饲粮类型对绒山羊瘤胃微生物数量的影响
2022-09-16杜海东韦玥瑞李科南张晓东娜梅拉娜仁花
■杜海东 韦玥瑞 李科南 张晓东 娜梅拉 娜仁花
(内蒙古农业大学动物科学学院,内蒙古呼和浩特 010018)
中国是世界上最大的绒山羊生产、加工以及销售国[1]。内蒙古白绒山羊作为我国著名的绒山羊品种,具有耐粗饲、适应性强、绒质好等优点,在内蒙古中西部荒漠、半荒漠地区被广泛养殖[2]。随着国家封山禁牧政策的实施,绒山羊的饲养方式已逐渐由放牧转为舍饲或半舍饲饲养。而在人工饲喂过程中,不同的饲料组成对绒山羊的生长、健康和产品质量有显著影响。
瘤胃是反刍动物饲料发酵、消化的重要场所,饲料中70%~85%的可消化物和50%的粗纤维在瘤胃内消化,定居于瘤胃的复杂微生物区系是瘤胃发挥发酵、消化功能的主要因素[3-4]。瘤胃微生物主要由细菌、古生菌、真菌和原虫共同组成,这些微生物通过将瘤胃内的饲料作为发酵基质产生代谢物包括挥发性脂肪酸(volatile fatty acids, VFA)、菌体蛋白、脂肪酸、二氧化碳和甲烷等[5-6]。其中VFA、菌体蛋白和脂肪酸等代谢产物不仅对瘤胃微生物的生长具有促进作用,还能够被宿主所吸收,对瘤胃的健康和动物的生长起到积极的作用,而二氧化碳和甲烷的产生降低了饲料能量的利用率,且两者作为温室气体排放到空气中,对环境也造成了消极影响[7-8]。研究发现,不同的饲料类型、组成形式、物理特性等对瘤胃微生物数量和发酵代谢产物有极大影响[9-10]。因此,本试验研究了不同饲粮类型对绒山羊瘤胃微生物数量的影响,从而筛选出最佳的饲粮类型,为优化绒山羊饲料配方提供科学依据。
1 材料与方法
1.1 试验动物与试验设计
试验选用24只体重相近的8月龄绒山羊母羊,随机分成4 个处理组,每组6 只母羊。经驱虫后进行试验,预试期20 d,正试期20 d。试验期间确保补饲时间、饲喂量、饲养条件相同,并保证自由饮水。4个处理组分别饲喂4种不同组成的饲粮,分别为苜蓿青干草(AH 组)、苜蓿青干草加补饲精料(AC 组)、玉米秸秆(CS组)和玉米秸秆加补饲精料(CC组)。试验日粮组成及营养水平见表1。
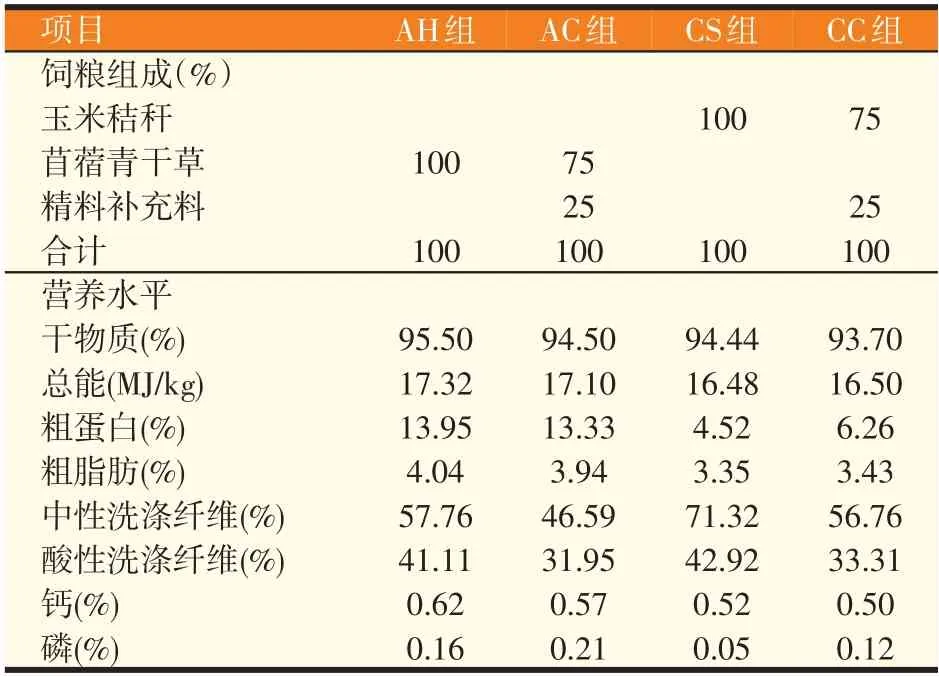
表1 饲粮组成及营养水平(风干基础)
1.2 瘤胃液的采集
在正试期的第5 天,试验羊空腹12 h 后,利用体外抽取法采集24 只母羊瘤胃液,采集的瘤胃液一部分经四层纱布过滤后分装于冻存管中放入液氮冷冻,置于-80 ℃冰箱中保存,用于微生物数量的测定。另取部分瘤胃液用双层纱布过滤,取滤液1 mL,加入甲基绿福尔马林染色液(由35%的福尔马林溶液100 mL,NaCl 8.0 g,甲基绿0.6 g 和纯化水900 mL 配制)2 mL,用于原虫数量的测定。
1.3 测试指标与方法
1.3.1 瘤胃微生物的提取和PCR扩增
参照韦玥瑞等[12]的方法提取瘤胃液中总DNA,将提取的DNA 在0.8%琼脂糖凝胶电泳检测,利用酶标仪测定样品DNA浓度和OD值,采用PCR技术对目的基因进行扩增。
1.3.2 质粒标准品的制备
利用PCR产物回收试剂盒(天根生化科技有限公司)将没有杂带的PCR 产物进行回收,然后将回收的DNA参照pMD19-T Simple Vector(TaKaRa)的克隆试剂盒说明书进行操作,挑选阳性克隆子测序鉴定,并在BLAST基因库进行在线对比,相似性达到90%以上的质粒用作标准质粒。
1.3.3 实时荧光定量PCR分析
对已知拷贝数的质粒标准品以10 倍梯度稀释,稀释的浓度梯度为4~6个,每个浓度梯度重复3次,进行PCR 反应制作相应微生物定量的标准曲线。对产甲烷菌、纤维分解菌和真菌进行实时定量PCR。绝对定量前将待测样品统一稀释到10 ng/μL,绝对定量后将测定样品的Ct值代入各自的标准曲线公式中,计算得到细菌拷贝数。实时定量PCR引物信息见表2。

表2 基因引物序列
1.3.4 原虫数量的测定
取瘤胃液后染色30 min,在光学显微镜下采用血球计数板计数法进行计算。
原虫数(个/mL)=X/S×D×1 000
式中:X——所观察的所有方格中原虫的总数(个);
S—计数的方格数;
D—稀释倍数。
1.4 数据统计与分析
试验数据经Excel整理后,采用SAS 9.2软件GLM模型进行单因素方差分析,组间均值进行Duncan's法多重比较,P<0.05表示差异显著。
2 结果与分析
2.1 荧光定量PCR的标准曲线结果(见表3)
由表3 可知,5 种瘤胃微生物标准曲线的相关系数值均大于0.99,且斜率为-3.8~-3.4。由以上数据得知,未知样均落于标准曲线上,各稀释梯度拷贝数与Ct值之间均存在良好线性关系,标准曲线斜率、相关系数与扩增效率均符合要求,可用于试验。
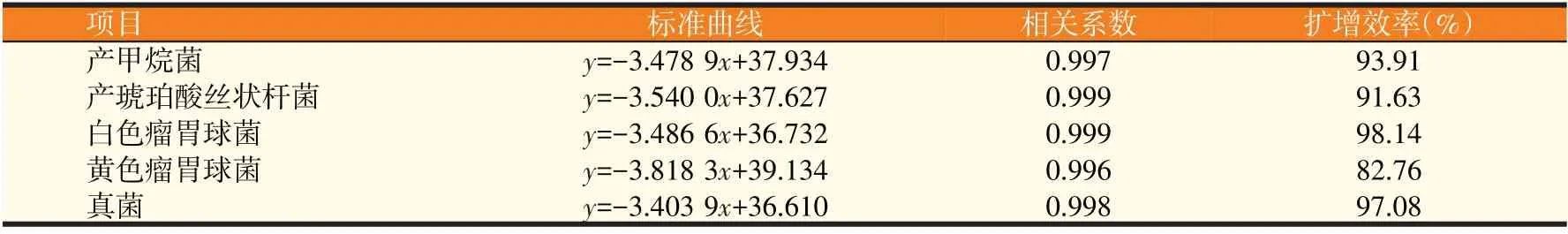
表3 瘤胃微生物标准曲线
2.2 不同饲粮对绒山羊瘤胃微生物数量的影响
2.2.1 不同饲粮对绒山羊瘤胃内产甲烷菌数量的影响(见表4)
由表4可知,饲喂不同饲粮瘤胃产甲烷菌数量表现为CS 组>CC 组>AH 组>AC 组,其中AC 组最少,为0.22×109拷贝数/g;CS 组最多,为3.30×109拷贝数/g。AC组产甲烷菌数量显著低于CS组与CC组(P<0.05),显著低于AH 组(P<0.05)。AH 组产甲烷菌数量显著低于CS与CC组(P<0.05)。CS组和CC组产甲烷菌数量无显著差异(P>0.05)。

表4 不同饲粮对产甲烷菌数量的影响(×109 拷贝数/g)
2.2.2 不同饲粮对绒山羊瘤胃内纤维分解菌数量的影响(见表5)
由表5可知,饲喂不同饲粮所产生的产琥珀酸丝状杆菌数量表现为CS组>CC组>AH组>AC组,其中AC组最少,为15.00×108拷贝数/g;CS组最多,为20.90×108拷贝数/g;AC组产琥珀酸丝状杆菌显著低于AH组、CS组和CC组(P<0.05)。AH组产琥珀酸丝状杆菌数量显著低于CS组与CC组(P<0.05)。CS组和CC组产琥珀酸丝状杆菌数量无显著差异。不同饲粮处理对绒山羊白色瘤胃球菌和黄色瘤胃球菌无显著影响(P>0.05)。

表5 不同饲粮对纤维分解菌数量的影响(×108 拷贝数/g)
2.2.3 不同饲粮对绒山羊瘤胃内真菌数量的影响(见表6)

表6 不同饲粮对真菌数量的影响(×106 拷贝数/g)
由表6得知,饲喂不同饲粮所产生的真菌数量表现为CS 组>AH 组>CC 组>AC 组,其中AC 组最少,为1.17×106拷贝数/g;CS 组最多,为6.35×106拷贝数/g;AC 组真菌数量低于AH 组、CC 组和CS 组,但差异不显著(P>0.05)。
2.2.4 不同饲粮对绒山羊瘤胃原虫数量的影响(见表7)
由表7可知,饲喂不同饲粮所产生的原虫数量表现为CS 组>CC 组>AH 组>AC 组,其中AC 组最少,为0.66×104个/mL;CS组最多,为4.02×104个/mL;AC组瘤胃原虫数量显著低于CS 组和CC 组(P<0.05),但AC组原虫数量与AH组无显著差异(P>0.05)。

表7 不同饲粮瘤胃原虫数量的比较(×104个/mL)
2.2.5 绒山羊瘤胃微生物相关性分析(见表8)
由表8 可知,显著性水平P<0.05,说明本次分析数据可靠,即研究样本可推广到整体。各研究变量之间的相关系数大部分都大于0.5,说明相关性较好。其中产琥珀酸丝状杆菌、产甲烷菌相关性最高,说明了这两个变量间的互相促进作用最强。
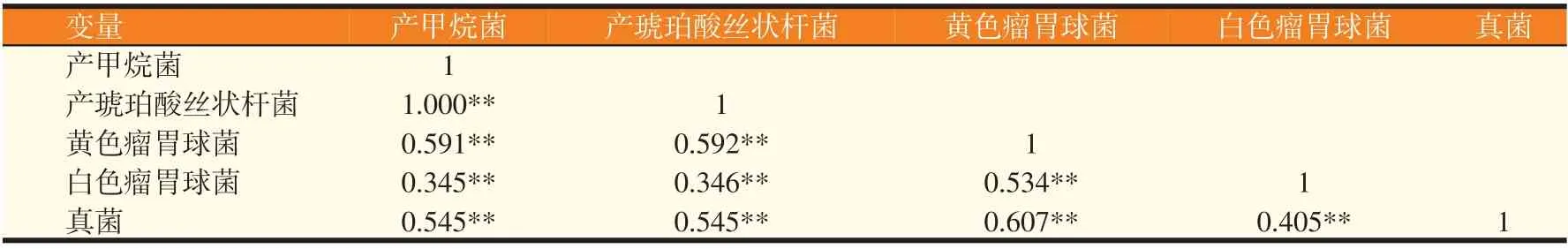
表8 不同微生物数量的相关性分析
3 讨论
3.1 饲粮类型对绒山羊瘤胃内产甲烷菌数量的影响
不同种类饲料的营养和生化特性能够影响瘤胃微生物种群和代谢,从而调控反刍动物优势菌群比例[13]。瘤胃中的产甲烷菌、纤维素分解菌、真菌和原虫的数量不仅可以反映机体对于粗饲料纤维消化水平的高低,同时也能间接反映机体对此类饲料的利用效率[14]。产甲烷菌利用瘤胃内的饲料作为发酵底物生成甲烷,这不仅降低了饲料的利用率,造成了能量的浪费,甲烷气体排放到空气中也造成了环境的污染。因此调控瘤胃产甲烷菌的数量是解决饲料能量浪费、降低甲烷排放的有效手段。侯路钊等[15]研究发现,荷斯坦公牛饲粮中的中性洗涤纤维/淀粉值降低能够减少甲烷排放量,降低瘤胃古菌丰富度指数和甲烷菌的数量。本研究也发现了相似的结果,苜蓿组与玉米秸秆组相比瘤胃甲烷菌的数量显著减少,而补饲精料进一步降低了瘤胃甲烷菌的数量,这可能是由于不同的饲粮组合能够通过影响瘤胃的发酵模式,进一步影响到瘤胃微生物的数量[16-17]。瘤胃产甲烷菌主要利用二氧化碳和氢气生成甲烷提供能量,高纤维结构性碳水化合物促使瘤胃乙酸生成量上升,乙酸生成过程会产生二氧化碳和氢气,为甲烷菌提供充足底物,而可消化碳水化合物促使瘤胃丙酸生成量上升,此过程消耗氢气[18]。本试验中,绒山羊饲喂玉米秸秆瘤胃产甲烷菌数量显著提高说明相对于优质干草和精料,玉米秸秆中木质素、纤维素含量高,其作为瘤胃甲烷菌发酵底物发酵程度更大。综上可知,甲烷菌数量的差异取决于饲料的类型与组成及其在瘤胃中的可用性,合理的饲料组成可减少产甲烷菌的数量。
3.2 饲粮类型对绒山羊瘤胃内纤维分解菌数量的影响
在反刍动物饲粮中纤维素是最普遍存在的,在瘤胃微生物中主要分解纤维素的菌包括产琥珀酸丝状杆菌、白色瘤胃球菌和黄色瘤胃球菌。各类纤维分解菌分泌的纤维二糖酶、木聚糖酶羧甲基、纤维素酶和果胶酶在纤维素降解过程中发挥重要作用[19]。纤维素分解菌的数量受饲粮结构的影响。并且,纤维素分解菌对不同物质的纤维素降解能力也存在差异[20]。董春晓等[21]研究不同饲料来源对胡羊瘤胃微生物组成的影响,结果发现相较于葵花籽壳和油菜秸秆组,玉米秸秆组胡羊瘤胃纤维素分解菌的数量显著提高。在本试验中饲喂玉米秸秆的绒山羊瘤胃中纤维分解菌数量大于饲喂苜蓿青干草的绒山羊,这与前人的研究结果一致。以上结果可能是因为高含量纤维饲料为细菌繁殖提供了充足的底物,从而增加了微生物的数量。此外,本研究中,绒山羊补饲精料瘤胃产琥珀酸丝状杆菌数量有所降低。Fernando 等[22]报道,瘤胃微生物群落的变化与饲粮的精粗比有关,当饲粮的精粗比从40∶60提高到60∶40时瘤胃微生物的组成也随之变换,并且,在以粗料为主的饲粮中纤维素分解菌的数量更多。杨硕等[23]也发现相较于放牧组山羊,放牧加补饲组山羊瘤胃纤维素分解菌数量显著降低。这可能是由于瘤胃微生物在分解精料中碳水化合物过程中,有机酸含量显著升高,降低瘤胃的pH并形成酸性环境抑制瘤胃纤维分解菌的生长[24]。
3.3 饲粮类型对绒山羊瘤胃内真菌数量的影响
瘤胃中的真菌占瘤胃微生物总量的5%~20%,在纤维消化中起着重要作用,由于其复杂而丰富的酶活性在消化结构性碳水化合物方面可能比细菌具有竞争优势。真菌能分泌产生纤维素酶,对植物纤维(纤维素、木质素和半纤维素)具有很强的消化能力,此外,真菌的穿透生长同时可以使植物组织纤维内部张力减小变得疏松而易于降解[19]。在本试验中,玉米秸秆组绒山羊瘤胃真菌的数量大于苜蓿青干草组和补饲精料组,且补饲精料组真菌数量在数值上明显下降。付子琳等[25]研究发现放牧组与舍饲组的成年滩羊瘤胃真菌多样性差异不显著,但两组间瘤胃菌群结构存在一定差异,相较于舍饲组,放牧组滩羊瘤胃真菌丰富度显著提高。杨宏波等[26]也发现,饲喂高精料饲料的犊牛瘤胃厌氧真菌的活性和数量会下降。孙云章等[27]发现,随着体外发酵时间的增加,精粗比低的组瘤胃真菌的数量有大幅度的上升,而随着精料的提高真菌数量显著下降甚至消失。以上结果表明,瘤胃中的厌氧真菌对底物的选择具有偏好性,真菌更偏好于附着在纤维素上,含较高纤维素的饲料更有利于厌氧真菌生长[28]。
3.4 饲粮类型对绒山羊瘤胃内原虫数量的影响
原虫在瘤胃大量存在,据估计每克瘤胃液原虫数量为105~106个,占瘤胃微生物总量的30%以上[29]。瘤胃中大约62%的纤维素降解与原虫有关。原虫的种群平衡对维持良好的瘤胃环境和其他微生物发酵功能至关重要。瘤胃原虫能够将可溶性糖和淀粉合成储备碳水化合物,且原虫对碳水化合物发酵的速度低于细菌,这预防了其他微生物快速发酵生成短链脂肪酸(SCFAs),从而避免了瘤胃pH 的下降,降低了动物酸中毒的风险,增加了饲料在瘤胃的留存时间[30]。此外,瘤胃原虫还是重要的氢生产者,瘤胃9%~25%的甲烷产生与其有关[31]。因此,合理调控瘤胃原虫的数量对反刍动物的健康和能量消耗具有重大意义。在本研究中,苜蓿组和玉米秸秆组绒山羊在补充精料后瘤胃中原虫数量均不同程度减少。Franzolin等[32]研究发现,相较于饲料喂养水牛,放牧组水牛瘤胃的原虫组成有显著变化,并且放牧组纤毛虫群落总量显著高于饲料组。杨硕等[23]研究发现对放牧绒山羊补饲精料后瘤胃中原虫数量显著下降。这与本研究结果一致。以上结果表明,原虫的数量受饲粮类型和组成的影响。因此,在生产中充分利用饲料组合技术可以有效地调节瘤胃微生物的数量。此外,本试验中饲喂苜蓿的绒山羊瘤胃中原虫数量低于饲喂玉米秸秆的绒山羊,这可能是因为苜蓿中含有丰富的皂苷对原虫的生长具有抑制作用[33]。
在瘤胃中,产甲烷菌、纤维素分解菌、真菌、原虫等各类微生物互相作用,相互影响,共同构成一个错综复杂的瘤胃微生物系统。纤维素分解菌、真菌、原虫等在利用瘤胃饲料的同时产生氢和二氧化碳,随着发酵的进行,氢和二氧化碳的产量逐渐积累增加发生负反馈调节,对纤维素的降解发挥抑制作用,而产甲烷菌利用氢和二氧化碳生成甲烷,提高纤维素利用率[34]。有研究表明,甲烷产量与纤维的降解率呈正相关,并且反刍动物瘤胃中产甲烷菌的数量和纤维降解菌的数量也呈正相关关系[35]。Wei等[36]研究发现,瘤胃甲烷菌和真菌体外共同培养对纤维素的降解能力优于仅真菌培养组。赵小蓉等[20]将纤维素分解菌体外培养研究其对纤维素物质的分解能力,结果发现,当真菌和纤维素分解菌共同接种时,对纤维素的降解速度优于单一菌株。本试验通过数据分析各变量之间的相关系数大部分都大于0.50,说明了不同微生物之间相关性较好,这与前人的研究结果相似。以上结果表明,不同种类的瘤胃微生物在对纤维素的分解利用过程中存在互作作用。
4 结论
本试验结果表明,饲喂苜蓿青干草的绒山羊,瘤胃内产甲烷菌和原虫的数量显著降低,因此,优质牧草有利于减少甲烷菌和原虫数量,并且补饲精料后会进一步降低甲烷菌和原虫的数量,本试验结果可为生产中的饲料组合技术提供科学合理的数据,同时也能为下一步优化反刍动物饲料结构提供参考。
