热带生态区玉米自交系雄穗生育特性研究
2022-09-19许海涛冯晓曦许波张军刚郭海斌王友华王成业
许海涛,冯晓曦,许波,张军刚,郭海斌,王友华,王成业
(驻马店市农业科学院/河南玉米产业技术体系驻马店综合试验站,河南 驻马店 463000)
玉米(Zea maysL.)是典型的杂种优势作物,雄穗着生在茎秆顶端,雌雄异位同株,属于异花授粉作物[1]。雄穗是玉米自交系的繁殖器官,同时也是产生花粉的器官,既是光合物质供应与形成的库与源,也是光合物质运输的流[2]。雄穗分枝数与玉米种子的繁殖特性存在相关性,分枝数多少对雄穗大小及花粉量起到决定性作用,分枝过长或者数量过多均可过量消耗营养,后期遮阴严重影响植株下层的通风透光[3]。雄穗主轴长度与玉米茎秆抗折力呈极显著正相关,通过雄穗分枝平均长度对雄穗小花数产生间接影响[4]。雄穗开花时期的长短、每日开花持续时长和散粉量的多少对玉米籽粒产量的形成起决定性作用[5]。
遗传因素和环境因素共同影响玉米雄穗生育进程和分化速率,继而进一步影响到雄穗的开花散粉[3]。有研究指出,雄穗分枝数与产量呈显著负相关,雄穗重量在未追施氮肥条件下与产量也呈显著负相关,小雄穗有益于提高植株下层叶片的透光性,降低营养过量消耗,但从保存玉米自交系种质资源与杂交种子生产方面来看又必须确保雄穗具有充足的小花与花粉量[6]。霍仕平[7]研究发现,成熟期不同玉米品种在相同环境条件下其雄穗的生育分化速率也不一致,雄穗分化早熟型品种比晚熟型要快,雄穗分枝平均长度、雄穗主轴、小花数目及着生密度主要受显性与加性遗传基因支配。也有文献报道,发达的雄穗可对玉米个体后代繁衍提供竞争的能力及稳定的条件[8]。
前人有关玉米雄穗生育分化、开花散粉方面的研究多集中在我国大陆生态区[9-11]。目前我国玉米育种院所、公司、个人在热带生态区的海南三亚、陵水、九所、利国、崖州、梅山、黄流等地进行玉米加代、繁育、制种已成为玉米育种工作常态,而有关该生态区玉米自交系雄穗生育特性方面的研究尚鲜有系统报道。为此,本研究以自选骨干自交系为材料,在位于热带生态区的乐东黎族自治县九所镇研究不同基因型玉米自交系的雄穗主轴、分枝生育特性,雄穗逐日开花、逐时散粉规律,以及雄穗鲜重、雄穗体积与花粉量的相关性,以期明确雄穗的合理结构及热带生态区雄穗生育特性,确切了解雄穗集中开花时期及集中散粉时间,为解决种子生产花期不遇时的人工补救和自交系种质改良提供技术支撑。
1 材料与方法
1.1 试验地概况
试验于2021年在海南省乐东黎族自治县九所镇的驻马店市农业科学院南繁基地(18°26′41″N,108°57′38″E,海拔高度36 m)进行。该地处于海南岛西南部,光照充足,热量充沛,属于典型的热带季风气候。试验期内干旱少雨。供试土壤为砂质壤土,砂质含量为68.91%。
玉米雄穗生育与散粉期气温、相对湿度、日照时数分别见图1、图2和图3。用Elitech RC-4HC温湿度记录仪实测田间气温、相对湿度,人工记录田间日照时数。结果为:期间最高气温19~29℃,最低气温14~20℃,9时气温16~23℃,12时气温19~26℃,15时气温18~29℃,18时气温16~27℃;9时相对湿度62%~97%,12时相对湿度44%~94%,15时相对湿度41%~75%,18时相对湿度61%~78%;日照时数0~10.8 h,平均日照时数7.6 h。

图1 玉米自交系雄穗生育与散粉期气温

图2 玉米自交系雄穗生育与散粉期相对湿度

图3 玉米自交系雄穗生育与散粉期日照时数
1.2 试验材料
供试玉米自交系:由驻马店市农业科学院玉米研究所提供,分别为CL85、17H16、PHHJC、KWS2-4321、13H6、13H37,其中CL85、17H16、KWS2-4321、13H6、13H37为自选骨干自交系。
供试肥料:西洋复合肥(N∶P2O5∶K2O=16∶16∶16,总养分含量≥48%,贵州诺威施生物工程有限公司生产)、尿素(46% N,河北正元氢能科技有限公司生产)。
1.3 试验设计与田间管理
试验采用随机区组设计,重复3次。小区行长6 m,15行区,行距60 cm。小区面积54 m2。南北行向,四周设置2行保护行。
2021年11月14日,按照当地垄作模式旋耕后起垄,伴随起垄集中施肥,增厚耕作层,既利于排水防风蚀,又可提高地温。具体做法为:用牛工先犁开1行沟,集中撒施375 kg/hm2复合肥于沟内作基肥,再从沟两侧向内各翻1犁形成垄,垄面用钉耙平整同时去除杂草。垄高20 cm,垄间距60 cm。11月15日按照株距人工点播于垄面中间,每穴2粒,密度7.5万株/hm2。11月16日喷水,确保足墒出苗。11月21日出苗,11月28日定苗。12月2日追施尿素125 kg/hm2,12月9日追施复合肥500 kg/hm2。玉米生育期间干旱少雨,人工喷灌浇水,其它管理同当地大田生产。
1.4 测定项目与方法
雄穗生育动态:各自交系均于雄穗未抽出用手可摸到时,选取雄穗生长基本一致且无虫害植株3株开始取样,其开始取样至结束取样日期见表1。从植株上剥离雄穗后立即放入干燥洁净自封袋内,之后室内测定雄穗主轴长度(直尺)、主轴直径(分度值0.01 mm,游标卡尺)、雄穗分枝长度(直尺)、分枝直径(分度值0.01 mm,游标卡尺),每隔3 d测定1次。
雄穗逐日开花动态:各处理均选雄穗大小基本一致且无虫害植株3株,自雄穗始花起每天18时调查雄穗主轴小花开花数、分枝小花开花数,当日调查结束后用平头镊子夹掉开过的小花,以免影响到第2天的调查,直至雄穗开花结束为止。各自交系始花调查日期至终花结束日期见表1。
雄穗逐时散粉动态:各处理均于雄穗第3天开花旺盛期选取生长一致且无虫害的植株3株,7时用羊皮纸袋完全套牢雄穗,将纸袋开口处裹紧雄穗主轴、曲别针固定,防止袋内花粉外泄;8时开始每隔1 h收集1次羊皮纸袋内花粉,收集完立即更换新纸袋,以便下次收取花粉。备好网筛(图4a),过筛花粉混合物除去颖片等杂物(图4b),筛后新鲜花粉(图4c)放入洁净干燥自封袋内(图4d),干燥后称重(分度值0.1 mg,SI-234天平,丹佛仪器北京有限公司生产),取样至18时终止。各自交系取样日期见表1。

表1 6个自交系不同处理取样开始日期与结束日期 (年-月-日)

图4 玉米自交系雄穗花粉处理示意图
雄穗鲜重、雄穗体积与花粉量相关性研究:各自交系雄穗开花第3 d时选取雄穗生长一致且无虫害植株3株,7时用羊皮纸袋完全套牢雄穗,将纸袋开口处裹紧雄穗主轴、曲别针固定,防止袋内花粉外泄;18时收取花粉后按照图4处理花粉,干燥后称重。另采同等标准雄穗3个快速放入干燥洁净自封袋内,之后放入冰盒内用于室内测定雄穗鲜重和体积(排水法,雄穗体积V=V2-V1,图5)[12]。各自交系取样日期见表1。

图5 排水法测定玉米自交系雄穗体积示意图
1.5 数据处理与分析
采用Microsoft Excel 2003进行数据处理、作图和雄穗生育动态及其相关变化动态分析。
2 结果与分析
2.1 玉米自交系雄穗主轴生育动态
玉米自交系雄穗是育种过程中必需研究的关键性状之一,较小雄穗可降低养分消耗及提高中下层叶片透光率。由图6可知,玉米自交系雄穗主轴长度、主轴直径随生育进程基本上整体呈现“S”型曲线动态变化。雄穗生育第1~4天主轴长度迅速增加,第4~7天增速变缓,第10~13天达到峰值,之后略有降低。具体看,自交系CL85、17H16、PHHJC雄穗主轴长度随生育推进呈现持续增加趋势,第1~7天增幅较大,第10天后平稳增加,第16天达到峰值,雄穗生育时期明显高于其它自交系;KWS2-4321雄穗主轴长度第1~4天急剧增长后,第4~10天缓慢增加,第13天达最大值,之后又有所降低;13H6、13H37雄穗主轴长度前期生长较快,第10天即达峰值,第13~16天又平稳下降,第16天显著低于CL85、PHHJC。
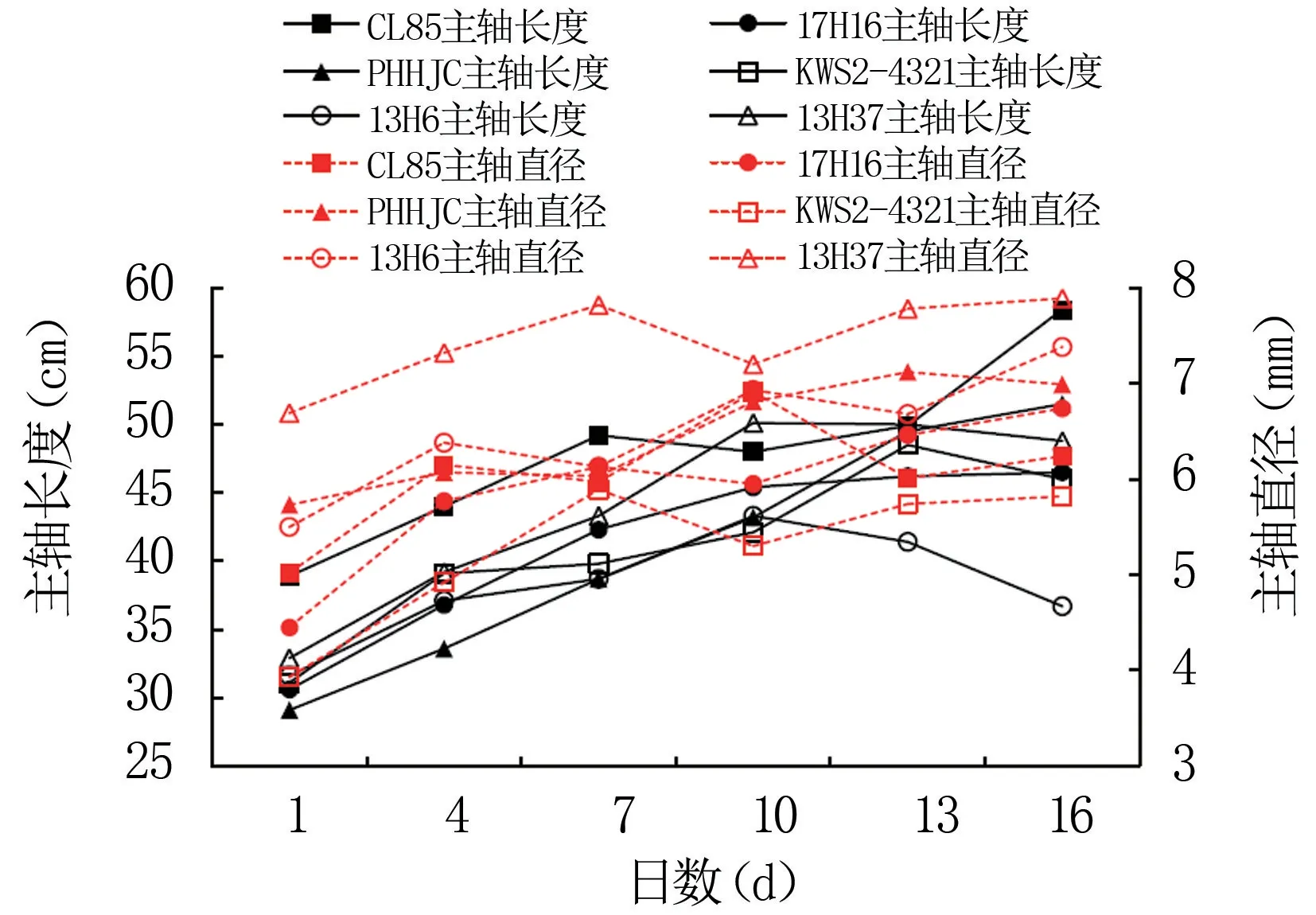
图6 玉米自交系雄穗主轴生育动态
雄穗主轴直径与长度变化不尽一致。CL85、PHHJC、13H6、13H37雄穗主轴直径第1~4天迅速增加,第7天之后变化幅度相近,雄穗主轴粗度形成较快,其中CL85第10天达到峰值,远远快于PHHJC、13H6、13H37,后者第16天形成峰值;17H16、KWS2-4321雄穗主轴直径生育时期大于CL85、PHHJC、13H6、13H37,第1~7天大幅生长后进入缓增期,分别于第16天、第7天达到峰值,17H16雄穗主轴直径生育时期显著高于KWS2-4321。
自交系雄穗主轴增长、直径变粗是雄穗体积增大的重要因子。玉米自交系雄穗主轴长度、主轴直径的生育与生育日数之间符合二次多项式数量关系:y=ax2+bx+c(a、b、c为常数,x为生育日数),其数学模型见表2。

表2 玉米自交系雄穗主轴长度、主轴直径与 生育日数间的数学模型
2.2 玉米自交系雄穗分枝生育动态
由图7可知,整体上看,自交系雄穗分枝长度与分枝直径随生育进程呈现类似“S”型变化动态,其中雄穗分枝长度第1~4天生长缓慢,第4~7天生长较快,第10天之后又平稳变化。自交系CL85雄穗分枝长度随生育进程连续变长,第1~7天长度变化幅度较小,第10天之后迅速增长,第16天达到最大值;17H16雄穗分枝长度经1~7 d缓增期后第10天快速增长,第13天达到峰值后略有降低;PHHJC、KWS2-4321、13H37雄穗分枝长度缓增期较短,为1~4 d,第7天达最大值,之后稍有降低;13H6雄穗分枝长度经7 d缓增期后第10天达到峰值,第13~16天又有所下降。

图7 玉米自交系雄穗分枝生育动态
自交系雄穗分枝直径较主轴直径生育持续时期长。随生育进程,自交系CL85雄穗分枝直径第1~10天持续增粗,第13天略有降低,第16天直径最粗;17H16、PHHJC、KWS2-4321、13H6雄穗分枝直径渐增期明显高于13H37,但低于CL85,雄穗分枝直径随生育进程逐渐增粗,第13天达最大值后有所下降;13H37雄穗分枝直径缓增期较短,第10天即达峰值。
2.3 玉米自交系雄穗逐日开花动态
玉米自交系雄穗开花数的多少与当日散粉量密切相关。雄穗主轴开花先从中上部始,再向上,最后向下依次开放;雄穗分枝上部花先开放,后向下开放[13,14]。由图8可知,玉米自交系雄穗主轴、分枝开花均呈现典型的抛物线动态变化,开花数随雄穗生育进程呈现先升高后下降趋势。不同基因型玉米自交系主轴开花比分枝提前1~2 d,但分枝终花比主轴又延迟1~2 d;雄穗主轴开花持续5~7 d,分枝开花持续5~6 d;主轴盛花期集中在开花第2~3天,占主轴开花总数的63.9%~77.4%,雄穗分枝盛花期多集中在开花第4~5天,占分枝开花总数的61.6%~83.2%。

图8 玉米自交系雄穗逐日开花动态
自交系CL85第1~2天主轴开花较少,均在10朵以下,第3天开花显著增加,第4天又显著下降,第5天之后又降到10朵以下;17H16雄穗主轴开花第2天达到最大值,第5天下降到10朵以下,第6天终止开花,持续开花期远低于CL85、13H6、13H37;PHHJC、KWS2-4321雄穗主轴开花持续期均为6 d,开花数分别于第2天、第3天达到峰值,第5~6天均降到10朵上下;13H6、13H37雄穗主轴开花分别于第3天、第2天达到峰值,第4天均显著下降,第5天降至10朵左右,开花持续期均为7天。
17H16、13H6雄穗分枝开花始于第3天,始花均在40朵以上,远大于CL85、KWS2-4321、13H37始花数,均于第4天达到最大值,两者持续开花期最长为6 d,17H16分枝日开花最大值超其它自交系98.4~168.7朵;CL85、KWS2-4321、13H37雄穗分枝第3天始日开花数量较少,均在10朵以下,第4天达高峰值后又明显下降,第7天显著降至10朵以下;PHHJC雄穗分枝日开花数量第3天始至第6天变化幅度相近,远低于其它自交系,第4天达到顶峰,至第7天下降至2朵以下。
2.4 玉米自交系雄穗逐时散粉动态
玉米自交系雄穗散粉与温度、相对湿度密切相关[1],散粉量随每时刻温度、相对湿度的变化呈现一定的规律性,基本符合其开花规律,不同自交系间不同时刻散粉量存在较大差异。由图9可知,不同自交系雄穗散粉量随时间进程呈先增加后降低趋势:8时由于温度低散粉量较少,为8~18.8 mg;9—10时为雄穗散粉旺盛期,散粉量急剧增加,约占当日散粉总量的44.4%~52.7%;10—11时大幅下降;12时之后散粉量缓慢降低,散粉持续到17—18时。
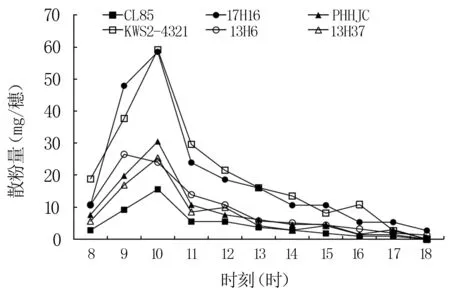
图9 玉米自交系雄穗逐时散粉动态
自交系17H16、13H6当日散粉持续时间最长,18时还分别有1.3~2.7 mg/穗的散粉量,远大于CL85、PHHJC、KWS2-4321和13H37,但二者散粉盛期出现时间不同,分别于10、9时达到最大值;CL85、13H37散粉盛期相对集中,持续时间较短,9—10时散粉量均达当日散粉总量的50%以上,12时后明显下降,其散粉量分别占当日散粉总量的11.3%、12.3%,17时均在当日散粉总量的2%以下,18时终止散粉;PHHJC、KWS2-4321自交系8—10时散粉量逐时升高,10时达到最大值,散粉量分别为30.5、59.2 mg/穗,之后开始下降,12时散粉量分别下降到7.6、21.5 mg/穗,16时散粉量分别为10.8、1.5 mg/穗,且KWS2-4321散粉盛期持续时间明显大于PHHJC,二者散粉持续时间与CL85、13H37一致,但低于17H16、13H6。
2.5 玉米自交系雄穗鲜重、雄穗体积与散粉量的相关变化动态
由图10可知,6个玉米自交系雄穗散粉量与雄穗鲜重、雄穗体积的线性回归分析结果表明,自交系雄穗散粉量与雄穗鲜重、雄穗体积呈显著正相关。6个自交系雄穗散粉量与雄穗鲜重的拟合方程为y=0.0245x+6.4707,决定系数R2=0.7229,达显著水平,回归系数b=0.0245(>0),据此结果得出雄穗鲜重与散粉量呈显著正相关,即雄穗散粉量每增加1 mg,雄穗鲜重需提高0.0245 g;6个自交系雄穗散粉量与雄穗体积拟合方程为y=0.0747x+7.8464,决定系数R2=0.8212,达极显著水平,回归系数b=0.0747(>0),据此结果得出雄穗散粉量与雄穗体积呈显著正相关,即雄穗散粉量每增加1 mg,雄穗体积需提高0.0747 mL。说明较大的雄穗有利于提高其散粉量,利于花丝获得足够的花粉,提高结实率。

图10 玉米自交系雄穗鲜重、雄穗体积与 散粉量的相关变化动态
3 讨论与结论
董海合等[15]研究认为,玉米雄穗主轴长度通过雄穗分枝长度对雄穗小花数产生间接影响,分枝过长或者过多对营养会过度消耗,不利于提高籽粒产量。齐小芳[11]对雄穗发育形态指标的研究发现,雄穗主轴长度、主轴直径随发育过程的推进呈先缓慢增加后快速增长的变化趋势,符合二次多项式数学模型。这与本研究结果基本一致,即雄穗生育第1~4天主轴长度迅速增加,第4~7天增加变缓,第10~13天各自交系分别达到最高峰,峰后略有降低,整体上呈现“S”型曲线动态变化。徐洪文等[9]对不同基因型玉米雄穗主轴长度、主轴直径的相关研究表明,其与生育日数间也表现出二次多项式数学模型关系,不同基因型玉米雄穗主轴长度、主轴直径生育整体趋势相同,这也进一步印证了本研究结果的可靠性,但数学模型与变化幅度之间存在一定差异,与本研究结论也基本吻合。
前人研究指出,玉米雄穗分枝长度、分枝直径、分枝数量和雄穗的大小密切相关,雄穗分枝适宜既可提供足够花粉量、确保结实性,又可降低雄穗过大而对光合物质过多消耗,能向雌穗转移更多的光合物质,是玉米育种指标的一个选育方向[2,16]。本研究表明,适宜的雄穗分枝长度,其小花着生密度大,可确保足够的小花数及花粉量;不同自交系间雄穗分枝长度与分枝直径生育动态虽存在明显差异,但其发育基本上呈现先升后降趋势,与雄穗主轴生育动态类似,即雄穗分枝长度第1~4天生长缓慢,第4~7天生长较快,第10天之后又平稳变化。此结果与前人[9]有关玉米雄穗发育变化动态规律相吻合。
玉米雄穗开花散粉与温度、相对湿度密切相关,温度25~28℃雄穗开花最多,相对湿度70%~90%时雄穗盛花期为开花后第2~5天,第3~4天为开花高峰期,开花持续7~9 d[1]。本研究结果显示:开花数随雄穗生育进程呈先升高后下降趋势;不同基因型玉米自交系雄穗主轴、分枝逐日开花数存在一定差异;主轴比分枝提前1~2 d开花,但分枝终花比主轴延迟1~2 d;雄穗主轴开花持续5~7 d,分枝开花持续5~6 d;雄穗主轴盛花期集中在开花第2~3天,雄穗分枝盛花期多集中开花第4~5天。这与卢宗志[5]、张晓光[13]等的结论相同。本研究还表明,热带生态区雄穗集中开花期延迟1~2 d,与该区冬季雄穗开花时的温度、相对湿度适宜有关(图1、图2),两者远低于大陆生态区夏季玉米雄穗开花时的温度和相对湿度,这有利于雄穗持续开花,而高温干燥使大陆生态区玉米雄穗开花过于集中。
已有文献报道,大陆生态区夏季一天中晴天7—11时为雄穗盛花期,上午雄穗开花最多,12时后雄穗开花渐少,夜间极少开花,阴雨天延迟开花[11,13]。海南热带生态区冬季干旱少雨,光照充足(图3)。本研究表明,不同玉米自交系间不同时刻散粉量存在较大差异,但整体上雄穗散粉量随每时刻温度、相对湿度的变化呈现一定的规律性,基本符合其开花规律,呈先增加后降低趋势:8时由于温度低散粉量较少,为8~18.8 mg/穗;9—10时散粉旺盛,散粉量急剧增加;10—11时大幅下降;12时之后散粉量缓慢降低,散粉持续到17—18时。该散粉旺盛时刻与前人报道[5]不同,前人研究所处大陆生态区雄穗生育处于夏季,温度较高,白天温度上升快,雄穗开花散粉较本研究提前1~2 h,但整体变化动态趋势与本研究结论基本一致。
目前有关玉米自交系雄穗散粉量与雄穗鲜重、雄穗体积的相关关系尚未有文献报道。本研究结果表明,自交系雄穗散粉量与雄穗鲜重、雄穗体积呈显著正相关,符合线性回归分析数学模型,说明较大的雄穗有利于提高其散粉量,利于花丝获得足够花粉,提高结实率,但雄穗过大则会严重遮荫,过度消耗营养。因此,适宜大小的雄穗既能提供足量花粉、确保花丝完全授粉、提高结实性,又可降低光合产物的大量消耗,使其尽可能转入雌穗。
