRa公司;SYBR Green qPCR试剂盒购自美国Thermo Scientific公司;酶联免疫吸附试验(ELISA)试剂盒购自美国R&D Systems公司;pcDNA-NEAT1、NEAT1 control、miR-193a-3p mimic和miR-193a-3p control购自上海吉玛制药技术有限公司;PrimeScriptTM逆转录试剂盒、SYBR®Premix Ex TaqTMII购自日本TaKaRa公司;Super SignalTM化学发光底物购自美国Thermo Scientific公司;Toll样受体-4(Toll-like receptor-4,TLR-4)抗体、骨髓分化主要反应蛋白88(myeloid differentiation primary response 88,MYD88)抗体、核因子-κB p65(nuclear factor kappa-light-chain-enhancer of activated B,NF-κB p65)抗体、磷酸化NF-κB p65(p-NF-κB p65)抗体、β-actin兔单克隆抗体、辣根过氧化物酶-羊抗兔免疫球蛋白G(IgG)二抗均购自美国CST公司;TNF-α、白细胞介素-1β(interleukin-1β,IL-1β)、IL-6和白细胞介素-8(IL-8)试剂盒均购自江苏科晶生物科技有限公司。
1.1.3 主要仪器 -80 ℃冰箱购自日本SANYO公司;二氧化碳(CO2)恒温培养箱购自美国Precision公司;PT-3502A 酶标仪购自北京普天新桥技术有限公司;FACSAria flow流式细胞仪购自美国BD Biosciences公司;Step One PlusTMreal-time PCR System购自美国 Applied Biosystems公司;凝胶扫描分析系统购自英国 Syngene 公司;蛋白质免疫印迹法(Western Blot)电泳和转膜系统、ChemiDoc 成像系统购自美国BIO-RAD公司。
1.2 方法
1.2.1 双荧光素酶报告基因试验 通过LncBase(www.microrna.gr/LncBase)预测NEAT1的靶基因,发现miR-193a-3p和NEAT1的3′非编码区域(3′- untranslated region,3′ UTR)存在互补的核苷酸序列。分别将含有野生型NEAT1(NEAT1-WT)和突变型NEAT1(NEAT1-MUT)质粒和miR-193a-3p mimic或者 NC mimic共转染至H9c2细胞内,转染48 h后采用Bright- GloTM荧光素酶试剂盒确认荧光素酶活性,使用酶标仪检测荧光强度。
1.2.2 RNA结合蛋白免疫沉淀试验 按照生产商RNA结合蛋白免疫沉淀配套所用EZ-Magna rip Kit说明书进行操作。在DMEM培养液中加入EZ-Magna rip Kit的蛋白酶抑制剂和RNA裂解缓冲液进行细胞裂解。根据说明书进行RNA免疫沉淀。然后将抗Argonaute2(Ago2)抗体和正常小鼠IgG结合的磁珠孵育在RNA免疫沉淀缓冲液中进行孵育。然后加入细胞裂解液,用蛋白酶K缓冲液处理各标本,同时分离被沉淀的RNA。后续进行逆转录,采用聚合酶链式反应(PCR)法检测NEAT1和miR-193a-3p水平。
1.2.3 心肌细胞炎性损伤模型的建立 H9c2细胞用含10%胎牛血清、DMEM培养液于37 ℃、含有5%CO2的细胞培养箱中传代培养。收集对数生长期细胞,分别用不同浓度(0.0 μg/mL、0.5 μg/mL、1.0 μg/mL和2.0 μg/mL)的脂多糖刺激细胞24 h,然后通过CCK-8试验和流式细胞术测定各组细胞的活力,并且采用RT-PCR法测定细胞中NEAT1和miR-193a-3p mRNA的表达水平。选择合适的脂多糖进行后续试验。
1.2.4 细胞分组和转染 将胚胎大鼠心肌H9c2细胞用含10%胎牛血清、DMEM培养液于37 ℃、含有5%CO2的细胞培养箱中传代培养。收集对数生长期细胞,将细胞分为miR-193a-3p mimic组、miR-193a-3p control组、NEAT1组、NEAT1 control组、miR-193a-3p mimic + NEAT1组、正常对照组以及模型对照组,参照Lipofectamine 2000 说明书步骤进行转染操作,分别利用转染试剂 Lipofectamine 2000将miR-193a-3p mimic、miR-193a-3p control、pcDNA-NEAT1、NEAT1 control以及miR-193a-3p mimic+pcDNA-NEAT1转染至细胞中,正常对照组和模型对照组细胞未进行任何转染。
1.2.5 CCK-8试验 各组细胞经脂多糖处理24 h后,加入CCK-8试剂,用酶标仪检测各孔450 nm处光密度OD值。以时间为横坐标,OD值为纵坐标绘制细胞增殖曲线。
1.2.6 流式细胞术实验 各组细胞转染后常规培养6 h,除正常对照组外,其余各组用含脂多糖(1 μg/mL)的新鲜DMEM培养液培养24 h,离心收集细胞。用磷酸缓冲盐溶液(PBS)重悬细胞,离心弃去上清液,重复两次。使用100 μL的Binding Buffer悬浮细胞,再加入5 μL的 Annexin V-FITC和5 μL的荧光染料碘化甲啶,避光孵育15 min后,用Binding Buffer重悬细胞,使用流式细胞仪检测各组细胞的凋亡率。
1.2.7 RT-PCR实验 各组细胞转染后常规培养6 h,除正常对照组外,其余各组用含脂多糖(1 μg/mL)的新鲜DMEM培养液培养24 h,离心收集细胞。加入Trizol 液提取总RNA,异丙醇法浓缩 RNA,采用全自动酶标仪测定总RNA纯度,控制A260/A280在1.9~2.1,RNA 电泳检测其完整性。检测浓度及纯度后取2 μg总RNA应用逆转录试剂盒合成第一链cDNA。逆转录条件如下:37 ℃ 反应15 min,85 ℃反应5 s。将逆转录所得cDNA纯化后应用Step One PlusTMreal-time PCR System进行PCR扩增。反应总体积为20 μL∶1.5 μL cDNA 模板,1 μL 引物,10 μL SYBR®Premix Ex TaqTMII,7.5 μL H2O。PCR反应条件为:95 ℃预变性30 s,扩增循环 95 ℃ 5 s, 60 ℃ 34 s,共40 个循环。以甘油醛-3-磷酸脱氢酶(GAPDH)为参照,采用2-ΔΔCt法计算mRNA的相对定量表达量。
1.2.8 Western Blot实验 各组细胞转染后常规培养6 h,除正常对照组外,其余各组用含脂多糖(1 μg/mL)的新鲜DMEM培养液培养24 h,离心收集细胞。用RIPA裂解液裂解各组细胞,离心,使用 BCA 法测定蛋白浓度。蛋白变性后按每孔20 μg进行10%聚丙烯酰胺凝胶(SDS-PAGE)凝胶电泳(90 V 30 min,120 V 1 h)。电泳后湿法转至聚偏二氟乙烯膜(PVDF,400 mA,2h),用5%BSA封闭1 h,TBST缓冲液洗膜,分别加入一抗后4 ℃过夜,TBST缓冲液充分洗膜,加入辣根过氧化物酶标记的二抗后室温下孵育1 h,TBST缓冲液充分洗膜30 min,用化学发光试剂盒显色,显影。测定各组细胞目标蛋白表达量,以与β-actin条带相对表达量表示。
1.2.9 ELISA试验 各组细胞转染后常规培养6 h,除正常对照组外,其余各组用含脂多糖(1 μg/mL)的新鲜DMEM培养液培养24 h,收集各组细胞上清液,按照ELISA试剂盒说明书要求操作,测定各组细胞上清液中TNF-α、IL-1β、IL-6和IL-8含量。

2 结 果
2.1 miR-193a-3p和NEAT1靶向的关系 在转染miR-193a-3p mimic的H9c2细胞中野生型NEAT1报告基因的荧光素酶活性明显降低(P<0.05),而突变报告基因的荧光素酶活性无明显变化(P>0.05)。荧光素酶报告基因结果证实NEAT1 可以作为 ceRNA 靶向结合miR-193a-3p。详见图1。

与突变型序列(3′URT-MUT)比较,*P<0.01。图1 H9c2细胞转染miR-193a-3p mimic后野生型和突变型NEAT1报告基因荧光素酶活性比较
2.2 免疫沉淀试验结果 与IgG免疫沉淀物对照相比,H9c2细胞中miR-193a-3p和NEAT1在Ago2免疫沉淀物中富集。结合荧光素酶报告基因结果,进一步证明miR-193a-3p是H9c2细胞中NEAT1的结合靶点。详见图2。
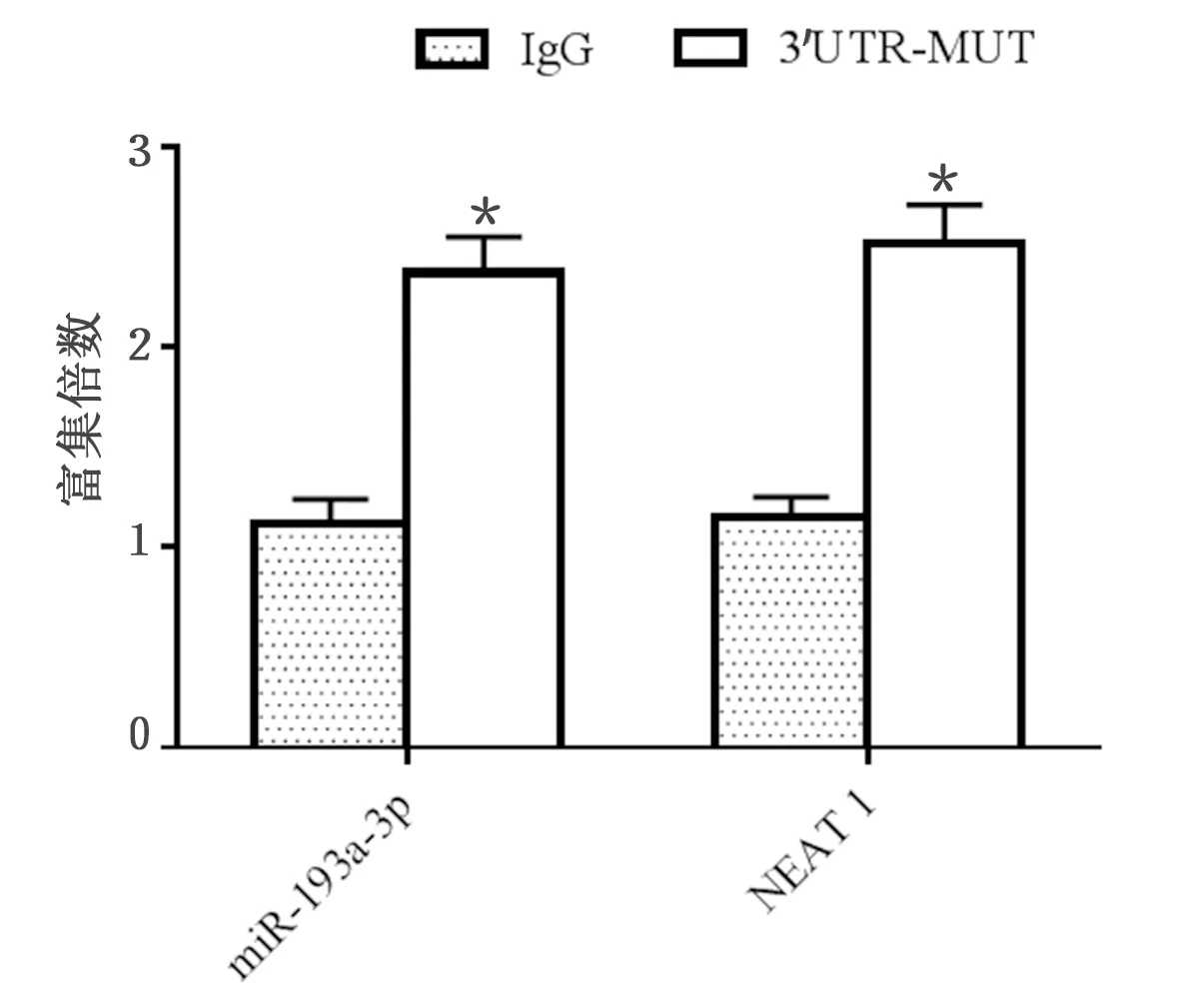
与IgG免疫沉淀物对照比较,* P<0.01。图2 免疫沉淀试验结果
2.3 脂多糖对H9c2细胞活力、NEAT1和miR-193a-3p mRNA表达的影响 当脂多糖浓度达到1 μg/mL和2.0 μg/mL时H9c2细胞活力明显降低(P<0.05或P<0.01),特别当脂多糖浓度达到2.0 μg/mL时H9c2细胞活力降低超过50%。随着脂多糖浓度的增加,H9c2细胞中miR-193a-3p mRNA表达水平明显降低(P<0.01),而NEAT1 mRNA表达水平明显升高(P<0.01),因此,NEAT1和miR-193a-3p可能参与脂多糖诱导的H9c2细胞损伤。详见图3~图5。

与0.0 μg/mL比较,*P<0.05,#P<0.01。图3 脂多糖对H9c2细胞活力的影响(n=6)

与0.0 μg/mL比较,*P<0.01。图4 脂多糖对H9c2细胞miR-193a-3p mRNA表达的影响(n=6)

与0.0 μg/mL比较,*P<0.01。图5 脂多糖对H9c2细胞NEAT1 mRNA表达的影响(n=6)
2.4 各组H9c2细胞中NEAT1和miR-193a-3p mRNA表达比较 将miR-193a-3p mimic转染至H9c2细胞中可使miR-193a-3p mRNA表达水平明显增加(P<0.01),同时可使NEAT1 mRNA表达水平明显降低(P<0.01);而将pcDNA-NEAT1转染至H9c2细胞中可使NEAT1 mRNA表达升高(P<0.01),同时可使miR-193a-3p mRNA表达水平明显降低(P<0.01)。详见图6。

模型对照组与正常对照组比较,*P<0.01;与模型对照组比较,#P<0.01。图6 各组H9c2细胞中NEAT1和miR-193a-3p mRNA表达比较(n=6)
2.5 NEAT1/miR-193a-3p对脂多糖所致H9c2细胞活力和细胞凋亡的影响 将miR-193a-3p mimic 转染至H9c2细胞后可改善H9c2细胞因脂多糖刺激导致的细胞活力降低和细胞凋亡增加(P<0.01),而将pcDNA-NEAT1转染至H9c2细胞可逆转miR-193a-3p mimic对细胞活力和凋亡的调节作用(P<0.01)。因此,过表达miR-193a-3p可改善H9c2细胞因脂多糖刺激导致的细胞活力降低和细胞凋亡增加,而过表达NEAT1可逆转miR-193a-3p对细胞活力和凋亡的调节作用。详见图7、图8。

模型对照组与正常对照组比较,*P<0.01;与模型对照组比较,#P<0.01;与miR-193a-3p mimic组比较,△ P<0.01。图7 NEAT1/miR-193a-3p对脂多糖所致H9c2细胞活力的影响(n=6)

模型对照组与正常对照组比较,*P<0.01;与模型对照组比较,#P<0.01;与miR-193a-3p mimic组比较,△ P<0.01。图8 NEAT1/miR-193a-3p对脂多糖所致H9c2细胞凋亡的影响(n=6)
2.6 NEAT1/miR-193a-3p对脂多糖所致H9c2细胞分泌炎性因子的影响 将miR-193a-3p mimic 转染至H9c2细胞后可使升高的TNF-α、IL-1β、IL-6和IL-8水平明显降低(P<0.01),而将pcDNA-NEAT1转染至H9c2细胞可逆转miR-193a-3p mimic对TNF-α、IL-1β、IL-6和IL-8的调节作用(P<0.01)。因此,过表达miR-193a-3p可降低H9c2细胞因脂多糖刺激导致的TNF-α、IL-1β、IL-6和IL-8水平升高,而过表达NEAT1可逆转miR-193a-3p对TNF-α、IL-1β、IL-6和IL-8水平的调节作用。详见图9。

模型对照组与正常对照组比较,*P<0.01;与模型对照组比较,#P<0.01;与miR-193a-3p mimic组比较,△ P<0.01。图9 NEAT1/miR-193a-3p对脂多糖所致H9c2细胞炎性因子分泌的影响(n=6)
2.7 NEAT1/miR-193a-3p对脂多糖所致H9c2细胞中TLR-4/NF-κB信号通路相关蛋白表达的影响 将miR-193a-3p mimic 转染至H9c2细胞后可使升高的TLR-4、MYD88和p-NF-κB p65蛋白表达水平明显降低(P<0.01),而将pcDNA-NEAT1转染至H9c2细胞可逆转miR-193a-3p mimic对TLR-4、MYD88和p-NF-κB p65蛋白表达的调节作用(P<0.01),但是miR-193a-3p mimic 和pcDNA-NEAT1转染对NF-κB p65蛋白表达水平无明显影响(P>0.05)。因此,过表达miR-193a-3p可降低H9c2细胞因脂多糖刺激导致的TLR-4、MYD88和p-NF-κB p65蛋白表达水平升高,而过表达NEAT1可逆转miR-193a-3p对TLR-4、MYD88和p-NF-κB p65蛋白表达水平的调节作用,但是过表达miR-193a-3p或过表达NEAT1对NF-κB蛋白表达水平无影响。详见图10。

模型对照组与正常对照组比较,*P<0.01;与模型对照组比较,#P<0.01;与miR-193a-3p mimic组比较,△ P<0.01。图10 NEAT1/miR-193a-3p对脂多糖所致H9c2细胞中TLR-4/NF-κB信号通路相关蛋白表达的影响(n=6)
3 讨 论
多种miRNA可通过调节心肌细胞炎症、纤维化、细胞自噬、细胞凋亡及新生血管形成的表型机制参与心脏疾病的发生和发展[8-9]。Eguchi等[10]研究显示,心肌细胞外泌体中的miRNA-214具有抗细胞凋亡作用,心肌细胞通过网格蛋白介导的内吞作用捕获miRNA-214从而抑制心肌细胞凋亡。miRNA-488-3p 通过靶向ZNF791 抑制急性心肌梗死后的心肌细胞凋亡[11]。miR-325-3p 通过受体相互作用蛋白激酶1(RIPK1)/受体相互作用蛋白激酶3(RIPK3)/人磷酸化混合系列蛋白激酶样结构域(p-MLKL)途径抑制坏死性凋亡,抑制RIPK3的表达和程序性坏死,保护心肌梗死后的心脏[12]。miR-193a-3p来源于miR-193家族,其抗肿瘤活性已广泛报道,但是在心肌细胞损伤中的作用鲜有报道。lncRNA 是一种非编码RNA,在真核细胞内被普遍转录,其表达具有组织特异性,与动脉粥样硬化、慢性心力衰竭等心血管疾病的发生发展密切相关。随着对lncRNA的深入研究,许多研究结果表明其在炎症介质产生及炎症细胞分化、迁移等多种生物过程中起重要作用。Zhang等[13]报道抑制NEAT1可明显降低脂多糖诱导的趋化因子和细胞因子的表达。张飞飞等[14]探索NEAT1在先天免疫反应中的作用,结果表明NEAT1参与Toll样受体-2(TLR-2)介导的先天免疫调节,并且选择性地调控TLR-2信号活化缓慢持续诱导的晚期反应炎性因子的表达。Chen等[15]研究显示,NEAT1可通过调节NF-κB通路影响脓毒症诱导的急性肾细胞凋亡和炎症反应。
lncRNA可作为ceRNA并竞争性结合到miRNA,而NEAT1作为ceRNA并结合miR-193a-3p[7]。本研究通过荧光素酶报告基因结果证实NEAT1 可以作为ceRNA 靶向结合miR-193a-3p,并且免疫沉淀试验进一步证明miR-193a-3p是H9c2细胞中NEAT1的结合靶点。目前,关于NEAT1和miR-193a-3p在心肌细胞损伤的作用鲜有报道。本研究结果显示,脂多糖可致H9c2细胞活力明显降低,并且细胞中miR-193a-3p mRNA表达水平明显降低(P<0.01),而NEAT1 mRNA表达水平明显升高(P<0.01),说明NEAT1和miR-193a-3p可能参与脂多糖诱导的H9c2细胞损伤。通过转染miR-193a-3p mimic或pcDNA-NEAT1至H9c2细胞,改变细胞中miR-193a-3p mRNA和NEAT1 mRNA的表达水平,过表达miR-193a-3p可改善H9c2细胞因脂多糖刺激导致的细胞活力降低和细胞凋亡增加,而过表达NEAT1可逆转miR-193a-3p对细胞活力和凋亡的调节作用。IL-6、TNF-α等炎性因子水平升高导致心肌功能紊乱及心肌细胞凋亡、坏死[16]。炎症反应是包括心肌细胞在内的多种细胞凋亡、坏死的重要病理改变,降低细胞炎性因子含量有助于减轻细胞损伤。本研究结果显示,过表达miR-193a-3p可降低H9c2细胞因脂多糖刺激导致的TNF-α、IL-1β、IL-6和IL-8水平升高,而过表达NEAT1可逆转miR-193a-3p对TNF-α、IL-1β、IL-6和IL-8水平的调节作用。因此,miR-193a-3p和NEAT1在H9c2细胞损伤中发挥相反的作用,NEAT1可反向调节miR-193a-3p在心肌细胞损伤中的作用。
越来越多的研究表明,TLR-4/NF-κB信号通路可调节细胞增殖和细胞凋亡,是涉及免疫和炎症反应的重要细胞信号传导通路,并且是细胞凋亡引起炎症性损伤的原因[17]。TLR-4是一种在天然免疫和炎症反应中起重要作用的模式识别受体,革兰阴性菌可被TLR-4特异性识别,并激活TLR-4招募MYD88,导致NF-κB抑制蛋白IκBα磷酸化,NF-κB从复合物上解离,最后NF-κB 进入细胞核内与靶基因结合,刺激TNF-α、IL-6 和IL-1β等炎性因子的生成[18],进一步诱导炎症反应[19],导致血管内皮细胞功能障碍和组织氧化损伤,进一步促使血管内皮细胞衰老、死亡,诱导血管内皮功能紊乱,进而诱发动脉粥样硬化、冠心病等心血管疾病[20]。使TLR-4/NF-κB信号通路失活一直是抑制炎症的有效方法和治疗靶点。Zhang等[21]研究显示,聚乙二醇可通过阻断TLR-4/NF-κB改善脂多糖诱导的神经炎症。下调lncRNA MALAT1可通过抑制TLR-4/NF-κB介导的炎症反应,减轻饱和脂肪酸诱导的心肌炎症损伤[22]。Pan等[23]研究显示,miR-21可抑制TLR-4/NF-κB通路,以减少心肌细胞凋亡和炎性因子生成。在本研究中,过表达miR-193a-3p可降低H9c2细胞因脂多糖刺激导致的TLR-4、MYD88和p-NF-κBp65蛋白表达水平升高,而过表达NEAT1可逆转miR-193a-3p对TLR-4、MYD88和p-NF-κBp65蛋白表达水平的调节作用,因此,miR-193a-3p可通过调节TLR-4/NF-κB信号通路影响脂多糖诱导的细胞凋亡和炎症反应,而NEAT1可通过调节miR-193a-3p发挥其作用。因此,NEAT1可能通过海绵化miR-193a-3p来调节TLR-4/NF-κB信号通路的活化,从而减轻脂多糖诱导的炎症和细胞凋亡,从而影响心肌细胞损伤。
总之,NEAT1通过miR-193a-3p可影响脂多糖诱导的心肌细胞凋亡和炎症反应,提示NEAT1可作为竞争性内源RNA海绵miR-193a-3p参与心肌损伤过程,并且与调控TLR-4/NF-κB信号通路有关。
