鹞落坪保护区四种不同生境的夏季鸟类多样性差异
2022-09-17赵彬彬徐文玖李春林
赵彬彬, 徐文玖, 李春林
鹞落坪保护区四种不同生境的夏季鸟类多样性差异
赵彬彬, 徐文玖, 李春林*
安徽大学资源与环境工程学院, 安徽 30601
研究不同生境中鸟类群落的差异可以为其管理与保护提供科学依据。于2017年7—8月在鹞落坪国家级自然保护区的不同海拔区域内开展夏季森林鸟类群落调查, 并分析鸟类群落在森林、公路、村村通和村庄等生境之间的差异。结果共调查到78种4225只鸟, 隶属于9目30科。由于较高的郁闭度和较单一的植被类型, 森林生境中的鸟类物种数、个体数和多样性指数均较低, 与穿越森林的公路无显著差异。尽管村村通和公路生境的鸟类个体数与多样性指数没有显著差异, 但村村通生境中鸟类物种数显著高于公路生境, 仅次于村庄, 这可能与其较高的开阔度和较低的干扰有关。村庄鸟类物种数、个体数和香农威纳多样性指数均显著高于其他生境, 这可能与村庄比较开阔的生境和丰富的食物资源有关。研究结果表明, 在森林景观中, 村落等干扰较大的生境中夏季鸟类群落物种多样性较高。
鸟类群落; 鹞落坪; 物种多样性; 道路; 村庄
0 前言
生物多样性是生态系统稳定的基础, 鸟类是生物多样性的重要组成部分, 发挥着重要的生态系统功能[1]。由于人类对自然资源的过度利用, 自然景观和环境发生了剧烈的变化, 生物多样性因此受到了强烈的影响, 其中, 鸟类是受到影响最为显著的类群之一[2]。我国亚热带和北温带的森林生态系统长期受人类活动的影响, 其自然景观和环境已发生显著改变[3]。研究该地区不同生境中鸟类群落的差异, 可以为鸟类物种多样性的管理与保护提供科学依据。
人类活动对鸟类的影响体现在多个方面。总体而言, 一方面, 人类活动可以直接导致鸟类种群的下降, 体现为对鸟类的捕杀、人为活动和设施对鸟类造成直接的伤害、环境污染提高鸟类的死亡率等[4-5]。另一方面, 人类活动也可能造成鸟类栖息地的退化与丧失、改变鸟类与其他物种和环境的相互关系、引发全球气候变化等, 从而构成对鸟类分布和物种多样性的间接影响[6-7]。人类活动因干扰类型和强度的不同而对鸟类产生不同的影响, 鸟类对人为干扰的响应也因物种而异[8-9]。
道路和人口聚居区建设是人类改造自然景观最常见的两种类型, 在全球范围内对自然景观和生物多样性产生了深远的影响[10-11]。有研究表明, 道路建设对大多数陆生脊椎动物(包括鸟类)的栖息地选择、存活率、繁殖率和扩散能力产生了显著的影响[12–15]。村镇和城市等人口聚居区对鸟类的影响主要体现为直接的干扰和土地利用变化带来的间接影响[16]。道路和人口聚居区对鸟类的影响因其类型和干扰强度的不同而异, 不同物种的响应对策及结果也有所差异[16-17]。我国亚热带和北温带的森林景观中存在密集的道路和村庄, 对生物多样性可能存在广泛的影响, 亟待深入研究。
鹞落坪国家级自然保护区是森林生态系统类型的自然保护区, 其主要保护对象是大别山北亚热带山地森林生态系统及其丰富的生物多样性[18]。由于历史原因和当地社会经济的发展, 保护区内存在很多自然村落和省道、县道及村村通等各种等级的道路[19]。然而, 由于没有相关研究, 这些道路和村庄对保护区内森林鸟类群落的影响仍不得而知。我们在鹞落坪保护区的不同海拔区域内开展森林鸟类调查, 分析森林、村村通、公路和村庄等不同生境中夏季鸟类群落物种组成和多样性的差异, 以期为森林生态系统类型自然保护区中鸟类物种多样性的管理和保护提供科学依据。
1 材料与方法
1.1 研究区域概况
鹞落坪国家级自然保护区(116°02′—116°11′ E, 30°57′—31°06′ N)位于安徽省岳西县, 总面积123 km2, 主要保护对象为大别山区典型代表性的森林生态系统[18]。鹞落坪保护区地处大别山腹地, 受北亚热带季风气候影响, 低海拔区域(<1000 m)年平均气温为13 ℃—14 ℃, 年降水量为1400—1600 mm; 高海拔区域(>1000 m)年均气温为11 ℃—12 ℃, 年降水量为1600—1800 mm。
鹞落坪保护区内植被属北亚热带落叶-常绿阔叶混交林, 植物区系复杂、植物种类丰富、区系起源古老、特有现象明显, 共记录有维管植物182科826属2099种。动物地理区划位于古北区黄淮平原亚区的南限和东洋区东部丘陵平原亚区的北限, 是中国东部地区东洋界和古北界过渡带始端, 共记录有动物58目326科1936种, 其中脊椎动物有31目84科187属286种[19]。
1.2 研究方法
1.2.1 样线设置
在鹞落坪自然保护区内布设24条鸟类固定调查样线, 每条样线长度为1 km。根据生境类型的不同, 将调查样线分为森林样线、村村通样线、村庄样线和公路样线。每种类型的调查样线分别为6条, 高海拔区域(>1000 m)和低海拔区域(<500 m)各3条。在正式调查开始前, 对每一条样线开展预调查, 以熟悉样线周围的环境, 了解各样线中人为干扰类型及强度, 并最终确定样线的布设。村村通样线沿4—5 m宽的水泥路布设, 样线两侧为森林植被, 人为干扰主要是行人和小型交通工具, 鲜有汽车等大型交通工具。森林样线位于林相保存较为完好的森林内部, 沿宽度小于1 m的土路布设, 鲜有人为干扰。村庄样线位于村庄内部, 植被类型多样, 生境开阔, 人为干扰主要为当地居民生活。公路样线沿宽度大于10 m的省道或县道布设, 两侧均为森林植被, 主要干扰为汽车等大型交通工具。
1.2.2 鸟类调查
2017年7—8月, 在晴朗无风的天气里, 沿24条固定样线开展夏季鸟类群落调查。调查从日出后半小时开始, 持续三个小时, 即6:00—9:00。每天调查三条样线, 全部调查完后再开始新一轮调查。每条样线调查三次, 同一条样线的调查次序在不同日期中均不相同。样线调查由两名(同一组)经验丰富的鸟类调查队员完成。正式调查时, 调查队员沿调查样线以1—2 km·h-1的速度行进, 使用双筒望远镜(10×42 WB Swarovski)观察并记录样线两侧30 m范围内的鸟类物种和个体数, 而不记录样线两侧30 m外的及正在飞行的鸟类。鸟类的物种名和分类依据《中国鸟类分类与分布名录》第二版[20]。
1.2.3 数据分析
在每条样线历次调查数据的基础上, 计算物种丰富度、物种多样性指数和均匀度指数。鸟类物种丰富度指数直接使用物种数; 物种多样性指数使用Shannon-Wiener指数(), 其计算公式为:

式中为物种的个体数占各样线记录鸟类个体总数的百分比。
物种均匀度指数采用Pielou指数(), 其计算公式为:
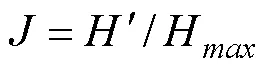
式中,H= ln,为各样线记录物种数。
利用Shapiro-Wilk检验分析4个因变量(即鸟类个体数、物种丰富度、香农威纳指数和均匀度指数)的正态性, 并对其进行对数转换, 使其近似服从正态分布。采用双因素方差分析检验生境类型和海拔对转换后的因变量的影响, 以及生境类型和海拔的交互效应。
采用Berger-Parker优势度指数确定每条样线的优势种, 其计算公式为:

式中N为物种个体数,为各样线记录到的鸟类个体总数。P>10%的物种为优势种, 1%≤P<10%的物种为常见种。
采用主坐标分析(PCoA)分别对高、低海拔区域四类生境类型中的鸟类物种组成的差异性进行检验。与其他间接梯度分析相比, 主坐标分析的优势是在1—3个主坐标轴上可以整合更多的信息用于排序[21]。使用R包的函数执行主坐标分析, 置换次数为999次, 群落间距离指数采用Bray- Curtis相异性指数, 该距离的计算使用R包的函数。所有数据分析均在R 3.6.3中完成。
2 结果与分析
2.1 种类组成
在三次夏季调查中, 总共记录到78种4225只鸟(表1), 隶属于9目30科, 其中雀形目鸟类最多, 为55种4009只, 占总物种数的70.5%, 占总个体数的97.9%。在记录到的物种中, 留鸟有54种, 占总物种数的69.2%, 夏候鸟有16种, 占20.5%, 冬候鸟有6种, 占7.7%, 旅鸟有2种, 占2.6%; 东洋型42种, 占53.8%, 古北型20种, 占25.6%, 广布型16种, 占20.5%。被列入IUCN红色名录易危物种的有3种, 分别为白冠长尾雉()、白喉林鹟()和白颈鸦(); 被列为国家II级重点保护野生动物的有7种, 分别为白冠长尾雉()、勺鸡()、小鸦鹃()、白尾鹞()、赤腹鹰()、黑冠鹃隼()和普通鵟()。
2.2 不同生境中鸟类群落物种多样性的差异
鹞落坪自然保护区夏季鸟类个体数受生境类型和海拔的共同影响, 生境类型与海拔之间不存在显著的交互效应(表2)。在干扰强度不同的生境类型中, 村庄鸟类个体数显著多于其他生境, 而村村通、公路和森林的鸟类个体数不存在显著的差异; 低海拔区域鸟类个体数显著多于高海拔区域(图1)。夏季鸟类物种数受生境类型、海拔及二者互作效应的共同影响(表2)。村庄鸟类物种数最多, 其次为村村通, 而公路和森林鸟类物种数最少; 低海拔区域鸟类物种数显著高于高海拔区域。生境类型和海拔间的互作效应表明, 海拔对鸟类物种数的影响只在村庄和村村通两类生境类型中得以体现, 而在公路和森林中不显著(图1)。
夏季鸟类物种香农威纳多样性指数在不同生境类型和海拔间存在显著差异, 但生境类型和海拔间不存在显著的交互效应(表2)。村庄鸟类物种多样性显著高于其他类型的生境, 而村村通、公路和森林的鸟类物种多样性间不存在显著的差异(图1)。不同类型的生境间物种均匀度指数不存在显著的差异, 但低海拔区域的物种均匀度指数显著高于高海拔区域(表2, 图1)。

表1 鹞落坪国家级自然保护区夏季鸟类群落物种组成

表2 生境类型和海拔对鹞落坪国家级自然保护区夏季鸟类个体数和物种多样性指数的影响

图1 鹞落坪国家级自然保护区各类型生境夏季鸟类个体数和物种多样性指数
Figure 1 Summer bird abundance and species diversity in different types of habitats in the Yaoluoping National Nature Reserve
2.3 不同生境中夏季鸟类物种组成的差异
四类生境中夏季鸟类群落的优势种和常见种如表3所示。主坐标分析(PCoA)表明, 无论是在高海拔(= 0.521,= 0.001), 还是在低海拔(= 0.514,= 0.001), 四种生境中鸟类群落间均存在显著的种类组成差异(图2)。在高海拔区域, 村庄和村村通鸟类群落物种组成差异显著, 而在低海拔区域, 这两种生境的鸟类组成有较强的相似性。无论是在高海拔区域, 还是在低海拔区域, 公路和森林夏季鸟类组成都没有差异, 而这两种生境的鸟类组成与其他两种生境的鸟类组成存在显著差异, 尽管在高海拔区域村村通与公路鸟类组成重叠较多。

表3 鹞落坪国家级自然保护区各类型生境中夏季鸟类优势种(*)和常见种(根据Berger-Parker优势度指数)

图2 鹞落坪国家级自然保护区各类型生境中夏季鸟类群落主坐标分析结果
Figure 2 Results of principal coordinates analysis (PCoA) for the summer bird communities in different types of habitats in the Yaoluoping National Nature Reserve
3 讨论
尽管森林植被给鸟类提供了赖以生存的栖息地, 但植被郁闭度过高反而会降低鸟类的多样性[22]。经过近30年的退耕还林、严禁乱砍滥伐等保护措施的实施, 鹞落坪保护区内的林地面积显著增加, 加之植被的次生演替, 保护区内森林植被的郁闭度显著提高, 植被类型也变得单一[23], 这可能是森林生境中夏季鸟类多样性较低的主要原因。骆鹰等人的研究结果也表明林地鸟类的多样性较低, 与我们的研究结果基本一致[24]。
已有研究表明, 由于干扰类型和强度的不同, 不同等级的道路对野生动物的影响有所差异[25]。与之类似, 研究结果表明村村通对夏季鸟类物种数的影响比公路小。这主要是由于村村通的交通流量较小, 由此产生的噪音等干扰较小; 此外, 村村通的修建使原本郁闭度较高的森林变得较为开阔, 异质性增高, 从而吸引了更多种类的鸟类在此活动。公路尽管也增加了生境的开阔性, 但其较大的交通流量和较快的车速给鸟类带来了强烈的干扰, 从而降低了其作为鸟类生境的适宜性。由于所调查的公路穿越森林生境, 所以, 这两种生境的鸟类组成基本没有差异, 而与村村通和村庄的夏季鸟类物种组成差异较大。
村庄及周边的人类活动一般较为频繁, 在此活动的鸟类多为伴人物种或对人为干扰耐受能力较强的物种, 如金腰燕、丝光椋鸟和麻雀等(表3)。森林景观中的村庄多为紧靠山林的自然村落, 如果村落规模不大, 人为干扰强度适中, 这些自然村落也可能对很多森林鸟类产生正面影响[26-27]。一方面, 村庄及其周边一般较为开阔, 生境类型也较为多样, 可以为多种鸟类提供所需的栖息地; 另一方面, 村庄中人类遗弃的食物及产生的垃圾为多种鸟类提供了丰富的食物资源。因此, 如果能适当控制村庄中人为干扰的强度, 村庄可以与森林鸟类和谐共存[28]。
由于村村通连接着村庄, 与村庄的干扰类型有相似之处, 所以村村通和村庄生境中夏季鸟类物种的组成较为接近, 这种规律在低海拔区域表现得更为明显(图2), 这是因为低海拔区域的村村通与村庄的干扰类型更为接近。此外, 海拔对夏季鸟类群落的影响还体现在低海拔区域村庄和村村通生境的鸟类物种数和香农威纳多样性指数高于高海拔区域相应的生境, 这说明在低海拔区域夏季鸟类对这两种适度干扰类型的适应性更强。
4 结论
鹞落坪自然保护区低海拔区域内夏季鸟类物种数和多样性指数均高于高海拔区域; 森林生境中鸟类物种数、个体数和多样性指数均较低, 与干扰较强的公路基本没有差异, 两种生境的鸟类群落物种组成也比较相似。村庄鸟类物种数、个体数和多样性指数均高于其他几种生境, 其物种组成与村村通生境较为相似。村村通与公路生境的鸟类个体数和多样性指数不存在显著差异, 但村村通生境中物种数显著高于公路生境。研究结果表明, 在森林景观中, 适度的人为干扰有利于增加夏季鸟类物种多样性。建议在森林鸟类的管理和保护中增加对自然村落周边鸟类的关注。
附表1 2017年鹞落坪夏季鸟类名录及分布
Appendix 1 List and distribution of summer avian in Yaoluping in 2017
续表
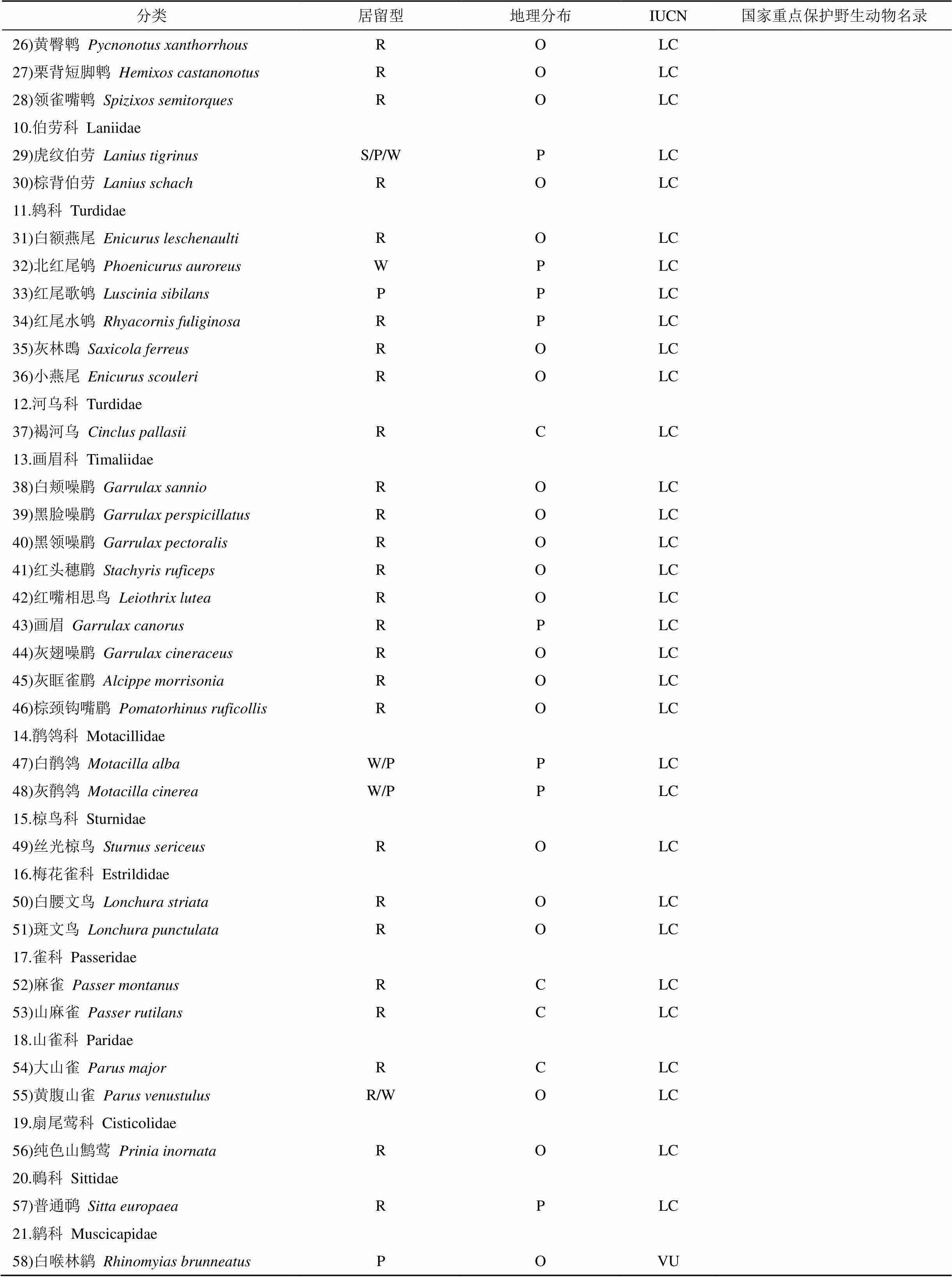
分类居留型地理分布IUCN国家重点保护野生动物名录 26)黄臀鹎Pycnonotus xanthorrhousROLC 27)栗背短脚鹎Hemixos castanonotusROLC 28)领雀嘴鹎Spizixos semitorquesROLC 10.伯劳科 Laniidae 29)虎纹伯劳Lanius tigrinusS/P/WPLC 30)棕背伯劳Lanius schachROLC 11.鸫科 Turdidae 31)白额燕尾Enicurus leschenaultiROLC 32)北红尾鸲 Phoenicurus auroreusWPLC 33)红尾歌鸲 Luscinia sibilansPPLC 34)红尾水鸲Rhyacornis fuliginosaRPLC 35)灰林鵖Saxicola ferreusROLC 36)小燕尾 Enicurus scouleriROLC 12.河乌科 Turdidae 37)褐河乌 Cinclus pallasiiRCLC 13.画眉科 Timaliidae 38)白颊噪鹛Garrulax sannioROLC 39)黑脸噪鹛Garrulax perspicillatusROLC 40)黑领噪鹛Garrulax pectoralisROLC 41)红头穗鹛Stachyris ruficepsROLC 42)红嘴相思鸟 Leiothrix luteaROLC 43)画眉Garrulax canorusRPLC 44)灰翅噪鹛Garrulax cineraceusROLC 45)灰眶雀鹛Alcippe morrisoniaROLC 46)棕颈钩嘴鹛Pomatorhinus ruficollisROLC 14.鹡鸰科 Motacillidae 47)白鹡鸰Motacilla albaW/PPLC 48)灰鹡鸰Motacilla cinereaW/PPLC 15.椋鸟科 Sturnidae 49)丝光椋鸟Sturnus sericeusROLC 16.梅花雀科 Estrildidae 50)白腰文鸟Lonchura striataROLC 51)斑文鸟Lonchura punctulataROLC 17.雀科 Passeridae 52)麻雀Passer montanusRCLC 53)山麻雀 Passer rutilansRCLC 18.山雀科 Paridae 54)大山雀Parus majorRCLC 55)黄腹山雀Parus venustulusR/WOLC 19.扇尾莺科 Cisticolidae 56)纯色山鹪莺Prinia inornataROLC 20.鳾科 Sittidae 57)普通䴓Sitta europaeaRPLC 21.鹟科 Muscicapidae 58)白喉林鹟Rhinomyias brunneatusPOVU
续表
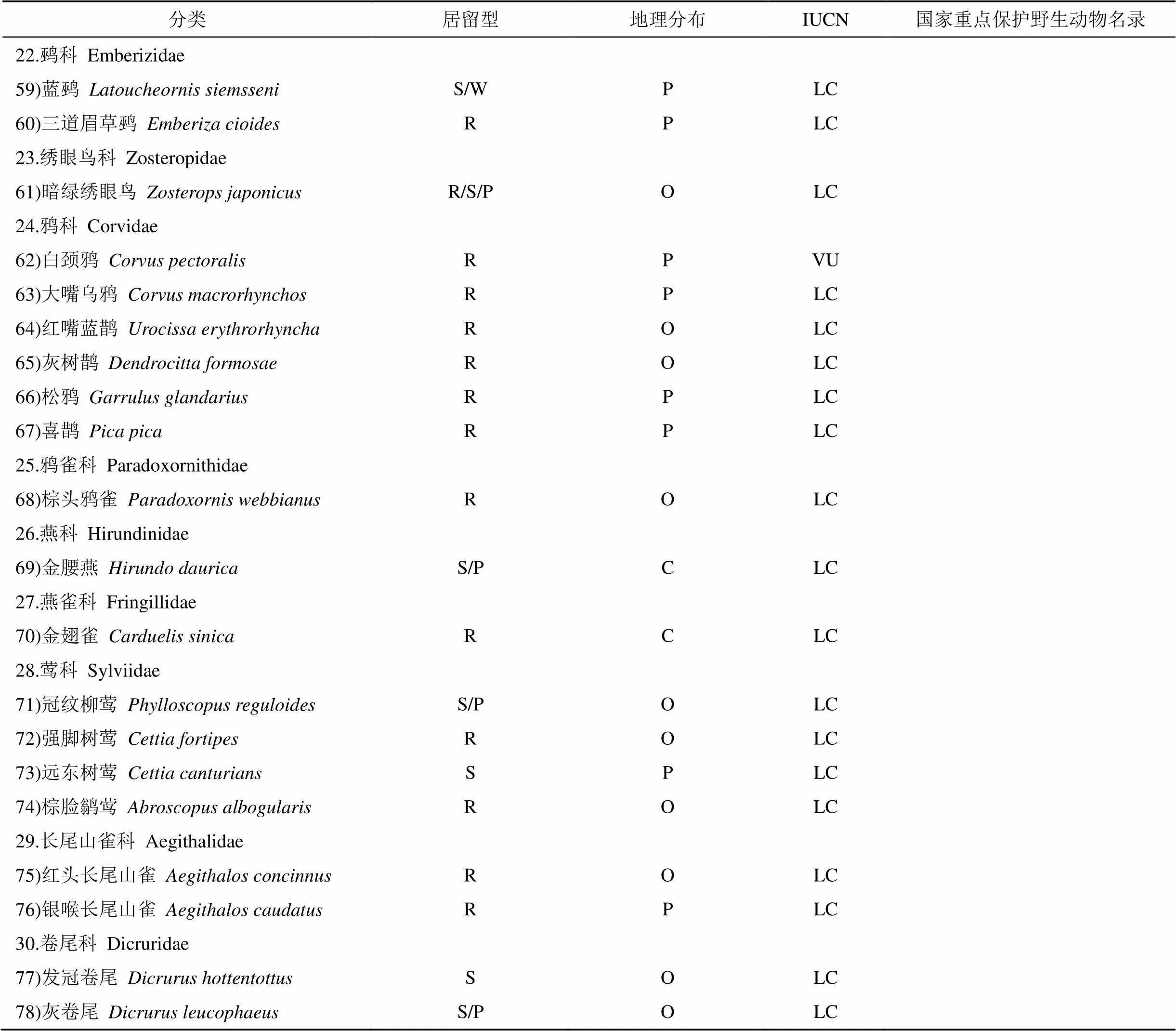
分类居留型地理分布IUCN国家重点保护野生动物名录 22.鹀科 Emberizidae 59)蓝鹀 Latoucheornis siemsseniS/WPLC 60)三道眉草鹀 Emberiza cioidesRPLC 23.绣眼鸟科 Zosteropidae 61)暗绿绣眼鸟Zosterops japonicusR/S/POLC 24.鸦科 Corvidae 62)白颈鸦Corvus pectoralisRPVU 63)大嘴乌鸦Corvus macrorhynchosRPLC 64)红嘴蓝鹊 Urocissa erythrorhynchaROLC 65)灰树鹊Dendrocitta formosaeROLC 66)松鸦 Garrulus glandariusRPLC 67)喜鹊Pica picaRPLC 25.鸦雀科 Paradoxornithidae 68)棕头鸦雀Paradoxornis webbianusROLC 26.燕科 Hirundinidae 69)金腰燕Hirundo dauricaS/PCLC 27.燕雀科 Fringillidae 70)金翅雀Carduelis sinicaRCLC 28.莺科 Sylviidae 71)冠纹柳莺Phylloscopus reguloidesS/POLC 72)强脚树莺Cettia fortipesROLC 73)远东树莺Cettia canturiansSPLC 74)棕脸鹟莺Abroscopus albogularisROLC 29.长尾山雀科 Aegithalidae 75)红头长尾山雀Aegithalos concinnusROLC 76)银喉长尾山雀Aegithalos caudatusRPLC 30.卷尾科 Dicruridae 77)发冠卷尾Dicrurus hottentottusSOLC 78)灰卷尾Dicrurus leucophaeusS/POLC
注:居留型: S—夏候鸟, W—冬候鸟, R—留鸟, P—旅鸟; 区系划分: P—古北界种类, O—东洋界种类, C—广布界物种; IUCN: EN—濒危, VU—易危, NT—近危, LC—低危; 国家重点保护野生动物名录: I—一级保护, Ⅱ—二级保护。
[1] SEKERCIOĞLU C H, WENNY D G, WHELAN C J. Why birds matter: Avian ecological function and ecosystem services[M]. Chicago: The University of Chicago Press, 2016.
[2] LIANG Chenxia, YANG Guisheng, WANG Na, et al.. Taxonomic, phylogenetic and functional homogenization of bird communities due to land use change[J]. Biological Conservation, 2019, 236: 37–43.
[3] YANG Qinli, ZHANG Heng, PENG Wanshan, et al. Assessing climate impact on forest cover in areas undergoing substantial land cover change using Landsat imagery[J]. Science of the Total Environment, 2019, 659(2): 732–745.
[4] 刘吉平, 张顺, 陈智文. 人类活动对三江平原东北部湿地鸟类的干扰[J]. 东北林业大学学报, 2008, 36(12): 40– 42.
[5] ZHANG Weiwei, SHI Jinze, HUANG Huiqin, et al. The impact of disturbance from photographers on the Blue-crowned Laughingthrush ()[J]. Avian Conservation and Ecology, 2017, 12(1): 15.
[6] 宋景舒, 胡洁, 江波, 等. 天祝夏玛林场不同生境夏季鸟类多样性[J]. 生态与农村环境学报, 2020, 36(5): 606– 611.
[7] MARION S, DAVIES A, DEMŠAR U, et al. A systematic review of methods for studying the impacts of outdoor recreation on terrestrial wildlife[J]. Global Ecology and Conservation, 2020, 22: e00917.
[8] PEH K S-H, JONG J D, SODHI N S, et al. Lowland rainforest avifauna and human disturbance: persistence of primary forest birds in selectively logged forests and mixed-rural habitats of southern Peninsular Malaysia[J]. Biological Conservation, 2005, 123(4): 489–505.
[9] TEJEDA-CRUZ C, SUTHERLAND W J. Cloud Forest Bird Responses to Unusually Severe Storm Damage[J]. Biotropica, 2005, 37(1): 88–95.
[10] PERZ S G, CABRERA L, CARVALHO L A, et al. Regional integration and local change: road paving, community connectivity, and social-ecological resilience in a tri-national frontier, southwestern Amazonia[J]. Regional Environmental Change, 2012, 12(1): 35–53.
[11] LAURANCE W F, GOOSEM M, LAURANCE S G W. Impacts of roads and linear clearings on tropical forests[J]. Trends in ecology & evolution, 2009, 24(12): 659–669.
[12] LAURANCE S G W, STOUFFER P C, LAURANCE W F. Effects of Road Clearings on Movement Patterns of Understory Rainforest Birds in Central Amazonia[J]. Conservation Biology, 2004, 18(4): 1099–1109.
[13] LEES A C, PERES C A. Gap-Crossing Movements Predict Species Occupancy in Amazonian Forest Fragments[J]. Oikos, 2009, 118(2): 280–290.
[14] CLAIR T C C S. Factors Affecting the Permeability of Transportation and Riparian Corridors to the Movements of Songbirds in an Urban Landscape[J]. Journal of Applied Ecology, 2009, 46(6): 1314–1322.
[15] SPOONER P G, SMALLBONE, L. Effects of road age on the structure of roadside vegetation in south-eastern Australia[J]. Agriculture, Ecosystems and Environment, 2009, 129(1/3): 57–64.
[16] GARAFFA P I, FILLOY J, BELLOCQ M I. Bird community responses along urban–rural gradients: Does the size of the urbanized area matter? [J]. Landscape and Urban Planning, 2009, 90(1/2): 33–41.
[17] MAMMIDES C, KOUNNAMAS C, GOODALE E, et al. Do unpaved, low-traffic roads affect bird communities? [J]. Acta Oecologica, 2016, 71: 14–21.
[18] 周磊, 万雅琼, 洪欣, 等. 利用红外相机技术对安徽省鹞落坪国家级自然保护区大中型兽类及林下鸟类的调查[J]. 生物多样性, 2018, 26(12): 1338–1342.
[19] 钱者东, 陈皓, 张昊楠, 等. 安徽鹞落坪自然保护区聚落用地适宜性评价[J]. 生态与农村环境学报, 2019, 35(11): 1412–1419.
[20] 郑光美. 中国鸟类分类与分布名录[M]. 北京: 科学出版社, 2011.
[21] MINCHIN P R. An Evaluation of the Relative Robustness of Techniques for Ecological Ordination[J]. Vegetatio, 1987, 69(1-3): 89–107.
[22] ROSIN Z M, SKÓRKA P, PÄRT T, et al. Villages and their old farmsteads are hot spots of bird diversity in agricultural landscapes[J]. Journal of Applied Ecology, 2016, 53(5): 1363–1372.
[23] CHETTRI N, DEB D C, SHARMA E, et al. The Relationship Between Bird Communities and Habitat[J]. Mountain Research and Development, 2005, 25(3): 235– 243.
[24] 胡静, 唐雪海, 黄庆丰, 等. 鹞落坪自然保护区土地利用动态格局研究[J]. 水土保持通报, 2019, 39(6): 246–251, 323.
[25] WOLF I D, HAGENLOH G, CROFT D B. Vegetation moderates impacts of tourism usage on bird communities along roads and hiking trails[J]. Journal of environmental management, 2013, 129(15): 224–234.
[26] 骆鹰, 李常健, 万亮, 等. 湖南省永州市鸟类区系特征及群落结构研究[J]. 四川动物, 2014, 33(5): 750–757.
[27] 高斌, 陈鹏, 杨菲, 等. 阳际峰国家自然保护区夏季鸟类群落结构调查[J]. 南昌大学学报(理科版), 2017, 41(1): 90–96.
[28] DUBAY S G, REEVE A H, WU Yongjie. Human disturbance provides foraging opportunities for birds in primary subalpine forest[J]. Journal of Ornithology, 2017, 158(3): 833–839.
Bird diversity differences in four different habitats in Yaoluoping Nature Reserve in summer
ZHAO Binbin, XU Wenjiu, LI Chunlin*
SchoolofResourcesandEnvironmentalEngineering, AnhuiUniversity, Anhui230601, China
Investigating differences in bird communities among various habitats can provide scientific basis for the management and conservation. We surveyed forest birds in areas of different elevations in Yaoluoping National Nature Reserve from July to August, 2017, and compared summer bird communities among forests, highways, village-to-village roads and villages. We recorded a total of 4225 birds belonging to 9 orders and 30 families. Due to the high canopy cover and simplified vegetation, bird species richness, number of individuals and species diversity were all low in forests, not different from those along highways crossing forests. Although there were no significant differences in number of individuals and species diversity between village-to-village road and highways, species richness along village-to-village roads was significantly higher than that along highways, which might be related to the lower disturbances. Species richness, number of individuals and Shannon-Wiener species diversity index in villages were significantly higher than those in other habitats, which could be attributed to the relatively open habitats and rich food resources from humans in villages. Our study indicates that the species diversity of summer bird communities is higher in habitats with greater disturbance such as villages in forests.
bird community; Yaoluoping; species diversity; road; village
10.14108/j.cnki.1008-8873.2022.06.008
Q958.1
A
1008-8873(2022)06-063-10
2020-09-28;
2020-11-05基金项目:国家自然科学基金项目(31770571, 31970500); 生态环境部生物多样性调查、观测和评估项目
赵彬彬(1992—), 男, 安徽淮北人, 硕士, 主要从事鸟类生态学研究, E-mail: 885045538@qq.com
通信作者:李春林, 男, 博士, 副教授, 主要从事鸟类生态学等方面研究, E-mail: lichunlin1985@163.com
赵彬彬, 徐文玖, 李春林. 鹞落坪保护区四种不同生境的夏季鸟类多样性差异[J]. 生态科学, 2022, 41(6): 63–72.
ZHAO Binbin, XU Wenjiu, LI Chunlin. Bird diversity differences in four different habitats in Yaoluoping Nature Reserve in summer[J]. Ecological Science, 2022, 41(6): 63–72.
