生态位构建对具有资源竞争物种的进化分布动态的影响
2022-09-17李建媚韩晓卓
李建媚, 韩晓卓
生态位构建对具有资源竞争物种的进化分布动态的影响
李建媚, 韩晓卓*
广东工业大学数学与统计学院, 广州 510520
生态位构建是有机体通过自身活动、代谢等行为调节或影响其环境或其他生物有机体的过程, 进而导致有机体在自然选择过程中产生不同的进化轨迹。已有研究结论是基于双位点种群遗传模型、基于个体模型以及及元胞自动机模拟模型得出的。目前分析物种进化入侵的常用方法之一是进化分布动态模型。通过引入该方法,着重研究了依特征调节的生态位构建作用对具有种内资源竞争的物种进化动态的影响。结果表明: 依特征调节的生态位构建确实能够引起物种进化轨迹的明显改变。若生态位构建中心位于特征值较小的特征时, 将减少进化分支, 且相较于不存在生态位构建的情况而言, 种群密度提高; 与此相反, 生态位构建中心为特征值较大的特征时,生态位构建维持进化多样性, 尽管其破坏了原有进化的对称分布。这意味着生态位构建通过与环境长期的正反馈作用, 使具有较强依赖的种群特征出现明显差异, 分支数虽有减少, 但显著提升了优势特征的种群密度, 且物种进化分支数随着构建强度的增加而减少。强烈的种内竞争和较弱的生态位构建作用将有利于维持种群的进化多样性。
进化分支; 进化多样化; 生态位构建; 竞争核; 对称分布
0 前言
生态位构建理论(Niche Construction Theory, NCT)是指有机体通过自身的新陈代谢、活动和选择行为, 部分的创建或毁灭自身生态位、改变环境、进而改变其环境中生物与非生物的自然选择源的一种能力[1-2]。简单地讲, 生态位构建体现的是有机体与环境在进化过程中的相互反馈, 从而使进化成为自然选择与生态位构建共同作用的过程。该理论强调了有机体在进化过程中不仅是自然选择的被动受体, 也是一个主动的“工程师”改变着自身的环境[3-7]。这一概念自被提出以来, 许多生态学家和进化生物学家进行了大量的实验和理论研究[3-7]。结果表明, 有机体与环境之间的这种反馈作用可以触发调节机制, 从而改变其自身的进化轨迹[8-13]。因此, 该理论的提出在一定程度上扩展和丰富了现有的进化思想, 不仅为物种适应性研究提供了新颖的理论依据, 而且也为解释不同尺度上的生态学现象提供了有效的机理模式[1-2,6,9]。
目前, 有关生态位构建的理论性成果大部分基于群体遗传学模型, 将其作用于个体、结构种群、集合种群等不同尺度, 用以评估生态位构建对不同生物类型的进化动态的影响[9-17]。最早, Laland教授和他的团队于1996年建立了具有生态位构建作用的双位点种群基因模型, 通过在该模型中引入基因型频率与资源环境动态的耦合关系, 证实了生态位构建作用确实可以产生多样的进化动态, 揭示了生态位构建可以作为潜在的进化推动者[9]。随后, 惠苍和韩晓卓等人将该模型发展到具有空间结构的种群水平和集合种群上, 得到了一系列理论结果[8, 10-13]。例如, 生态位构建可以导致结构种群多态分布的空间模式, 有利于种群稳定多态的形成; 由生态位构建引发的有机体-环境关系可对集合种群的动态、竞争以及多样性产生深入的影响[10]。生态位构建在空间生境上可以形成“生态烙印”, 导致环境异质性与物种分布界限间的自组织结构, 从而弱化竞争系统中的空间竞争强度[11-13]。
基于有机体与环境之间的微分动力系统来探究生态位构建对动植物的进化动态或生态响应的影响也是部分研究所采用的主要方法[14-17]。在这些模型中, 空间异质性被认为是具有生态位构建特征的稀有物种能够在敌对环境中持续存在的重要因素[14]。作为植物和环境循环系统中的重要机制, 生态位构建同时也影响植物的进化分布和自身土壤养分库的适应性调节[15]。在具有双物种资源竞争的捕食与被捕食系统中也得出了类似的结果, 物种可通过生态位构建作用调节种内竞争和种间竞争之间的平衡[16]。当然, 在自然界中也存在许多例子来阐明有机体的生态位构建确实具有推动进化的作用, 这其中包括蚯蚓、海狸、布谷鸟、松树和一些病原体。也有研究表明, 生态位构建能够导致微生物种群快速进化是一种常见的现象, 类似于酵母菌(酿酒酵母)以改变其在水果中的微生物环境来吸引果蝇, 从而促进其自身繁殖的现象[17]。
然而, 利用种群遗传学模型或微分动力系统研究生态位构建的进化或生态响应具有一定的局限性, 这些结果仅从种群遗传学的角度或常规的有机体与资源的耦合关系入手。随着研究方法或进化理论的不断发展, 进化分布动态(Evolutionary Distribution Dynamics)和自适应动态(Adaptive Dynamics)已成为目前国际上研究进化时常用的进化入侵分析的主要方法。它们常被用来从表型特征的角度出发探讨进化生物学的核心问题, 特别是涉及与进化多态性和多样化有关的问题[18-21]。虽然这两种方法都以物种的生物特征为研究对象, 但有其各自的适用条件。自适应动态适用于当适合度密度依赖或频率依赖由稀有且微小的突变所引起的进化改变[22-23]; 不过, 该方法以平均性状的进化为关注点, 常忽视特征之间的差异性, 有可能忽略掉重要的进化特征[24]。而在进化分布动态中, 性状分布的演化是在连续的特征谱中进行, 其优点在于一旦进化动态达到稳定, 则该状态代表了一系列所有可能突变体的进化稳定策略[25-26]。
本文将以进化分布动态为研究方法, 尝试性地建立具有生态位构建作用的特征与依特征调节的种内资源竞争相结合的物种进化动态的基本模型, 着重分析该特征所引起的进化分布动态及其对进化多样性的影响。就研究方法来说, 本文所获结果是对生态位构建理论在进化层面做出的有效补充, 具有一定的理论意义。
1 研究方法与模型建立
1.1 研究方法
进化分布动态是以物种的生物特征为研究对象, 基于偏微分方程中的反应扩散方程发展而来的一种进化入侵分析工具, 其中, 方程中的反应项代表进化中的自然选择过程, 扩散项代表特征的突变过程[25-26]。选择该方法作为研究生态位构建所引起进化效应的另一种有效途径, 是基于物种的某些生物特征是具有生态位构建作用的事实。例如, 控制寄生虫的体型大小的特征会影响其遇到不同宿主的几率或改变宿主环境的能力[27]。宿主与寄生虫相互作用的演化过程就可以看作是生态位构建的模式。同样, 病原体与宿主之间的进化也符合生态位构建的特点[28]。病原体的生命周期往往比宿主的生命周期快得多, 以特定的方式改变宿主环境的病原体突变也会不可避免地改变其自身的进化轨迹。因此, 物种的生态位构建作用可以被看成是性状依赖的, 这是建立模型的根本。
1.2 模型建立
首先, Doebeli 和 Ispolatov为阐明由简单的资源竞争便可维持生物多样性的机制, 于2010年提出了一个经典的频率依赖的竞争模型[29]。在时刻, 特征为的种群密度记为(,), 则其依特征频率的变化动态可由如下偏微分方程给出:
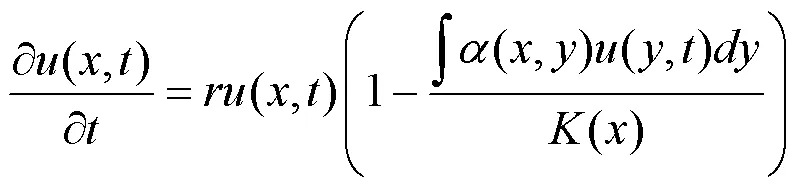
其中,为种群的内禀增长率;(,)反应了具有表型与表型的个体之间的竞争剧烈程度, 称之为竞争核, 通常被假设为距离|–|的单峰函数。(,)(,)是所有特征为的种群(,)与特征的种群产生竞争关系的效应总和。()是环境承载函数, 假设在=0处取得最大值的单峰函数。通常竞争核和承载能力选用高斯函数形式:(,)=exp(–(–)2)和()=0exp(–2), 参数>0和>0用于衡量频率依赖的竞争部分和自然选择部分的相对重要性, 常数0则表示环境的最大承载能力。在动态模型(1)中, 当<时, 种群动态收敛于=0的峰值; 当>时, 系统趋于有多个峰值的正态分布的平衡状态, 即证实了强烈的频率依赖性可以保持物种多样性[29]。
进化分布动态方法是反应扩散方程的进一步拓展, 描述了物种特征在连续空间中的进化分布动态。如果在上述动态模型(1)中引入扩散项, 即可给出频率依赖的竞争物种的进化分布动态[30]:
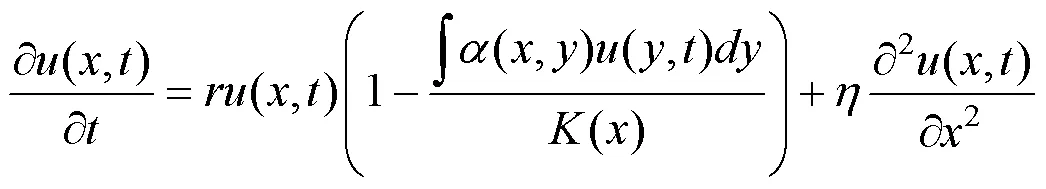
其中, 扩散项系数在这里表示特征的突变率, 如无特别说明, 本文取=0.000001。
生态位构建被认为是通过有机体及其环境之间的积极反馈而能够产生意想不到的进化结果的力量[9, 31]。通常, 有机体及其周围环境的反馈作用被用来描述生态位构建的动态过程[32-34]。在依特征调节的进化分布动态(2)中, 生态位构建引起生物与环境之间反馈的关键是考虑到承载能力()不再是静止的状态, 而是受到特征影响随时间而变化。根据Laland模型的基本框架[9], 假设资源承载量的变化受生态位构建活动影响, 且部分依赖于该特征的构建能力以及种群密度所占比例, 其中, 正(负)生态位构建过程分别表示构建引起资源的增加(减少)。除此之外, 资源本身也存在自身更新和消耗的过程, 可用方程表示如下:
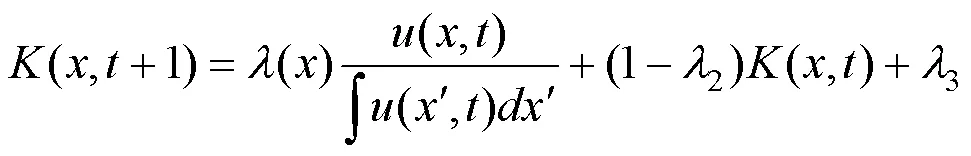
其中,()表示特征依赖的生态位构建能力, 简称生态位构建核, 选用生物学中常见的Sigmoid函数:()=1/(1+exp(–(–0))), 意味着特征越大, 构建能力越强。参数1不仅控制生态位构建能力的最大值, 还反映构建的正负方向。本文只考虑发生正生态位构建的情况, 即1>0; 而参数>0的大小意味着构建能力在中心点0附近的变化趋势;2和3两个参数分别为决定资源独立损耗和更新程度的系数。当环境承载能力为动态变化时, 方程(2)则相应地改写为:
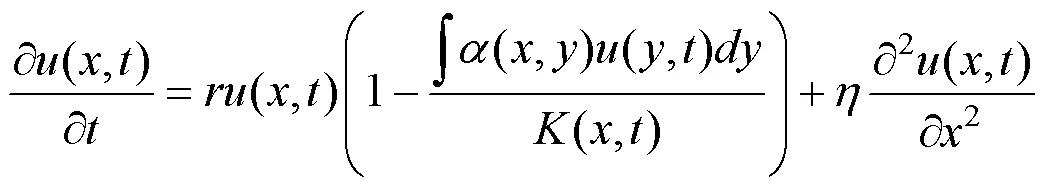
结合方程(3)和(4), 即可探讨生态位构建作用下的特征依赖的物种进化动态。特别地, 当方程(3)中的参数1=0时, 意味着进化动态不受生态位构建的影响, 此时环境容纳量将保持在固定的平衡值equ=1/2, 那么, 该系统退化为方程(2), 其结果即为具有资源竞争影响模式下的经典结论[30]。
1.3 数值模拟
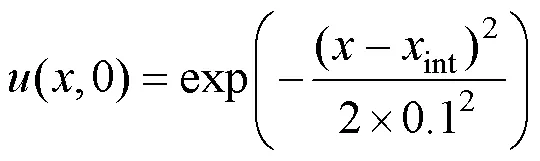
2 结果与分析
首先通过对照分析给出生态位构建作用对特征进化分布动态的影响。当系统不具有生态位构建(1=0)时, 该系统特征的进化分布情况与经典进化分布模型的结果一致[30], 见图1(a), 即由一个初始特征演化出6个进化分支(Evolutionary Branching), 并呈现出对称分布模式。然而, 当存在特征依赖的生态位构建时, 即1≠0, 进化分布动态受构建核中的参数1,0和的共同作用而发生明显改变。当生态位构建核的中心0位于种群初始特征int左侧(0=0.5)时, 初始特征得以保留(图1(b)), 并未分化, 在其两侧的特征各自分化后, 整体演化出5个进化分支; 当0的位置与初始特征int重合(0=1.5)时, 其进化分支分布模式与图1(b)相似(见图1(c)); 而当生态位构建核的中心0位于初始特征int右侧(0=2.5)时, 种群初始特征受生态位构建的影响逐渐减小, 并于初始特征附近形成分支; 尽管整体与不受生态位构建影响下的对称分布动态相似, 但在右侧演化出的分支间距变大, 对称模式被破坏, 并且中心分支发生所需的进化时间延迟(见图1(d))。
同时, 生态位构建显著改变系统平衡后的种群密度(见图2)。当生态位构建发生, 构建核的中心0位于初始特征左侧(0=0.5)以及与初始特征重合(0=1.5)时, 系统整体种群密度较无生态位构建作用时显著增加, 且最右侧特征值较大的特征的密度明显升高, 初始特征的种群被保留, 且密度维持在一定的水平上(见图2(b)和图2(c))。但随着生态位构建核的中心0向右移动, 整体种群密度逐渐减少, 当构建核的中心移至初始特征右侧(0=2.5)时, 初始特征消失, 其进化分支密度明显减少, 最右侧特征密度具有明显优势(见图(d))。
为了综合体现生态位构建核对系统的影响, 选定1=1.5, 根据不同的值和0值, 绘制每一组参数下的()函数值与种群进化分支数的对应关系, 如图4所示。结果显示: 生态位构建核取值较小时(()<0.4), 进化分支数量为6; 当生态位构建核取值位于0.4—0.8之间, 种群的进化分支数量减少为5; 直到()>0.8后, 进化分支数量为4, 该结果意味着: 构成()函数的三个参数1、和0, 也即生态位构建的强度、特征依赖程度以及发生的位置共同影响着种群的进化分支数, 并且分支数随该函数值的增加而减小。综合考虑到图2的结果, 这说明生态位构建通过与环境长期的正反馈作用, 使得具有较强依赖的种群特征差异明显, 分支数虽有减少, 但明显提升了优势特征的种群密度。
另外, 已有研究已表明竞争核σ的宽度变化对单纯的资源竞争系统的进化动态起到决定性作用[29-30]。引入特征相关的生态位构建后, 重点观察构建作用与竞争强度对种群进化动态的共同影响, 见图5。在固定值和0值的情况下, 随着种内竞争核宽度的逐渐增大, 特征的进化分支也逐渐减少; 随着生态位构建强度的增加, 当生态位构建强度1=0.5时, 若种内竞争核狭窄(σ=0.04), 进化分支不变, 若种内竞争核宽度增加(σ>0.04), 则减少一个进化分支。且随着生态位构建强度的逐渐增强, 环境中生态位构建的能力得到增强, 已具备生态位构建作用的生态系统需要更强烈的生态位构建才能对进化分支产生影响。事实上, 种内竞争核宽度的逐渐增大, 导致连续性状之间存在越来越大的差异, 因此使得种内竞争减少, 分支数量减少。同时, 生态位构建强度的增强, 导致在局部环境中与特征相关的生态位构建能力也增强, 使得特征构建适宜自身生存的繁殖环境的能力增强, 周围能够满足其生存和发展需要的生态位增多, 特征的进化选择压力减少不产生分支, 导致进化分支数量减少。狭窄的竞争核以及较弱的生态位构建强度可维持生态系统中的物种多样性。
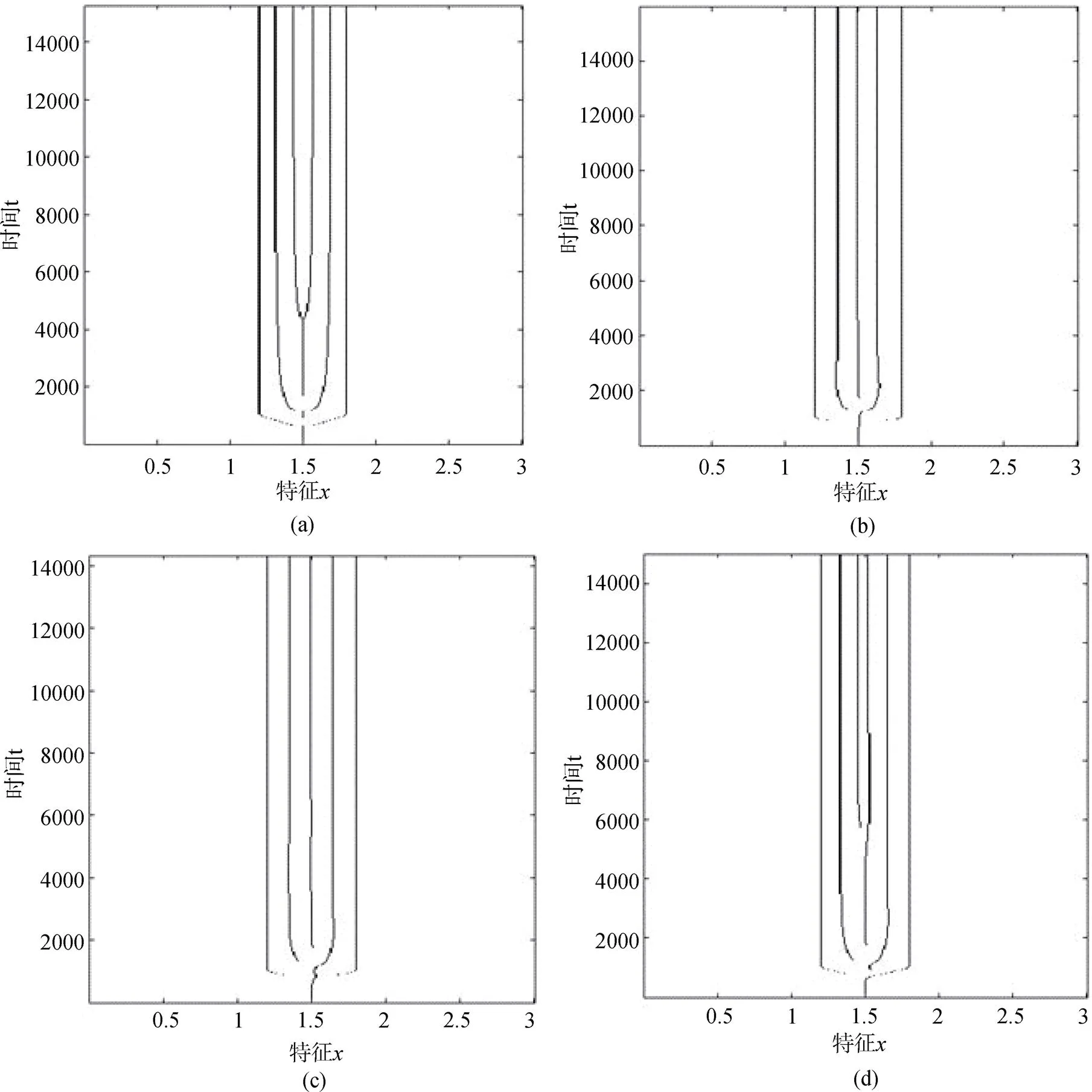
图1 不同模式下的进化分布动态(a.没有生态位构建, 参数λ1=0; b.存在生态位构建, 参数为λ1=0.6, x0=0.5; c.存在生态位构建, 参数为λ1=0.6, x0 =1.5; d.存在生态位构建, 参数为λ1=0.6, x0 =2.5; 其他参数为σA=0.06, λ2=0.01, λ3=4, k=1, σk=3)
Figure 1 Evolutionary distribution dynamics (a. without niche construction, parameters1=0; b. with niche construction, parameters are1=0.6,0=0.5; c. with niche construction, parameters are1=0.6,0=1.5;d. with niche construction, parameters are1=0.6,0=2.5; other parameters areσ=0.06,2=0.01,3=4,=1,σ=3)
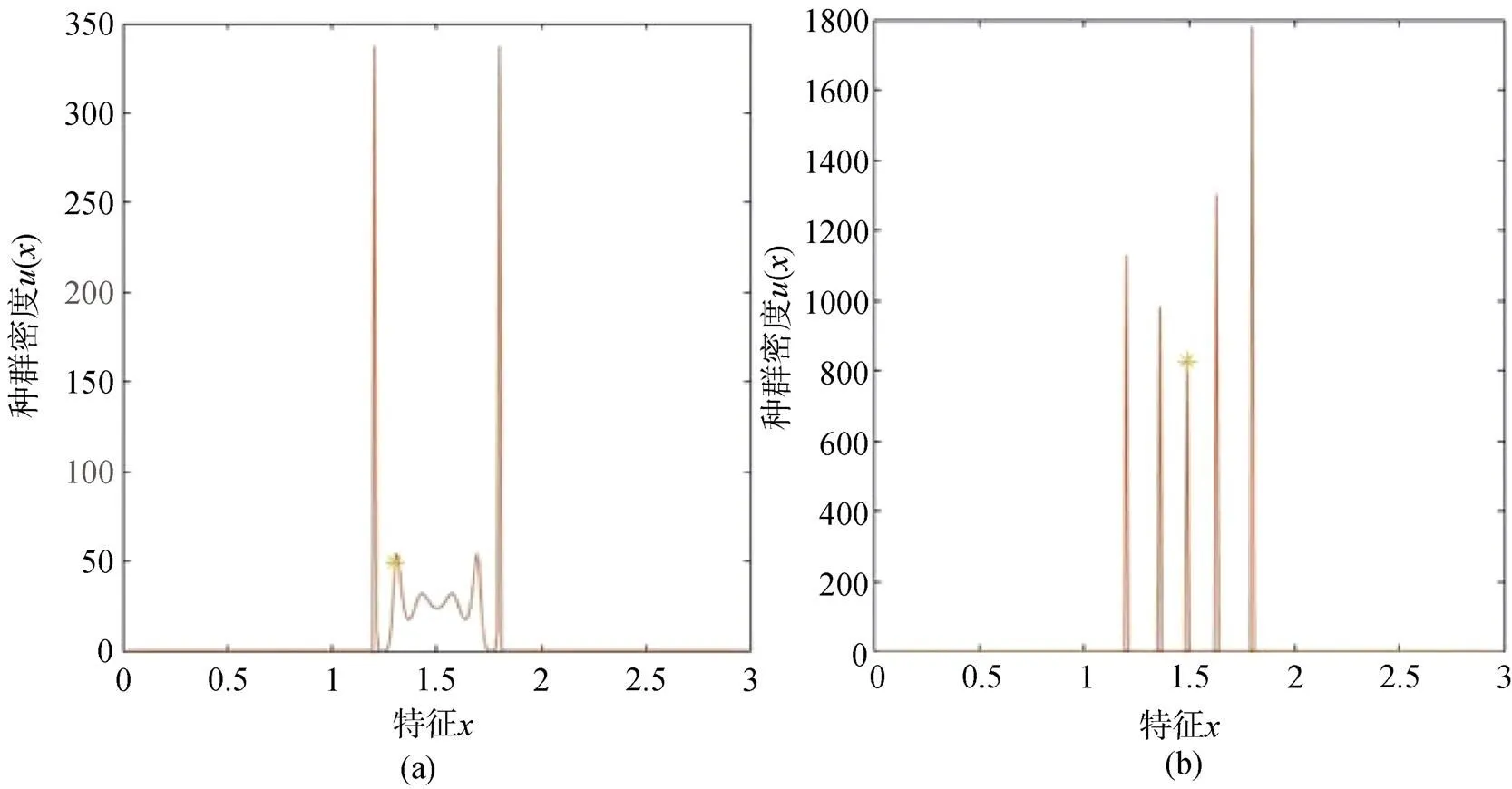
Figure 2 Density distribution of species equilibriums(the parameters are respectively corresponding with those in Figure 1)
图3 生态位构建作用下的进化分布动态(a. k=5, x0=1.5; b. k=5, x0=2.5; c. k=0.3, x0=2.5; 构建系数λ1=0.6, 其他参数同图1a)
Figure 3 Evolutionary distribution dynamics with niche construction(a.=5,0=1.5; b.=5,0=2.5; c.=0.3,0=2.5; the intensity of niche construction is1=0.6; other parameters are the same as those in Figure 1a)

图4 不同生态位构建模式下的进化分支数图(参数为λ1=1.5, σA=0.06, λ2=0.01, λ3=4, σk=3)
Figure 4 The number of evolutionary branching with niche construction(parameters are1=1.5,σ=0.06,2=0.01,3=4,σ=3)
3 讨论
进化分布动态是进化入侵分析的主要方法之一,对于研究连续适应性特征具有较大的优势[8,33]。大多数研究人员利用适应性动力学来探索进化多样化, 但该方法局限于平均性状的进化可能忽略了重要的进化特征。在本文中, 使用进化分布动态方法来探讨物种特征的生态位构建模型, 以揭示生态位构建对进化多样化的影响。与之前已有的研究工作相比, 本文所采用的新的分析工具更适用于此类问题的研究。
实验结果表明, 特征相关的生态位构建对进化分布动态产生显著的影响。生态位构建对进化分布动态的影响因生态位构建能力强弱的参数1, 生态位构建核的中心0, 种内竞争核σ以及构建核中的参数变化而变化。生态位构建核的中心位置变化, 其他参数不变时, 将特征值较小的特征作为生态位构建的中心, 特征值大于该特征的特征具有的生态位构建能力均强于该特征, 在该特征保留的情况下, 特征值较大的特征或其连续特征也得以保留, 且整体种群密度增加, 特征值较大的一侧特征密度增加明显。将特征值较大的特征作为生态位构建的中心, 特征值小于该特征的特征具有的生态位构建能力均弱于该特征, 因此生态位构建对这些特征的影响并不明显, 仅破坏了不存在生态位构建作用时的对称分布模式, 同时随着生态位构建中心逐渐向右移动, 环境中整体种群密度逐渐减少。构建核中的参数也对进化分布动态造成影响, 较大的参数表示生态位构建能力在相邻特征间的变化变的剧烈。这会导致特征无法适应与较大特征的竞争, 进化的选择压力增大, 进化分支增多。生态位构建能力强弱的参数1与种内竞争核σ共同影响进化分布动态, 狭窄的竞争核以及较弱的生态位构建强度可维持生态系统中的物种多样性。其中, 竞争核的宽度直接影响选择结果, 进而影响进化多样性。竞争核的宽度越窄, 特征竞争越激烈, 导致扩散, 这是出现进化多样化的一个普遍条件, 这一结果也与关于竞争核的已有结论一致[29-30]。而生态位构建的强度影响特征构建自身生态位的能力。生态位构建强度越弱, 特征构建适应其自身生存和繁衍生态位的能力越小, 使得自身选择压力增大, 导致扩散。当竞争核σ较宽, 种内竞争较弱, 生态位构建能力强弱1较强, 生态位构建能力较强时, 特征的进化选择压力减小, 进化分支减少。其他研究也得出了类似的结论, 即适度的生态位构建可能是提高物种在进化和生态尺度中的持久性、共存性和多样性的关键因素[10-13,33]。
Figure 5 The effect of evolutionary branches caused by competition kernelσand the width of niche construction1(the parameters areσ=3,=1,0=1.5,2=0.01,3=4, in which◆represents 8 branching, ●represents 7 branching, ■represents 6 branching, ★represents 5 branching, ◆represents 4 branching, ▲represents 3 branching,◢represents 2 branching,◣represents 1 branching)
需要说明的是, 该模型只考虑了一种简单的特征相关的生态位构建模式, 文中的资源容纳量是指具有特定特征的个体可以访问的公共资源, 物种的初始生长速度仅取决于资源容纳量。事实上, 我们可以做出一个更复杂的假设, 即资源与其环境中的某一特征有关。例如, 寄居蟹的壳或洞穴的大小是衡量资源含量的一个特征。以这种方式, 生物与其环境之间的反馈将通过将两个各自特征的耦合关系建立起来。这就是生态位构建理论中提到的, 发生在非生物环境的真正的进化动力学。在生物体与其生物环境之间的相互作用中, 生态位构建也更有可能出现。在多物种共同进化的背景下, 阐明生态位构建的效果至关重要。有证据表明, 一个物种的生态位构建过程由于改变了周围环境影响了共存于同一环境的其他物种的进化过程[30,37]。因此, 考虑具有生态位构建的物种之间的协同进化是接下来计划进行的工作。
4 结论
以特征调节的生态位构建为研究对象, 建立了具有生态位构建作用的物种在资源竞争作用下的进化分布动态模型, 探究了生态位构建和竞争作用对物种进化多样性的影响。基于特征值越大对应的生态位构建能力越强的假设, 研究结果表明: 随着生态位构建能力的增强, 对应特征的种群密度也增强。若生态位构建能力在相邻特征间的变化较大, 则导致特征的自然选择的进化压力增大, 从而进化分支增多。同时, 生态位构建能力的强弱与种内竞争核的宽度共同影响进化分布动态, 狭窄的竞争核以及较弱的生态位构建强度可维持生态系统中的物种多样性。
[1] ODLING-SMEE J, ERWIN D H, PALKOVACS E P, et al. Niche construction theory: a practical guide for ecologists[J]. Quarterly Review of Biology, 2013, 88(1): 3–28.
[2] MATTHEWS B, MEESTER L D, JONES C G, et al. Under niche construction: an operational bridge between ecology, evolution, and ecosystem science[J]. Ecological Monographs, 2014, 84(2): 245–263.
[3] JONES C G, LAWTON J H, SHACHAK M. Positive and negative effects of organisms as physical ecosystem engineers[J]. Ecology, 1997, 78(7): 1946–1957.
[4] JONES C G, LAWTON J H, SHACHAK M. Organisms as ecosystem engineers[J]. Oikos, 1994, 69(3): 373–386.
[5] BEERLING D J. Leaf evolution: gases, genes and geochemistry[J]. Annals of Botany, 2005, 96(3): 345-352.
[6] DONOHUE K. Niche construction through phenological plasticity: life history dynamics and ecological conesquences[J]. New Phytologist, 2005, 166(1): 83–92.
[7] FLACK J C, GIRVAN M, DE WAAL F B M, et al. Policing stabilizes construction of social niches in primates[J]. Nature, 2006, 439(7075): 426-429.
[8] HAN Xiaozhuo, HUI Cang, HUANG Yaling. Spatial distributions of niche-constructing populations[J]. Computational Ecology and Software, 2015, 5(4): 286–298.
[9] LALAND K N, ODLING-SMEE F J, FELDMAN M W. Evolutionary consequences of niche construction and their implications for ecology[J]. Proceedings of the National Academy of Sciences USA, 1999, 96(18): 10242–10247.
[10] HUI Cang, YUE Dongxia. Niche construction and polymorphism maintenance in metapopulations[J]. Ecological Research, 2005, 20(2): 115–119.
[11] HAN Xiaozhuo, LI Zizhen, HUI Cang, et al. Polymorphism maintenance in a spatially structured population: A two-locus genetic model of niche construction[J]. Ecological Modelling, 2006, 192(1-2): 160–174.
[12] HAN Xiaozhuo, CHEN Baoying, HUI Cang. Symmetry breaking in cyclic competition by niche construction[J]. Applied Mathematics and Computation, 2016, 284(7): 66–78.
[13] HAN Xiaozhuo, HUI Cang. Niche construction on environmental gradients: the formation of fitness Valley and stratified genotypic distributions[J]. Plos One, 2014, 9(6): 1–9.
[14] SILVER M, PAOLO E D. Spatial effects favor the evolution of niche construction[J]. Theoretical Population Biology, 2006, 70(4): 387–400.
[15] KYLAFIS G, LOREAU M. Ecological and evolutionary consequences of niche construction for its agent[J]. Ecology Letters, 2010, 11(10): 1072–1081.
[16] KYLAFIS G, LOREAU M. Niche construction in the light of niche theory[J]. Ecology Letters, 2011, 14(2): 82–90.
[17] BUSER C C, NEWCOMB R D, GASKETT A C, et al. Niche construction initiates the evolution of mutualistic interactions[J]. Ecology Letters, 2015, 17(10): 1257–1264.
[18] METZ J A J, NISBET R M, GERITZ S A H. How should we define 'fitness' for general ecological scenarios?[J]. Trends in Ecology and Evolution, 1992, 7(6): 198–202.
[19] DOEBELI M, DIECKMANN U. Evolutionary branching and sympatric speciation caused by different types of ecological interactions[J]. The American naturalist, 2000, 156(S4): S77–S101.
[20] ZHANG Feng, HUI Cang, PAUW A. Adaptive divergence in Darwin’s race: how coevolution can generate trait diversity in a pollination system[J]. Evolution, 2013, 67(2): 548–560.
[21] LANDI P, HUI Cang, DIECKMANN U. Fisheries-induced disruptive selection[J]. Journal of Theoretical Biology, 2015, 365: 204–216.
[22] DIECKMANN U, LAW R. The dynamical theory of coevolution: a derivative from stochastic ecological processes[J]. Journal of mathematical biology, 1996, 34(5-6): 579–612.
[23] SASAKI A, DIECKMANN U. Oligomorphic dynamics for analyzing the quantitative genetics of adaptive speciation[J]. Journal of Mathematical Biology, 2011, 63(4): 601–635.
[24] ANDRIAMIHAJA R, AZIZ O, HUI Cang. Spatial assortment of mixed propagules explains the acceleration of range expansion[J]. Plos One, 2014, 9(8): e103409.
[25] COHEN Y. Distributed predator-prey co-evolution[J]. Evolutionary Ecology Research, 2003, 5(6): 819–834.
[26] COHEN Y. Evolutionary distributions[J]. Evolutionary Ecology Research, 2009, 11(4): 611–635.
[27] LYMBERY A J. Niche construction: evolutionary implication for parasites and host[J]. Trends in Parasitology, 2015, 31(4): 134–141.
[28] BONI M F, FELDMAN M W. Evolution of antibiotic resistance by human and bacterial niche construction[J]. Evolution, 2005, 59(3): 477–491.
[29] DOEBELI M, ISPOLATOV I. Complexity and diversity[J]. Science, 2010, 328(5977): 494–497.
[30] HUI Cang. Adaptive diversification in coevolutionary systems[M]. Evolutionary Biology: Biodiversification from Genotype to Phenotype, Springer International Publishing, 2015.
[31] LALAND K N, ODLING-SMEE F J, FELDMAN M W. The evolutionary consequences of niche construction: A theoretical investigation using two-locus theory[J]. Journal of Evolutionary Biology, 1996, 9(3): 293–316.
[32] VANDERMEER J. The niche construction paradigm in ecological time[J]. Ecological Modelling, 2008, 214(2): 385–390.
[33] COHEN Y, GALIANO G. Evolutionary distributions and competition by way of reaction-diffusion and by way of convolution[J]. Bulletin of Mathematical Biology, 2013, 75(12): 2305–2323.
[34] HUI Cang, LI Zizhen, YUE Dongxia. Metapopulation dynamics and distribution, and environmental heterogeneity induced by niche construction[J]. Ecological Modelling, 2004, 177(1): 107–118.
[35] HAN Xiaozhuo, HUI Cang, ZHANG Yanyu. Effects of time-lagged niche construction on metapopulation dynamics and environmental heterogeneity[J]. Applied Mathematics and Computation, 2009, 215(2): 449–458.
[36] DOEBEL M, ISPOLATOV I. Diversity and coevolutionary dynamics in high-dimensional phenotype spaces[J]. American Naturalist, 2017, 189(2): 105–120.
[37] HUI Cang, MINOARIVELO H O, LANDI P, et al. Modelling coevolution in ecological networks with adaptive dynamics[J]. Mathematical Methods in the Applied Sciences, 2018, 41(18): 8407–8422.
[38] 韩晓卓, 覃姜色, 杨淑伶, 等. 一类进化分布动态模型及其应用[J]. 生态科学, 2017, 36(3): 135–141.
The effects of trait-mediated niche construction on evolutionary distribution dynamics of species with resource competition
LI Jianmei, HAN Xiaozhuo*
Guangdong University of Technology, School of Mathematics and Statistics , Guangzhou 510520, China
Niche construction refers to the process in which organisms regulate or influence their environments or other biological organisms through their own movement and metabolism, thus leading to different evolutionary trajectories of organisms in the process of natural selection. These conclusions were drawn based on the two-locus genetic models or mathematical ecological models that built with individual-based model (IBM) or cellular automata (CA) simulation methods. At present, the evolutionary distribution dynamics model is one of the popular methods to analyze the evolutionary invasion. By introducing this approach, we investigated the influence of trait-mediated niche construction on the evolution dynamics of species with intraspecific resource competition. The results confirmed that the niche construction mediated by traits caused obvious changes in the evolutionary trajectory of species. Comparing to the situation without the niche construction, the evolutionary branching reduced and the population density increased when the niche construction center was located in the small trait. While the niche construction acted on the large trait, it could maintain the evolutionary diversity, although the symmetrical evolutionary distribution of classical model was broken. It means that niche construction made significant differences in species traits through the long-term strong positive feedback with their environment. With the increasing of niche-constructing intensity, the species density of dominant traits increased significantly and the number of evolutionary branching decreased. Strong intraspecific competition and weak niche construction would be beneficial to maintain the evolutionary diversity of the species.
evolutionary branching; evolutionary diversification; niche construction; competition kernel; symmetrical distribution.
10.14108/j.cnki.1008-8873.2022.06.020
Q141
A
1008-8873(2022)06-167-09
2020-10-20;
2020-12-28
国家自然科学基金面上项目(31670391)
李建媚(1993—), 女, 广东广州人, 硕士研究生, 主要从事数学生态学研究, E-mail: 410275104@qq.com
通信作者:韩晓卓(1978—), 女, 博士, 教授, 主要从事数学生态学研究, E-mail: hanxzh@gdut.edu.cn
李建媚, 韩晓卓. 生态位构建对具有资源竞争物种的进化分布动态的影响[J]. 生态科学, 2022, 41(6): 167–175.
LI Jianmei, HAN Xiaozhuo. The effects of trait-mediated niche construction on evolutionary distribution dynamics of species with resource competition[J]. Ecological Science, 2022, 41(6): 167–175.
