高能重离子束和X射线辐射水稻幼苗的当代生物学效应研究
2022-09-21许超丽任维宾王圆梦司怀军周利斌
许超丽 刘 霄 任维宾 杜 艳 王圆梦 李 燕 司怀军 周利斌,3,*
(1 甘肃农业大学生命科学技术学院,甘肃 兰州 730070;2 中国科学院近代物理研究所生物物理室,甘肃 兰州 730030;3 中国科学院大学,北京 100049;4 甘肃中医药大学药学院,甘肃 兰州 730030;5 四川农业大学西南作物基因资源发掘与利用国家重点实验室,四川 成都 611130)
水稻(Oryzasativa)作为世界上最主要的粮食作物之一[1],其新品种的选育与创新对世界粮食安全有着重要意义。随着辐射技术在水稻育种中的广泛应用,对其诱变机理的研究也日渐增多。物理诱变育种常用的电离辐射包括X射线、γ射线、快中子及重离子束等[2]。电离辐射能够诱导植物产生多种生物学效应,包括刺激效应、生理损伤以及遗传突变等[3]。然而,目前在辐射机理、突变筛选及相关分子机制等方面的基础研究报道相对较少,因此对不同类型辐射诱变机理的研究能够更好地了解辐射对植物造成的当代生理损伤及后代遗传突变情况。同时,加强辐射品质与诱变分子机理的研究可以为水稻高效诱变育种提供一定的理论基础。
高能重离子束辐射被认为是一种高效的物理诱变技术[4]。重离子束在水稻育种中的实用性及有效性是由于其具有传能线密度(linear energy transfer,LET)大和相对生物学效应(relative biological effectiveness,RBE)高等物理学及生物学特征[5],在育种过程表现出突变率高[6]、突变谱广[7]和育种周期相对较短等特点[8]。重离子束辐射对水稻生长发育的影响已有报道。王佳琪等[9]研究表明,碳离子束辐射不同粳稻干种子后,其发芽率和存活率存在差异,即不同品种粳稻的辐射敏感性不同;Xu等[10]利用不同能量的重离子辐射3个品种的水稻干种子,发现低能氮离子和中能碳离子束辐射对发芽率、存活率、苗高等当代生物学指标的影响不同。X射线是一种低LET辐射,通常由电子加速器加速的电子撞击金属靶产生,具有操作简便、使用成本较低、无需使用放射源等特点[11],在诱变育种中也有广泛应用。高能重离子束属带电粒子辐射,X射线属电磁波,两者与物质相互作用的原理不同,辐射诱导的植物生理生化变化及遗传突变也各不相同。已有研究表明,在植物诱变育种操作中,X射线单位剂量的诱变效率较重离子束低约10倍[12]。
植物经电离辐射处理后,其生理生化改变及分子响应过程较为复杂,研究植物经辐射处理后的形态学及生理学改变,能够更好地了解农作物对不同类型电离辐射的响应机制。前人研究多关注于辐射干种子对植物生长的影响,Zhang等[13]将粳稻和籼稻种子进行γ辐射,发现籼稻比粳稻更耐受辐射。关于重离子束辐射植物幼苗的诱变效应研究,Hase等[14]对拟南芥种子和幼苗分别进行碳离子束辐射,发现干种子的突变率是幼苗的1.4~1.9倍。对水稻幼苗进行辐射处理所产生的当代损伤效应的研究较少,关于高能碳离子束和X射线两种不同类型电离辐射对水稻幼苗生理损伤的比较研究更是鲜有报道。因此,本研究以水稻幼苗为起始材料,以高能碳离子束进行诱变处理,以低LET辐射的代表X射线处理为诱变参照,研究不同类型电离辐射处理对水稻生长、发育以及生理损伤的影响,以期为不同类型电离辐射诱变水稻时,辐射剂量及样品初始状态的选择提供理论依据。
1 材料与方法
1.1 植物材料
粳稻(OryzasativasspJaponica)Kitaake是日本特早熟品种,也是水稻研究的模式物种,从播种至开花平均仅需52个生育日。本研究中水稻材料源自四川农业大学西南作物基因资源发掘与利用国家重点实验室。
1.2 样品制备
试验于2021年4月进行,选取无病害、饱满度一致且同一批次的Kitaake水稻种子进行萌发,将水稻干种子置于浓度为3%的过氧化氢溶液中,消毒30 min后用去离子水冲洗3~5次,然后置于含湿润滤纸直径90 mm圆形培养皿中,培养5 d,其中1~4 d置于无光照恒温培养箱(32℃)中萌发,后移至光照培养间(温度为28~30℃,光周期光/暗为14/10 h,光强为250 μmol·m-2·s-1)中生长1 d。选取萌发一致的幼苗置于有湿润滤纸的35 mm圆形培养皿中,室温下进行辐射处理。
1.3 辐射处理
对5日龄幼苗进行辐射处理,高能12C6+束流由中国科学院近代物理研究所兰州重离子研究装置(Heavy Ion Research Facility in Lanzhou,HIRFL)提供,能量为80.55 MeV·u-1,LET为34 keV·μm-1,剂量率约为60 Gy·min-1,辐射剂量分别为5、10、15、20、25、30、40 Gy。X射线辐射处理采用X-RAD255 X射线辐照仪(Precision X-Ray,美国)进行,剂量率约为6.5 Gy·min-1,辐射剂量分别为10、20、30、40、50、60、80 Gy。对照置于与处理组相同的环境条件下但不进行辐射处理。
1.4 辐射后培养
辐射后将幼苗立即转移至水培盒中培养,培养条件为:纯水培养2 d,1/4×营养液培养3 d,1/2×营养液培养3 d,1×营养液(Coolaber营养液:1 000×硅酸钠,1 000×大量元素微量元素混合液,1 000×铁盐,1 000×钙盐)进行培养,水培盒置于光照培养间中。40 d后随机选取部分植株进行插秧种植,种植于直径为20 cm花盆中,每盆3株。
1.5 测定指标与方法
1.5.1 营养生长时期形态指标的测定 对辐射处理后6 d的植株进行根长和芽长测定。根长为从根尖到幼苗底部的距离,芽长为幼苗底部到芽尖的距离,相对根长=处理组平均根长÷对照平均根长×100%。每组处理选取20~30棵幼苗进行数码拍照并使用Image J软件测量长度。
1.5.2 生殖发育时期形态指标的测定 在植株成熟后,对植株株高、分蘖数、结实率以及千粒重进行统计。株高:从根部到植株顶端最长叶的长度;分蘖数:从水稻基部数水稻分蘖个数;结实率:每株选取10穗,结实率=实粒数/总粒数×100%;千粒重:用于统计育性的所有种子的重量,并换算为千粒重。每组均测定6株单株。
1.5.3 光合作用指标的测定 辐射后第30天对植株进行叶绿素含量测定,3株水稻混合取样,取样部位为新鲜水稻嫩叶距叶尖3~8 cm处。用乙醇提取法[15]测定叶片叶绿素含量。根据公式计算叶片单位鲜重的叶绿素a(chlorophyll a,Chla)、叶绿素b(chlorophyll b,Chlb)和总叶绿素(total chlorophyll)含量。辐射后第30天,对植株进行叶绿素荧光动力学参数测定,使用3~4 株水稻叶片混合测定,先将水稻暗适应20 min,选取植株第一、第二叶的距叶尖5~10 cm部位进行活体检测,用DUAL-PAM-100型叶绿素荧光仪(Walz,德国)在黑暗的环境中检测,并记录光系统Ⅱ(PSⅡ)最大光化学量子产量值(maximum photochemical efficiency,Fv/Fm)以及有效光化学量子产量值[actual photochemical efficiency,Y(Ⅱ)]。
1.5.4 抗氧化酶活性指标的测定 辐射后6、24、72 h对20 Gy高能碳离子束及40 Gy X射线处理组幼苗地上部分和根分别取样,未辐射组为对照,称量鲜重后液氮快速冷冻,并置于-80℃冷冻保存,用于后续抗氧化系统的分析。抗氧化酶活性测定:过氧化物酶(peroxidase,POD)活性,采用愈创木酚法在470 nm波长下检测吸光值;超氧化物歧化酶(superoxide dismutase,SOD)活性,采用氮蓝四唑(nitro-blue tetrazolium,NBT)光化学还原法在560 nm波长下检测吸光值;过氧化氢酶(catalase,CAT)活性,采用过氧化氢分解法在240 nm波长下检测光密度[16]。
1.5.5 遗传物质多态性分析 水稻抽穗后,取主穗旗叶进行遗传物质多态性的分析,以对照组单株水稻的基因组DNA为材料,使用CTAB法[17]提取主穗旗叶的DNA,并于-20℃条件下保存备用。利用简单重复间序列(inter-simple sequence repeat,ISSR)进行DNA多态性检测,共选用UBC系列引物29条(引物来源:University of British Columbia)进行条带扩增;检测扩增产物,选取扩增条带数多、稳定和条带清晰的引物用于后续研究,PCR扩增后,使用1.5%琼脂糖凝胶进行电泳,设置电压为110 V,时间为40 min,电泳完成后使用紫外凝胶系统成像,并进行多态性条带统计。
1.6 数据分析
试验重复3次,所得数据用平均值±标准差表示。差异显著性分析使用SPSS统计软件,采用单因素方差(One-way ANOVA)分析统计学差异,采用Duncan’s test;两组均值比较采用Student t-test检验。使用Origin软件、Excel 2016软件绘图。
2 结果与分析
2.1 高能碳离子束和X射线辐射对水稻幼苗生长发育的影响
由图1可知,两种辐射根长和苗高都随辐射剂量的增加,整体呈现降低趋势。对于高能碳离子束辐射处理组,根长在辐射剂量为15 Gy时比对照显著降低25.42%(图1-A);而对于苗高,辐射剂量为5 Gy时与对照相比显著降低(图1-B),表明地上部分对辐射的响应较根部更为敏感。对于X射线辐射组,根长在辐射剂量为20 Gy时与对照相比显著降低21.02%;苗高在辐射剂量为10 Gy时与对照相比显著增加7.38%(图1-C),这可能与低剂量辐射对植株的刺激效应有关[18];然而辐射剂量到20 Gy后一直呈降低趋势(图1-D)。
水稻植株的生长需要完整、强大的根系来吸收水分和无机盐,从而为植株积累养分,以供植株正常的生长发育。据图1计算可知,在高能碳离子束辐射剂量为20 Gy、X射线辐射剂量为40 Gy时,植株的相对根长最为接近,分别为69.54%和 68.53%。这两个剂量处理下水稻根部未完全受损,幼苗可以生长,并且与对照相比其根长又显著降低,因此选择这两个剂量进行后续研究。
2.2 高能碳离子束和X射线辐射对水稻主要农艺性状的影响
2.2.1 高能碳离子束和X射线辐射对水稻株高、分蘖数的影响 由于40 Gy高能碳离子束和80 Gy X射线辐射处理,使得水稻幼苗受到严重损伤而生长停滞,故无法进行主要农艺性状的统计分析。由图2可知,两种不同类型的电离辐射处理对成熟后水稻的株高影响不大,仅发现15 Gy高能碳离子束辐射处理组株高较对照略有增加且差异显著,其余各剂量的处理组与对照组相比均无显著性差异(图2-A);所有X射线辐射处理组与对照组的株高均无显著性差异(图2-C)。本试验结果发现辐射处理水稻幼苗后,其成熟期植株的各处理组间分蘖数差异较大。总体来看,在高能碳离子束辐射组中,不同剂量处理组之间的水稻分蘖数较对照无显著性差异(图2-B);而在X射线辐射组中,与对照相比,辐射剂量为50、60 Gy时,分蘖数显著和极显著增加(图2-D)。可知在X射线辐射处理下,较高的剂量对水稻植株分蘖能力造成了影响。
2.2.2 高能碳离子束和X射线辐射对水稻结实率、千粒重的影响 高能碳离子束和X射线辐射水稻幼苗的结实率、千粒重从整体上与对照相比呈下降趋势。由表1可知,高能碳离子束辐射剂量为5、10 Gy时,结实率及千粒重与对照相比无显著差异,15 Gy时出现显著降低,由于辐射处理组各单株有较大差异,导致出现特例,在辐射剂量为20 Gy时统计学上差异不显著。由表2可知,辐射的水稻幼苗经10、20、30 Gy X射线处理后,其结实率及千粒重与对照相比均无显著差异,在辐射剂量达到40 Gy后出现下降,且差异极显著,40 Gy X射线处理组的结实率不足对照组的50%。50、60 Gy的X射线辐射对水稻的生殖发育造成了严重的影响,造成M1植株的不育,无法产生后代。
2.3 高能碳离子束和X射线辐射对水稻叶片光合作用的影响
2.3.1 高能碳离子束和X射线辐射对水稻叶片叶绿素含量的影响 由图3可知,20 Gy高能碳离子束处理下,水稻叶片中Chla、Chlb和总叶绿素的含量与对照相比均无显著差异;在40 Gy X射线处理下,叶片中Chla、Chlb和总叶绿素的含量较对照均极显著提高。表明在根长抑制相似的情况下,经X射线辐射的水稻幼苗在处理后第30天,叶片中光合色素积累多,植株的光合作用能力强。
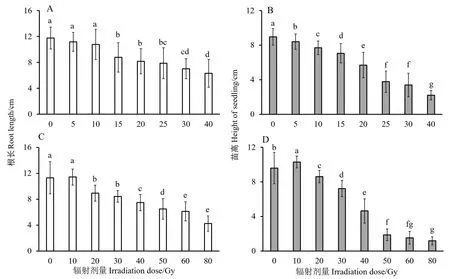
注: 不同小写字母表示不同辐射剂量之间差异显著(P<0.05)。Note: Different lowercase letters indicate significant differences among different irradiation does at 0.05 level.图1 高能碳离子束(A、B)和X射线(C、D)辐射对水稻幼苗根长和苗高的影响Fig.1 Effects of high-energy carbon ion beam(A、B) and X-rays(C、D) radiations on the root length and seedling height of rice
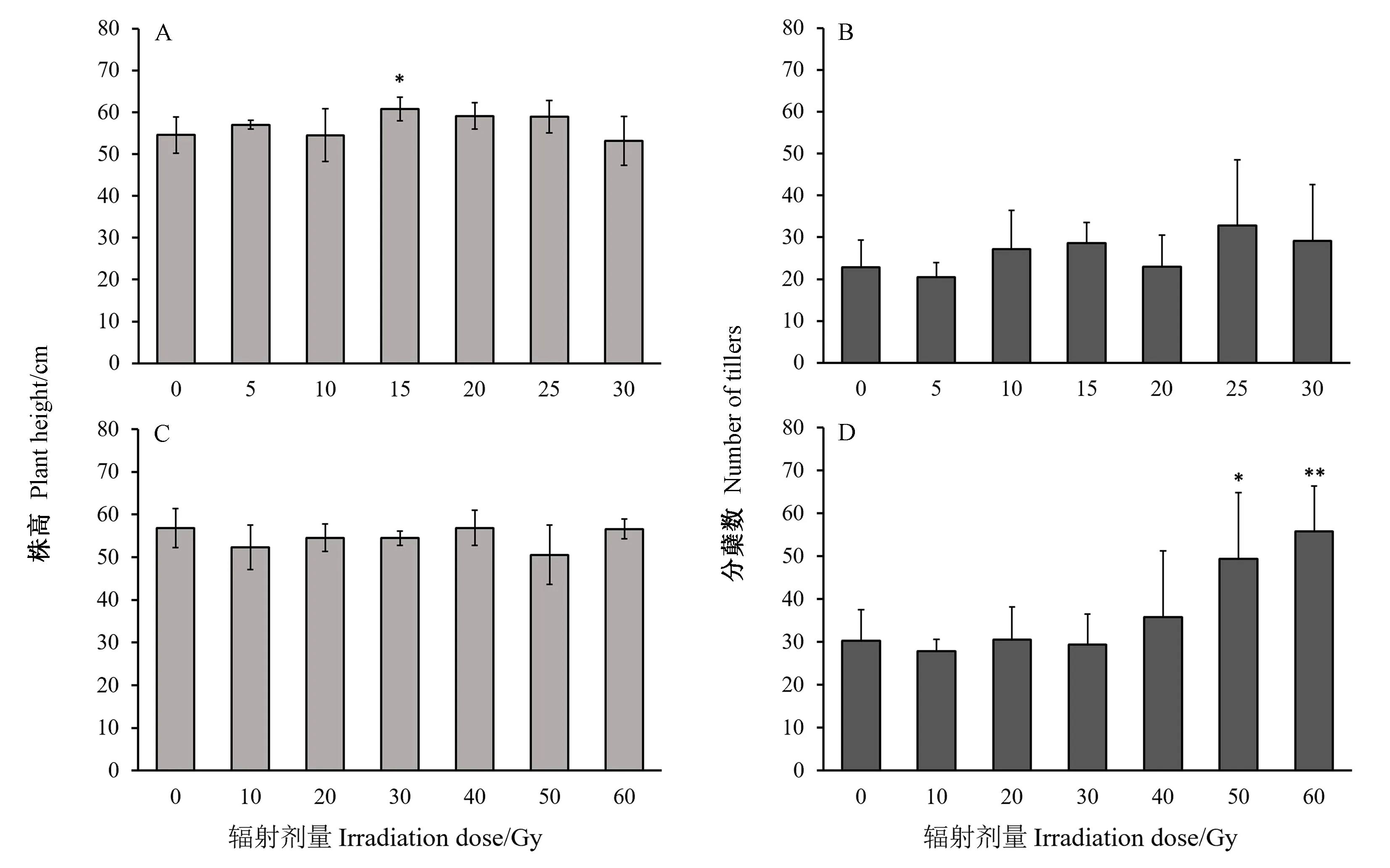
注:*表示与对照相比差异显著(P<0.05);**表示与对照相比差异极显著(P<0.01)。下同。Note:* means significant difference between treatments and control at 0.05 level. ** means extremely significant difference between treatments and control at 0.01 level. The same as following.图2 高能碳离子束(A、B)和X射线(C、D)辐射对水稻株高及分蘖数的影响Fig.2 Effect of high-energy carbon ion beam(A、B) and X-ray(C、D) radiations on rice plant height and number of tillers
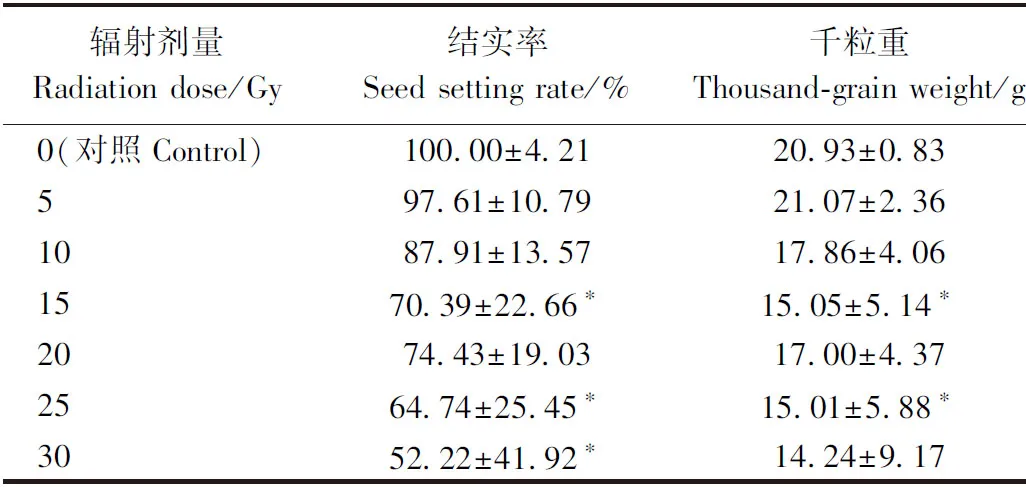
表1 高能碳离子束辐射对水稻结实率、千粒重的影响Table 1 Effects of high-energy carbon ion beam radiation on rice fertility and thousand-grain weight
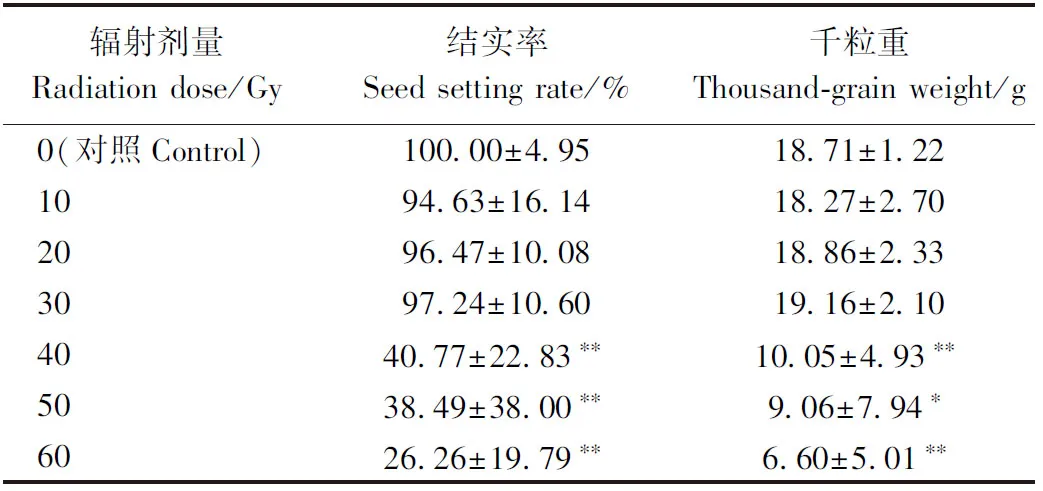
表2 X射线辐射对水稻育性、千粒重的影响Table 2 Effect of X-ray radiation on rice fertility and thousand-grain weight
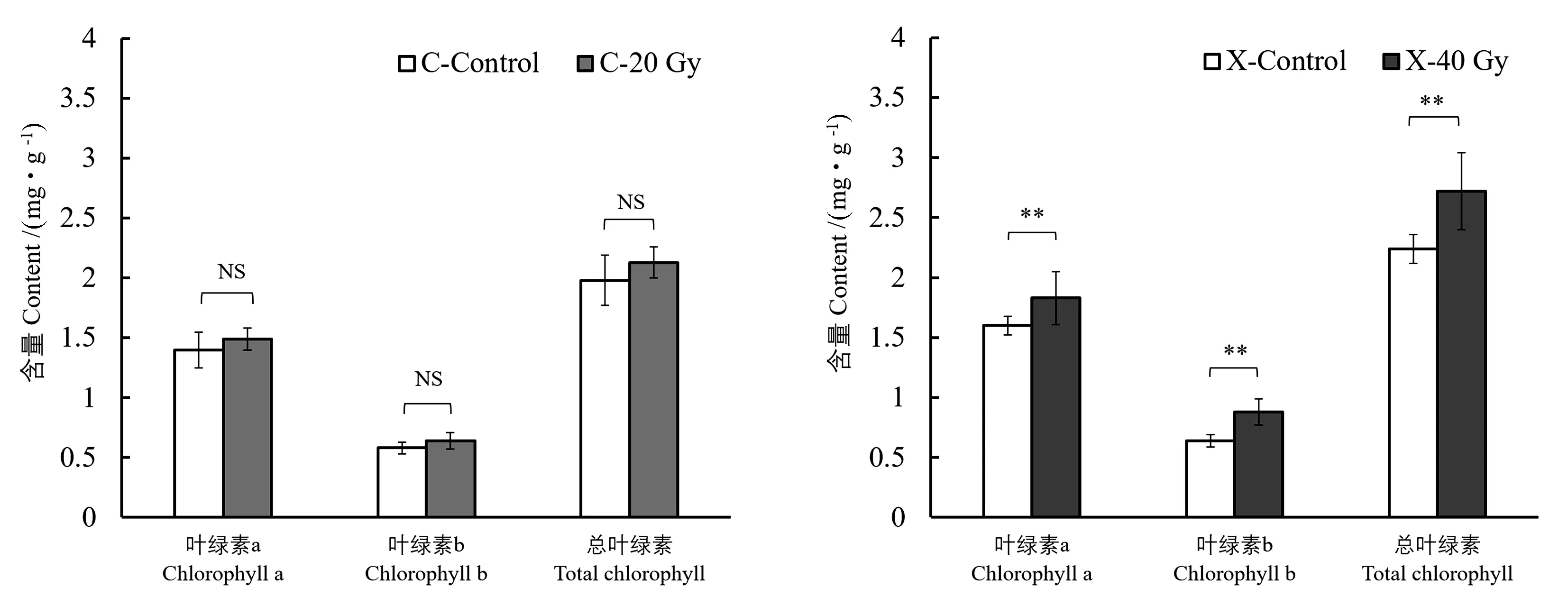
注:NS表示对照与辐射处理间无显著差异;**表示对照与辐射处理间差异极显著(P<0.01)。C-Control表示高能碳离子束处理组的对照,C-20 Gy表示高能碳离子束辐射处理剂量为20 Gy;X-Control表示X射线处理组的对照;X-40 Gy表示X射线辐射处理剂量为40 Gy。Note: NS means no significant difference among Control and irradiation treatments. ** means significant difference at 0.01 level among Control and irradiation treatments. C-Control represents the control of the high-energy carbon ion beam treatment group. C-20 Gy represents the high-energy carbon ion beam radiation treatment dose of 20 Gy. X-Control represents the control of the X-ray treatment group. X-40 Gy represents the X-ray radiation treatment dose of 40 Gy. 图3 高能碳离子束和X射线辐射对水稻叶片绿素含量的影响Fig.3 Effect of high-energy carbon ion beam and X-ray radiation on the chlorophyll content of rice leaves
2.3.2 高能碳离子束和X射线辐射对水稻叶片叶绿素荧光参数的影响 由表3可知,40 Gy X射线处理组Fv/Fm值较对照显著降低,20 Gy高能碳离子束则无显著变化,表明水稻叶片PSⅡ最大光能利用能力与所受的辐射类型相关。
2.4 高能碳离子束和X射线辐射对水稻幼苗抗氧化系统的影响
由图4可知,经高能碳离子束辐射后,水稻幼苗地上部分抗氧化酶活性整体高于根系,根系SOD活性在辐射处理后6、24、72 h出现先上升后下降再上升的波动,地上部分SOD活性仅在辐射处理后72 h出现显著上升(图4-A)。POD活性无论是根系还是地上部分都随着辐射后时间延长呈上升趋势(图4-C)。根系CAT活性在辐射后6 h出现一个峰值;而在芽中,CAT活性在辐射后24 h出现显著降低,到72 h时又升高(图4-E)。

表3 高能碳离子束和X射线辐射的水稻叶片绿素荧光特征比较Table 3 Comparison of chlorophyll fluorescence characteristics of rice leaves irradiated by high-energy carbon ion beam and X-ray
经X射线辐射处理水稻幼苗,根系SOD活性在处理后24 h出现峰值,到72 h时活性又下降至与对照无显著差异;而地上部分SOD活性在处理后6、24、72 h与对照相比均无显著差异(图4-B)。根系的POD活性在处理后24 h出现峰值,到72 h时活性又下降;而地上部分的POD活性随着处理后时间延长呈逐渐上升趋势,并且每个时间点与前一时间点相比酶活性均显著提升(图4-D)。根和芽CAT活性变化趋势大致相同,均表现为在辐射后6、24 h与对照相比无显著差异,在72 h时活性显著下降(图4-F)。
2.5 碳离子束和X射线辐射对水稻旗叶遗传物质多态性的影响
本研究共使用了UBC系列的29条引物,其中10条引物被用于数据分析。由表4可知,在高能碳离子束辐射下,10条引物共扩增出95条条带,平均每条引物扩增出9.5条条带,其中25条为多态性条带,占扩增条带总数的29.31%,其中UBC840产生的多态条带百分比最高(83.3%),UBC887和UBC889引物未产生多态条带。由表5可知,在X射线辐射下,10条ISSR引物共扩增出了92条条带,平均每条引物扩增出9.2条条带,其中33条为多态性条带,占扩增条带总数的37.76%,其中UBC890产生的多态条带百分比最高(62.50%),UBC886和UBC889引物产生多态条带百分比最低(20.00%)。表明高能碳离子束辐射和X射线辐射均影响了水稻旗叶的遗传物质多态性,其中经过X射线辐射处理的多态率较高。
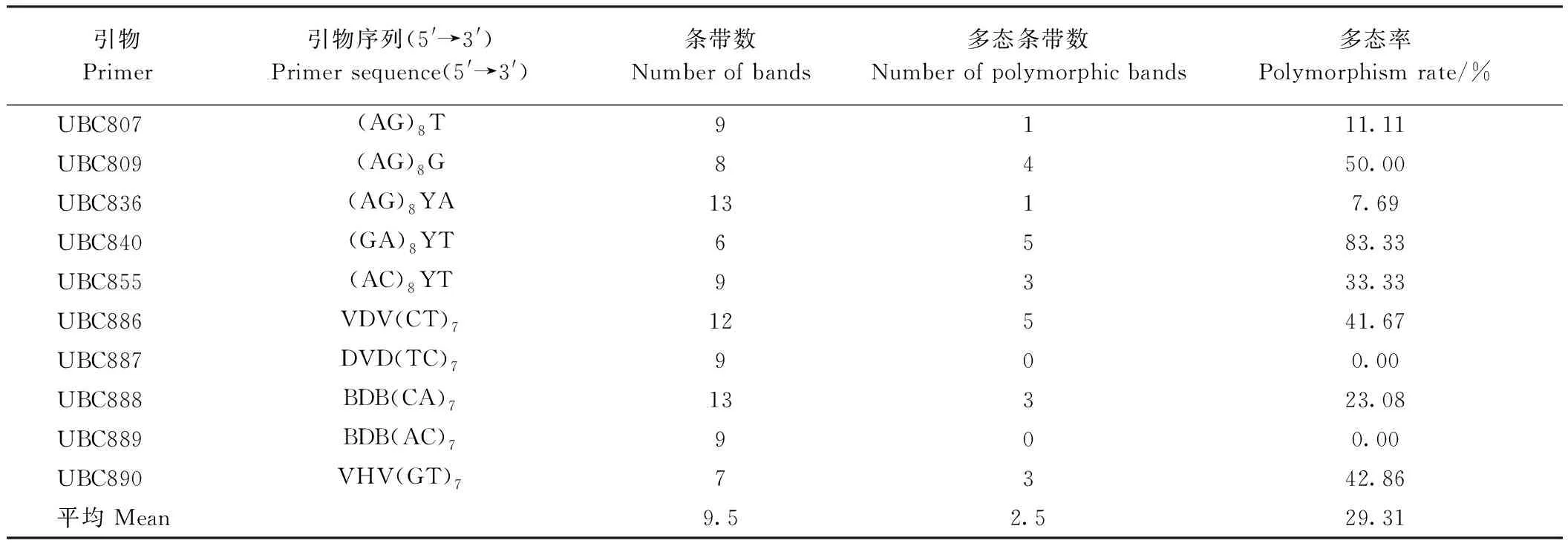
表4 高能碳离子束辐射水稻幼苗ISSR多态率分析Table 4 Analysis of ISSR polymorphism rate of rice seedling irradiated by high-energy carbon ion beam
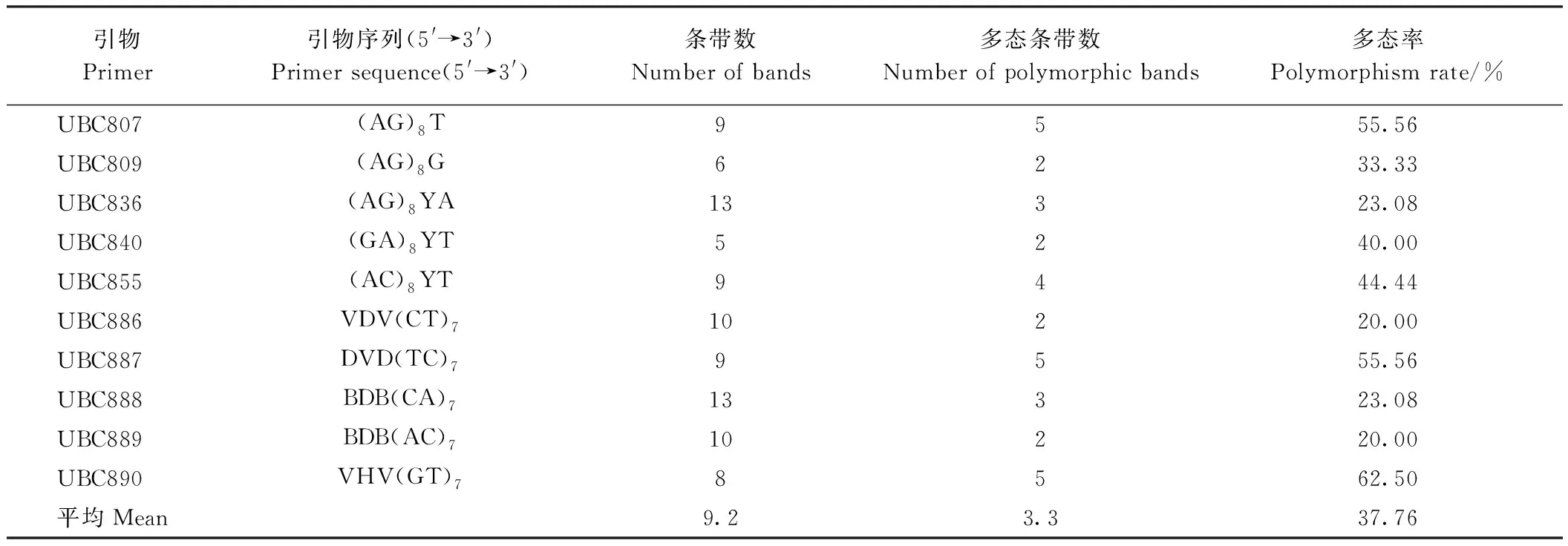
表5 X射线辐射水稻幼苗ISSR多态率分析Table 5 Analysis of ISSR polymorphism rate of rice seedlings irradiated by X-ray
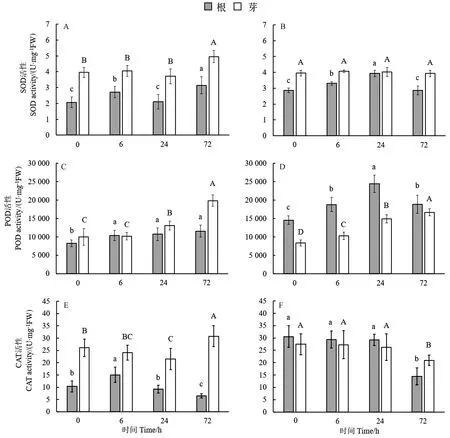
注:不同小写字母表示根部差异显著(P<0.05),不同大写字母表示地上部分差异显著(P<0.05)。Note: Different lowercase letters indicate significant differences in the roots,and different uppercase letters indicate significant differences in the shoot (P<0.05).图4 高能碳离子束(A、C、E)和X射线(B、D、F)辐射对水稻幼苗SOD、POD、CAT活性的影响Fig.4 Effects of high-energy carbon ion beam(A、C、E) and X-ray(B、D、F) radiation on SOD,POD and CAT activity of rice seedlings
3 讨论
辐射作为一种有效的物理诱变手段,能够影响植物生长发育、造成损伤并产生突变。前人研究表明,相比于X射线、γ射线等低LET电离辐射,重离子束辐射植物具有对M1代的生理损伤轻的特点[19]。梅曼彤等[20]研究表明,高LET重离子能更有效地抑制水稻种子萌发、幼苗生长,降低当代植株的结实率,程维民等[21]和杨瑰丽等[22]也证实了这一点。已有研究表明,与γ射线相比,重离子束辐射对水稻干种子的致死率及突变率有所不同[23]。本研究使用两种不同类型的辐射处理水稻幼苗,对当代损伤效应进行研究,结果表明两种辐射处理在较短时期内抑制了幼苗的生长,不论是经高能碳离子束还是X射线辐射处理,随辐射剂量的增加,水稻的根长和苗高整体呈现降低趋势。此外,研究还发现水稻幼苗存在低剂量辐射的刺激效应。由于两种辐射在植物体内能量沉积的方式不同,即LET不同,在造成相同损伤的情况下,对应的辐射剂量不同,表明辐射处理后在根长抑制相似的情况下,X射线的辐射剂量(40 Gy)大约是高能碳离子束辐射剂量(20 Gy)的2倍,即高能碳离子束对水稻幼苗根长抑制的RBE约为2[24]。
前人研究指出,水稻干种子经辐射处理后,株高会随着剂量的增加而下降[25]。而本研究中,辐射剂量的增加并未显著影响存活植株的株高,表明在一定剂量范围内,水稻幼苗经辐射处理后,存活植株的营养生长未受到辐射太大的影响,在表型上无显著变化,Xu等[10]在碳离子束辐射水稻干种子研究中发现了类似的现象。此外,在本研究中,在较高剂量的X射线辐射下,存活植株的分蘖能力、结实率及千粒重都受到了极大的影响,表现为水稻分蘖数显著增加,结实率及千粒重显著降低,这与刘宝海等[26]的研究结果类似。推测是控制分蘖的基因受到的损伤在生长过程中无法完全自我修复或产生了生殖生长紊乱,导致水稻无效分蘖数增加,不断消耗养分,影响有效分蘖的生长,进而使水稻产量受到严重影响并造成M1植物育性下降甚至不育。
光合作用是植物特有的生理功能,其中叶绿素的生物合成受到多种因素的影响[27]。本研究中,40 Gy X射线辐射处理水稻幼苗后,经过一段时间培养,叶绿素含量显著提高,说明40 Gy辐射在一定时间内提高了水稻光合作用。在20 Gy高能重离子束辐射处理水稻幼苗后,其叶绿素含量相比对照并未出现显著性差异。健康植物叶片的Fv/Fm值在0.80~0.85之间波动,X射线处理组的Fv/Fm值较对照虽有显著降低,但仍在正常值的范围内,说明植株健康。Y(Ⅱ)值可作为评价植物胁迫程度的指标[28]。本研究表明,在两种辐射处理中Y(Ⅱ)值都未出现显著变化,说明在辐射后第30天水稻植株可正常生长。
植物体内存在不同类型的抗氧化酶,抗氧化酶活性可以反映植物的抗氧化能力[29]。本研究中,水稻幼苗地上部分与地下部分抗氧化酶活性变化有很大差异,其中地上部分SOD活性在高能碳离子束辐射后72 h时显著提高,可能是因为SOD的合成通路被激活,在辐射处理后72 h时提高了SOD的表达量。地上部分POD活性随着辐射后时间的延长显著提高,植物抗氧化酶活性的提高可缓解胁迫带来的膜过氧化损伤[30]。植物体内过氧化物酶与光合作用、呼吸作用等代谢密切相关,能清除体内的H2O2,维持活性氧代谢平衡[31]、保护膜结构,从而在一定程度上缓解或抵御逆境胁迫。水稻幼苗经两种不同辐射处理后,3种抗氧化酶活性的变化趋势不尽相同,但在抗氧化酶系统的共同作用下,能够有效清除ROS的产生,将水稻体内的自由基维持在正常水平。
ISSR作为一种有效的基因指纹技术,对确定水稻品种间遗传变异和品种快速鉴定有较大价值[32]。本研究表明,高能碳离子束和X射线辐射均能诱发水稻M1植株遗传物质多态性的变化。其中,X射线辐射引起的水稻当代基因组DNA的平均遗传多态性高于碳离子束辐射,认为这与X射线辐射对M1代造成的损伤更为严重有关。冯慧[33]研究表明,经碳离子束辐射获得的M1和M3代拟南芥基因组DNA的平均多态率均高于γ射线,这与本研究中高能碳离子束辐射水稻幼苗M1代遗传多态性较低结果相反,这与诱变操作选取的物种及样品的起始状态均相关。在植物的诱变育种操作中,多数采用种子。种子是由多细胞构成,在受辐射处理后,多数情况下仅是个别细胞发生突变,因此,存活的植株是由带突变基因的细胞和未突变基因的细胞所组成[34]。本研究以水稻幼苗为材料进行辐射诱变操作,其构成的植株以及后代是否存在突变嵌合现象还需要进一步研究。此外,了解不同电离辐射对水稻植株所产生的当代生物学效应,开展辐射诱变机理研究,可为水稻种质创新提供更佳翔实的数据积累。未来将从多角度出发,进一步对水稻不同生长时期及更多农艺学性状展开研究,从而为育种工作者提供更加高效、更加多样化的诱变技术方法。
4 结论
植物对电离辐射的响应是一个复杂的过程,不仅与辐射的类型、剂量有关,也与样品所处的生理状态相关。本研究结果表明高能重离子束和X射线辐射处理水稻Kitaake幼苗后,在具有相同生物学效应下,植物对两种类型电离辐射所产生的生理响应不同。在较短时间内,两种辐射对幼苗的根及芽的抗氧化系统存在显著影响。高能重离子束辐射对水稻的叶绿素含量和Fv/Fm值均无显著影响;X射线辐射可增加水稻的叶绿素含量,降低Fv/Fm值。20 Gy高能碳离子束辐射组水稻幼苗ISSR多态率为29.31%,40 Gy X射线辐射处理水稻幼苗ISSR多态率为37.76%。总体而言,高能碳离子束辐射水稻幼苗所产生的当代损伤效应低于X射线处理,推荐的诱变剂量区间为:高能碳离子束20~30 Gy,X射线30~40 Gy。
