扬州泰州国际机场鸟类多样性及鸟击防控
2022-09-17陈婉袁思佳钱汝恩张俊胡超超常青
陈婉, 袁思佳, 钱汝恩, 张俊, 胡超超,4,*, 常青,*
扬州泰州国际机场鸟类多样性及鸟击防控
陈婉1, 袁思佳2, 钱汝恩2, 张俊3, 胡超超2,4,*, 常青2,*
1. 江苏开放大学环境生态学院, 南京 210036 2. 南京师范大学生命科学学院, 南京 210046 3. 扬州泰州国际机场, 扬州 225831 4. 南京师范大学分析测试中心, 南京 210046
鸟类群落多样性研究对机场鸟击防控有重要意义。国内机场现今均采用物种丰富度和均匀度等传统物种多样性指标来评价机场周边地区的鸟类群落, 该方法缺乏物种的生态系统功能和系统发育相关信息。为阐明机场鸟类群落多样性特点, 本文以扬州泰州国际机场为研究区域, 进行为期一年的调查, 从物种多样性、功能多样性、谱系多样性三个层次来研究。本研究共记录鸟类71种, 隶属13目33科, 雀形目鸟类物种数最多, 其次为鸻形目。从居留型看, 候鸟较多; 从区系上看, 广布种鸟类最多, 古北界和东洋界鸟类都较少。春季的FRic高于其他季节, 春、夏季的FEve和FDis高于秋冬季。FDiv在季节间无显著性差异, 群落物种的离散程度差异较小, 季节变化对FRic、FEve、FDis具有显著影响。各生境的FRic、FEve、FDiv无显著差异, FDis在湿地与草坪间存在显著性差异。栖息地类型的交错分布支持多种功能性状的鸟类, 使得各生境的鸟类功能分布无显著差异。谱系多样性分析发现, 鸟类群落呈现谱系发散趋势。结合鸟类多样性指数、分布系数及重要值, 严重危险鸟类有6种, 包括斑嘴鸭()、珠颈斑鸠()、夜鹭()、喜鹊()、家燕()、八哥()。结合季节和生境的群落多样性特征, 机场需针对性地开展机场生态环境治理, 以应对危险鸟类在机场区域的活动。
鸟击; 物种多样性; 功能多样性; 谱系多样性
0 前言
鸟击(birdstrike)是指鸟或蝙蝠撞击航空器并造成一定经济损失的情况, 对飞机的安全飞行构成严重威胁[1]。近5年来, 鸟击成为中国民航运输类飞机第一大事故征候类群, 占总事故征候的26.2%。鸟击事件不仅会损坏飞机部件、影响正常航班造成巨大经济损失, 严重时会导致机毁人亡的悲剧[2]。因此, 鸟击防范工作已成为整个航空业高度重视的安全管理工作之一。
近年来, 我国鸟击防范研究取得了一定进展。例如, 对各机场及周边区域的鸟类生物多样性进行调查, 分析其多样性特征, 对飞行安全有威胁的鸟类进行危险等级评估[3]; 通过研究食物(如昆虫、植物)等生境因子与鸟类的关系, 最大限度减少食物来降低鸟类出现的频率[4–5]; 通过对鸟击残留物的鉴定, 可以有效地确定高危鸟种并进行防范[6–7]。目前鸟击防范工作, 以调查鸟类多样性为主, 以研究生境因子和鸟击残留物鉴定为辅。国内机场现今均采用传统的多样性指标(物种丰富度和均匀度等)来衡量鸟类多样性, 只用物种相对丰度的大小表征物种的重要性, 没有提供物种的生态系统功能和系统发育相关性的信息[8]。为了更立体地阐明鸟类群落结构, 很多研究都在传统多样性的基础上引入功能多样性(functional diversity, FD)和谱系多样性(phylogenetic diversity, PD), 以更好地表征群落多样性[9–10]。目前许多机场鸟类群落的调查都未涉及功能多样性和谱系多样性。结合这两项指标, 能提供更多与群落构建有关的信息, 揭示不同生态过程对鸟类群落构成的影响[11], 为机场鸟击防范提供更立体的数据支持。
扬州泰州国际机场(以下简称扬泰机场)位于扬州市江都区丁沟镇境内, 距扬州市区约30 km、距泰州市区约20 km。2018年, 机场全年旅客吞吐量238.42万人次, 不仅是江苏省重要的民用机场, 也是华东地区的干线机场和备降机场之一。经过多年的扩建和发展, 机场及周边区域的生态环境日趋复杂多样, 鸟击防范工作也面临着前所未有的挑战。针对这一现状, 本研究于2019年7月到2020年6月, 对扬泰机场飞行区与机场周边区域鸟类多样性进行调查, 结合物种多样性、功能多样性、谱系多样性对鸟类群落进行系统分析, 并对鸟类的危险性等级进行评估, 为鸟击防范提供更系统的数据支持。
1 研究区域与研究方法
1.1 研究区概况
扬泰机场所处地为冲击平原, 灌溉渠道纵横交错, 周边生境复杂, 具有河流、农田、绿化林带、池塘、草坪等生境。扬泰机场跑道两侧土道面的植被比较单一, 泥土土层较薄, 植被主要为一些单子叶植物, 缺乏大型树木。
1.2 生境划分及样线设置
根据机场飞行区与净空区的鸟类栖息地与活动情况, 将机场及周边生境划分为机场草坪、居民区、湿地、林地、农田五种。在机场内布设1条样线, 机场外东南西北各布设1条样线, 样线长度为5 km, 单侧宽度为50 m。调查时间为2019年7月到2020年6月, 每月调查一次, 各样线均共计12次调查。
1.3 研究方法
1.3.1 调查方法
采用样线法[12]对扬泰机场及周围鸟类进行调查。手持博冠10×42双筒望远镜在日出后3 h内和日落前3 h内以1—2 km·h-1的行走速度观察并记录样线内鸟类的种类、数量以及栖息地类型。调查在天气晴朗、风力不大(一般在3级以下)的条件下进行。根据江苏省气候特点, 本文将2020年3月至5月的数据统计为春季调查的数据, 2020年6月和2019年7、8月统计为夏季的数据, 2019年9月至11月数据统计为秋季调查, 2019年12月和2020年1、2月划为冬季调查数据。
鸟类鉴定、分类地位、居留型划分参考《中国鸟类分类与分布》第三版。
1.3.2 分析方法
物种累积曲线: 在实际的生物群落调查中, 想全面调查很难。物种累积曲线(species accumulation curve, SAC)在物种丰富度估计以及抽样充分性判断方面应用很多, 其模型描述了随着抽样量的加大物种数增加的状况, 是判断调查充分性的有效工具[13]。
鸟类密度: 单位面积范围内鸟类个体数量, 其计算公式为=/(2)(ind·hm-2)[14], 其中为样线内记录的鸟类数量,为该次调查样线长度,为该次调查样线单边宽度(50 m)。
鸟类分布系数:= (/+/)×100%, 式中,与分别表示机场鸟类调查的样线总数与生境总数,与分别表示某种鸟类出现的样线数与生境数。根据分布系数, 将鸟类分为广性分布(100%—200%)、中性分布(50%—100%) 和狭性分布(0%—50%)[15]。
多样性分析:
采用Shannon-Wiener多样性指数进行物种多样性分析, 采用Simpson指数进行优势度分析, 采用Pielou均匀度指数进行均匀度分析, 计算公式分别为
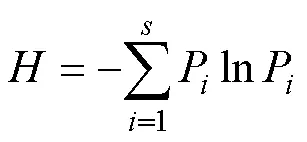


式(1)—(3)中,P为鸟类物种的个体数占所有鸟类物种个体总数的比例。
功能多样性指标包括功能丰富度(functional richness, FRic)、功能均匀度(functional evenness, FEve)、功能离散度(functional divergence, FDiv)和功能分异度(functional dispersion, FDis)。其中, FRic、FEve、FDiv是三个互补的指标[16], FDis是基于功能性状空间距离的指标[17]。这些指标构成了研究功能多样性的框架。FRic表示群落占据的功能空间体积。FEve是描述功能性状空间中物种数分布的均匀性, 先利用性状空间内的各个物种(点)生成总长度的最小路径图, 再分别计算各物种多度在该路径图上分布的均匀度。FDiv表明物种数是如何沿功能性状轴分布的。FDis是先将各物种的数量视为“质量”, 求出群落内物种的“质心”后求得各物种距质心距离均值的偏差。本研究选生物量、食性和取食层作为觅食和栖息生境选择有关的性状, 其中生物量是连续变量, 食性(食软体动物、食鱼、食植和其他食肉)和取食层(水下、水面、地面-水面混合、地面)是分类变量[18]。
群落谱系多样性研究首先根据调查结果建立物种库, 然后从Birdtree (http://birdtree.org)上以Ericson All Species为建树资源下载对应物种的2000棵随机树, 再利用beast2中的TreeAnnotator 2.4.1软件合成合一树。根据合一树, 计算种间平均距离(mean pairwise distance, MPD)和最近种间谱系距离(mean nearest taxon distance, MNTD)。为了确定共生物种在谱系上是聚集还是发散, 使用独立的交换算法随机生成999个物种丰富度和发生频率与所观察到的群落相同的群落(零模型, null model), 并计算零模型MPD和MNTD的平均值。将观测到的MPD和MNTD与该平均值作比较, 计算谱系亲缘关系指数(nearest relative index, NRI)和最近种间亲缘关系指数(nearest taxon index, NTI)。计算公式为:
/=(null-obs)∕null(4)
式中Mobs为或的观测值,null为随机产生的999个或MNTD零模型模拟值的平均值,null为999个随机值的标准差。若和大于0, 表示群落谱系聚集, 反之群落谱系发散[19]。
危险指数: 重要值()=(相对数量成分+相对时间成分+相对空间成分+相对重量成分)/4, 式中, 相对数量成分=(某种鸟的个体数/数量最多的鸟种个体数)×100, 相对时间成分=(某种鸟出现的调查次数/总调查次数)×100, 相对空间成分=(某种鸟出现的样线数/总样线数)×100, 相对重量成分=(某种鸟的体重/体重最重的鸟体重)×100[20]。将调查中观察到鸟类的飞行高度分为0—5、5—30、30—50、50—100和100 m以上5个等级, 鸟类的风险系数分别赋值为0.1、0.5、1.0、0.5、0.1[21]。危险指数=重要值×风险系数。危险指数≥25定为严重危险, 15≤危险指数<25定为很危险, 5≤危险指数<15定为较危险, 危险指数<5定为一般危险[22]。
1.4 数据处理
物种多样性使用vegan包diversity函数[23], 功能多样性使用FD包dbFD函数[24], 谱系多样性使用picante包sesmpd和sesmntd函数[25], 所有指数用gvlma包gvlma函数进行正态性检验, 使用vegan包plot函数绘制物种累计曲线和箱型图。使用multcomp包aov函数对不同指数做单因子方差分析(one-way ANOVA), 若存在显著性差异, 则用Tukey HSD函数做多重比较; 使用t.test函数对NRI和NTI做单组样本均值t检验(One-sample t-test), 判断其是否显著区别于0。数据统计整理用Excel 2016完成。数据处理和绘图在R 4.0.2中完成[26]。
2 结果与分析
2.1 物种组成
本调查共记录鸟类71种, 总计10177只, 分别隶属于13个目, 33个科(表1)。其中雀形目鸟类居多(18科, 37种), 其次是鸻形目鸟类(3科, 13种)。全年观测数量最多的是麻雀(), 其次是喜鹊()、八哥()、云雀()、乌鸫()。根据季节型划分: 留鸟22种, 占总数31%; 旅鸟18种, 占总数25%; 夏候鸟25种, 占总数35%; 冬候鸟6种, 占总数9%。根据区系划分: 广布种63种, 占88.74%; 古北界4种, 占5.63%; 东洋界4种, 占5.63%。
2.2 种群数量
全年鸟类物种数有一定的季节性变化, 在春季秋季出现的最多, 共记有48 种, 占据了调查全部鸟类的67%; 而冬季出现了鸟类物种数的最低值为37种, 占全部种数的51%。从全年的变化来看, 在春季的鸟类物种数有一个明显的峰值(图1)。
从鸟类个体数量的季节分布来看, 全年在秋季月份鸟类数量最多, 而在其余三个季节, 鸟类个体数量稍有减少(图1)。
2.3 物种累积曲线
基于抽样次数的物种累积曲线上升趋势平稳, 其后面逐渐变成一条渐近线(图2), 表明在这一年对机场鸟类的调查抽样量充分。
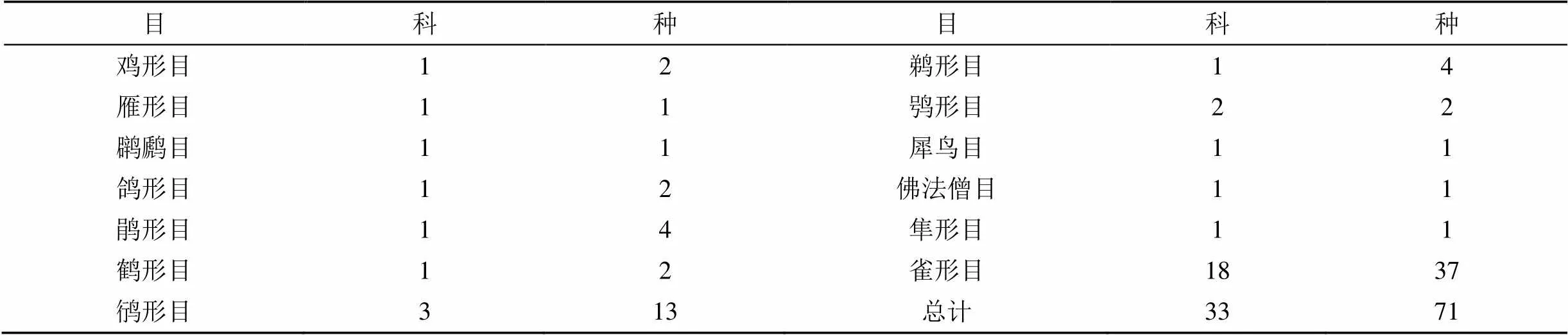
表1 机场及周边鸟类所属分类单元

图1 扬泰机场鸟类物种数以及个体数的季节分布
Figure 1 Seasonal distribution of the number of species and individuals at Yangzhou Taizhou International Airport
Figure 2 Species accumulation curve based on the samples of survey
2.4 群落多样性的季节变化
Shannon-Wiener指数在各季节间存在显著性差异(3, 56=6.53,<0.05; 春a, 夏ab,秋b,冬b; 字母相同表示两者间无显著性差异, 字母不同表示两者存在显著性差异, 下同)。Simpson指数存在显著性差异(3, 56=4.81,<0.05; 春a,夏ab,秋b,冬b)。Pielou均匀性指数存在显著性差异(3, 56=5.98,<0.05; 春a,夏ab,冬b,秋b)。FRic存在显著性差异(3, 56=3.45,<0.05; 春a,夏ab,秋ab,冬b)。FEve存在显著性差异(3, 56=6.08,<0.05; 夏a,春a,秋ab,冬b)。FDiv无显著性差异(3, 56=1.35,=0.28)。FDis存在显著性差异(3, 56=3.04,<0.05; 春a,夏ab,冬b,秋b)(图3)。
2.5 机场鸟类分布状况
本次调查在机场内发现43种鸟类, 以雀形目、鸻形目为主; 在机场外发现56种鸟类, 以雀形目、鸻形目、鹃形目为主。在发现的71种鸟类中, 广性分布的有20种鸟类, 中性分布的有13种鸟类, 狭性分布的有38种鸟类。可见, 扬泰机场以狭性分布的鸟类为主。
不同生境的鸟类分布存在一定差异。不同生境的Shannon-Wiener指数存在显著性差异(4, 39=4.33,<0.05; 林地a,农田a,湿地ab,草坪ab,居民区b)。Simpson指数存在显著性差异(4, 39=3.63,<0.05; 林地a,农田a,湿地ab,草坪ab,居民区b)。Pielou均匀性指数无显著性差异(4, 39=1.67,=0.18)。FRic(4, 39=0.90,=0.48)、FEve(4, 39=0.87,=0.49)、FDiv(4, 39=2.29,=0.08)无显著性差异。FDis存在显著性差异(4, 39=2.80,<0.05; 湿地a,农田a,林地ab,居民区ab,草坪b)(图4)。
2.6 机场鸟类谱系多样性
总体上扬泰机场NRI显著大于0(= 10.94,= 59,< 0.001), 呈现为谱系聚集, NTI显著大于0(= 6.00,= 59,< 0.001), 呈现为谱系聚集。
2.7 鸟类危险性分析
扬泰机场严重危险的鸟类有斑嘴鸭()、珠颈斑鸠()、夜鹭()、喜鹊、家燕()、八哥6种; 很危险的鸟类有白鹭()、池鹭()、灰椋鸟()等8种; 较危险的鸟类有凤头麦鸡()、牛背鹭()、灰喜鹊()等17种; 一般危险的鸟类有东方鸻()、小杓鹬()、小鸦鹃()等40种。
3 讨论
在为期一年调研中记录71种鸟类, 以雀形目和鸻形目鸟类为主, 麻雀和喜鹊等留鸟数量最多, 这与沈阳桃仙机场[22]、重庆江北机场[27]的研究一致。机场周边林地里面常见留鸟集群活动, 且林地上层有大量的喜鹊巢穴。扬泰机场位于中国候鸟东部迁徙路线上, 候鸟和旅鸟的数目较多。扬泰机场广布种的占比很高, 这与机场生境异质化有很大关系[28]。机场及周边地区富含草地、林地、湿地、农田等多种生境, 且林下又有不同程度的灌木, 栖息地的多样化容易吸引广布种鸟类在机场周边筑巢觅食[29]。
全年鸟类物种数和个体数量有一定的季节性变化, 在春季和秋季最多, 这与春秋迁徙季候鸟和旅鸟的活动有很大关系。进入越冬期后鸟类活动相对稳定, 记录的鸟类物种数和个体数量都较低。春夏季由于经常能见到麻雀、喜鹊等留鸟的集群, 因此均匀度高于秋冬季, 导致春夏季物种多样性高于秋冬季[30]。
春季的FRic明显高于其他季节, 表明春季群落在功能空间上占据的体积较大[16]。春夏季的FEve和FDis高于秋冬季。FEve体现群落内物种对有效资源的利用效率[31], FDis描述物种功能和性状空间中物种组(若干物种聚集在一起)所处位置的差异程度[17]。一方面, 春夏两季植被成熟, 食物资源增加, 能够提供大量的捕食生态位, 鸟类物种多样性较高, 在多维生态位空间内的物种分布较秋冬季广[32]。另一方面, 春季候鸟迁徙期和夏季留鸟和候鸟的繁殖消耗大量能量, 需捕食以补充能量, 扬泰机场栖息地食物资源对其繁殖或迁徙有着直接作用[33], 因此春夏季鸟类群落利用有效资源的效率较秋冬季高, 在性状空间内所处位置的差异也高于秋冬季。FDiv为群落物种在功能空间内的离散程度。各季节的FDiv无显著性差异, 表明各季节群落物种的离散程度差距不大, 不同季节物种及物种数的改变影响FRic、FEve、FDis的变化, 导致春夏两季的功能多样性高于秋冬季。

图3 扬泰机场鸟类群落各季节的多样性指标
Figure 3 Biodiversity metrics of bird communities in different seasons at Yangzhou Taizhou International Airport

图4 扬泰机场各生境鸟类群落的多样性指标
Figure 4 Biodiversity metrics of bird communities in different habitats at Yangzhou Taizhou International Airport
机场周围不同生境的鸟类多样性有很大区别。林地、农田、湿地生境的物种数高于机场内草坪和居民区。林地和湿地生境可为留鸟和候鸟提供隐蔽场所和食物资源, 而农田适合麻雀等众多以采食农作物的留鸟生存[34], 内部的水域又为鹭科、鸻鹬类等候鸟提供觅食和栖息场所[35], 因此这几种生境的多样性指标较高。居民区内人为干扰大, 吸引鸟类的主要是人工绿化带, 其鸟种多为麻雀、八哥等人类伴生鸟种, 因此多样性较低。机场内草坪由于人工打理, 草本植物保持在合适的高度, 鸟类难以获得食物资源和隐蔽场所, 因此物种和功能多样性较低[36]。各生境的FRic、FEve、FDiv无显著差异, 仅湿地与草坪的FDis存在显著性差异。虽然扬泰机场及其周边地区的生境异质性高, 栖息地类型多样化, 但各栖息地的分布较为均匀, 生境交错分布, 可以支持多种功能性状的鸟类, 因此各生境的功能多样性指标无明显差异。
综合鸟类的体重、数量、遇见频率及生境利用率等因素对飞机安全飞行的影响, 对鸟类危险等级进行划分。机场内6种严重危险鸟类3种为留鸟, 3种为候鸟。留鸟全年在机场出没, 喜鹊活动范围广且经常集群在机场里觅食, 珠颈斑鸠和八哥由于食性广容易进入机场活动。斑嘴鸭和夜鹭体型较大, 且在春秋迁徙季活动频繁, 对飞行安全危险较大, 需重点防范。在很危险和较危险鸟类中, 鹭科和鸻鹬类鸟类等候鸟占多数, 需在迁徙期防范。
扬泰机场NRI和NTI显著大于0, 表明扬泰机场的鸟类群落谱系呈聚集趋势, 生境过滤主导鸟类群落的构建。这表明生境对于扬泰机场鸟类群落的构建有着重要作用。因此需及时排除生境内部影响鸟类活动的因素, 以防范鸟击事件的发生。
基于研究结果, 对扬泰机场鸟击防范提出以下建议: (1)整治机场草坪。机场内, 草地生态环境是主要的食物链源头, 成为机场内吸引鸟类的主要因素。可以通过控制植被的高度及群落结构的方法来有效切断鸟类、昆虫以及小型啮齿类动物的食物来源和隐蔽场所, 从而降低机场对鸟类的吸引。(2)在迁徙和繁殖季节, 关注鸟类在机场周边的觅食、迁徙等活动, 防止鸟类撞击飞机。(3)防止雨后形成临时的明水区, 及时去除周围水塘的挺水植物, 不给水鸟提供隐蔽的场所, 降低水鸟在其中活动的频率。
致谢:感谢国家科技资源共享服务平台-国家地球系统科学数据中心-长江三角洲分中心(http://nnu. geodata.cn:8008)提供数据支撑。感谢扬州泰州国际机场对鸟类调查提供帮助。
[1] MCKEE J, SHAW P, DEKKER A, et al. Problematic Wildlife[M]. Switzerland, Springer International Publishing, 2016.
[2] JEFFERY R F, BUSCHKE F T. Urbanization around an airfield alters bird community composition, but not the hazard of bird-aircraft collision[J]. Environmental Conservation, 2018, 46(2): 1–8.
[3] 廖峻涛, 吕鸿, 张峰, 等.昆明长水国际机场飞行区鸟类飞行高度及鸟撞风险评估[J]. 云南大学学报(自然科学版), 2018, 40(1): 192–200.
[4] 李永民, 李东伟, 聂传朋, 等.阜阳机场草丛动物群落特征与食虫鸟类相关性[J].生态学杂志, 2017, 36(3): 734–739.
[5] 徐洁, 高晋, 刘进伟, 等.南京禄口国际机场昆虫月际发生规律及其与鸟类关系[J].生态学杂志, 2018, 37(7): 2045–2050.
[6] Yang Rong, Wu Xiaobin, Yan Peng, et al. Using DNA barcodes to identify a bird involved in a bird strike at a Chinese airport[J]. Molecular Biology Reports, 2010, 37(7): 3517–3523.
[7] 高云. 武汉天河国际机场鸟击特点分析及防范对策[J]. 野生动物学报, 2018, 39(4): 820–826.
[8] TSCHARNTKE T, TYLIANAKIS J M, RAND T A, et al. Landscape moderation of biodiversity patterns and processes eight hypotheses[J]. Biological Reviews, 2012, 87: 661–685.
[9] LI Chunlin, ZHANG Yong, ZHA Daode, et al. Assembly processes of waterbird communities across subsidence wetlands in China: A functional and phylogenetic approach[J]. Diversity and Distributions, 2019, 16(42): 1–12.
[10] 丁志锋, 梁健超, 冯永军, 等. 澳门城市栖息地斑块中鸟类群落功能和谱系多样性[J]. 生态学杂志, 2020, 39(4): 1238–1247.
[11] 张敏, 车先丽, 彭逸生, 等. 广东雷州湾和汕头沿海湿地越冬水鸟多样性[J]. 生态与农村环境学报, 2020, 36(5): 553–559.
[12] 许龙, 张正旺, 丁长青.样线法在鸟类数量调查中的运用[J].生态学杂志, 2003, 22(5): 127–130.
[13] 李巧. 物种累积曲线及其应用[J].应用昆虫学报, 2011, 48(6): 1882–1888.
[14] 高玮, 杨志杰. 人工落叶松林中冬季鸟类混合群的相互关系研究[J].动物学杂志, 1991, 26(4): 9–12.
[15] 侯建华, 武明录, 胡永富, 等.森林/草原过渡地带鸟类群落空间结构研究[J].动物学报, 2001, 47(S1): 148–156.
[16] VILLÉGER S, MASON N W H, MOUILLOT D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology[J]. Ecology, 200889(8): 2290–2301.
[17] LALIBERTÉ E, LEGENDRE P. A distance-based framework for measuring functional diversity from multiple traits[J]. Ecology, 2010, 91(1): 299–305.
[18] WILMAN H, BELMAKER J, SIMPSON J, et al. EltonTraits 1.0: Species-level foraging attributes of the world's birds and mammals[J]. Ecology, 2014, 95(7): 2027.
[19] SUKUMARAN J, HOLDER M T. DendroPy: A Python library for phylogenetic computing[J]. Bioinformatics, 2010, 26(12): 1569–1571.
[20] 栾晓峰, 胡思军, 徐宏发.上海农耕区鸟类群落特征及与几种生境因子的关系[J].动物学研究, 2004, 25(1): 20–26.
[21] 王丽君, 周立志, 万政云, 等.骆岗机场鸟类多样性及鸟击风险分析[J].野生动物, 2010, 31(3): 127–130.
[22] 丁振军, 李东来, 万冬梅, 等. 沈阳桃仙国际机场鸟类多样性及鸟撞防范[J]. 生态学杂志, 2015, 34(9): 2561–2569.
[23] OKSANEN J, BLANCHET FG, FRIENDLY M, et al. vegan: Community Ecology Package. R package Version 2.5-6[CP/OL]. R Core Team, 2019 [2020-11-05]. https://CRAN.R-project.org/package=vegan/.
[24] LALIBERTÉ E, LEGEBDRE P, SHIPLEY B. FD: Measuring functional diversity (FD) from multiple traits, and other tools for functional ecology. R package Version 1.0-12[CP/OL]. R Core Team, 2014 [2020-11-05]. https://CRAN.R-project.org/package=FD/.
[25] KEMBEL S W, ACKERLY D D, BLOMBERG S P et al. picante: Integrating Phylogenies and Ecology. R package Version 1.8.2[CP/OL]. R Core Team, 2020 [2020-11-05]. https://CRAN.R-project.org/package=picante/.
[26] R CORE TEAM. R: A language and environment for statistical computing. R Foundation for Statistical Computing [CP/OL]. Beijing, China, 2020 [2020-06-30]. https://www.r-project.org/.
[27] 吴雪, 杜杰, 李晓娟,等. 重庆江北机场鸟类群落结构及鸟击防范[J]. 生态学杂志, 2015, 34(7): 2015–2024.
[28] FISCHER C, FLOHRE A, CLEMENT LW, et al. Mixed effects of landscape structure and farming practice on bird diversity[J]. Agriculture Ecosystems & Environment, 2011, 141(1): 119–125.
[29] SCHÜTZ C, SCHULZE C H. Functional diversity of urban bird communities: effects of landscape composition, green space area and vegetation cover[J]. Ecology and Evolution, 2015, 5(22): 5230–5239.
[30] 柳毅, 徐焕然, 袁红, 等. 天津滨海国际机场鸟类群落结构及多样性特征[J]. 生态学杂志, 2017, 36(3): 740–746.
[31] 路兴慧, 臧润国, 丁易, 等. 抚育措施对热带次生林群落植物功能性状和功能多样性的影响[J]. 生物多样性, 2015, 23(1): 79–88.
[32] ZENG Qing, REID J, SAINTILAN N, et al. Contrasting diversity patterns of breeding anatidae in the northern and southern hemispheres[J]. Ecology and Evolution, 2019, 9(17): 9990–10003.
[33] 王勇, 许洁, 杨刚, 等. 城市公共绿地常见木本植物组成对鸟类群落的影响[J]. 生物多样性, 2014, 22(2): 196–207.
[34] 韦定菊, 赵俊松, 李波, 等. 荥经县农耕区鸟类群落多样性[J]. 四川农业大学学报, 2016, 34(4): 499–504.
[35] 张芳, 王淼, 钟稚昉, 等. 盐城自然保护区典型农田不同作物期的鸟类群落及其对生境的利用[J]. 生态科学, 2019, 38(2): 36–44.
[36] CONKLING T J, BELANT J L, DEVAULT T L, et al. Impacts of biomass production at civil airports on grassland bird conservation and aviation strike risk[J]. Ecological Applications, 2018, 28(5):1168–1181.
Avian diversity and bird strike avoidance at Yangzhou Taizhou International Airport
CHEN Wan1, YUAN Sijia2, QIAN Ruen2, ZHANG Jun3, HU Chaochao2,4,*, CHANG Qing2,*
1.College of Environment and Ecology, Jiangsu Open University (The City Vocational College of Jiangsu), Nanjing 210036, China2. School of Life Science, Nanjing Normal University, Nanjing 210023, China 3. Yangzhou Taizhou International Airport, Dinggou, Yangzhou225831, China 4. Analytical and Testing Center, Nanjing Normal University, Nanjing 210023, China
Investigating the diversity of bird community surrounding the airport plays an important role in bird strike avoidance. Traditional taxonomic diversity is used in Chinese airports to evaluate bird communities, however, this cannot provide information on the ecosystem functions and phylogeny of the species. In order to investigate the diversity of bird communities, we conducted a one-year survey at Yangzhou Taizhou International Airport and studied it from three aspects: taxonomic, functional and phylogenetic diversity. A total of 71 species were recorded, belonging to 13 orders and 33 families. Most of the birds were widely-distributed species, and migrant birds accounted for a relatively high proportion in the community. FRic in spring was higher than in other seasons, and FEve and FDis in spring and summer were higher than in autumn and winter. There was no significant difference in FDiv in each season. The variation of FRic, FEve and FDis was influenced by seasonal species and the number of species. There were no significant differences in FRic, FEve, and FDiv in each habitat, only the FDis of wetland and grassland were significantly different. The intersperse distribution of multiple habitats supported birds with multiple functional traits, so that the functional distribution of birds in each habitat had no significant difference. Phylogenetic diversity analysis revealed that the bird community in the airport showed a trend of overdispersion. Furthermore, the bird strike risk index for each bird species was calculated by multiplying the importance value and risk index. Severe risk birds included,,,,,. Combining the community diversity characteristics of different seasons and habitats, the airport needs to carry out targeted environmental management to deal with the activities of dangerous birds in the airport area.
bird strike; taxonomic diversity; functional diversity; phylogenetic diversity
10.14108/j.cnki.1008-8873.2022.06.023
Q145
A
1008-8873(2022)06-193-09
2020-10-25;
2020-11-29
江苏省自然科学基金项目(BK20181076); 江苏省高校青蓝工程项目(2019SZJS-003)
陈婉(1987—), 女, 江苏开放大学环境生态学院, E-mail: 544934275 @qq.com
通信作者:胡超超 (1986—), 男, 高级实验师, E-mail: huweichen@126.com; 常青 (1966—), 男, 教授, E-mail: changq@njnu.edu.cn
陈婉, 袁思佳, 钱汝恩, 等. 扬州泰州国际机场鸟类多样性及鸟击防控[J]. 生态科学, 2022, 41(6): 193–201.
CHEN Wan, YUAN Sijia, QIAN Ruen, et al. Avian diversity and bird strike avoidance at Yangzhou Taizhou International Airport[J]. Ecological Science, 2022, 41(6): 193–201.
