基于陆地棉种质资源表型性状的遗传多样性分析
2022-09-15李有忠赵曾强王志军董永梅谢宗铭
李有忠,赵曾强,王志军,董永梅,谢宗铭*
基于陆地棉种质资源表型性状的遗传多样性分析
李有忠1,2,赵曾强1,2,王志军1,2,董永梅1,2,谢宗铭1,2*
(1.新疆农垦科学院生物技术研究所,新疆 石河子 832000;2.作物种质创新与基因资源利用兵团重点实验室,新疆 石河子 832000)
对收集的630份陆地棉种质资源的表型性状进行遗传多样性、相关性、主成分和聚类分析。结果表明:16个性状中有效铃数、果枝始节位的变异系数最大;各性状的遗传多样性较高,平均遗传多样性指数为2.11;纤维上半部平均长度与纤维比强度、纤维整齐度呈显著正相关关系,生育期与果枝始节位、纤维伸长率呈显著正相关关系;棉花表型性状中前5个主成分的特征值均大于1,累计反映了总信息量的74.798%;第1主成分的特征值为4.536,贡献率为28.353%,纺纱一致性指数、纤维整齐度、纤维上半部平均长度的特征向量值较大;第2主成分的特征值为2.574,贡献率为16.088%,纤维成熟度、马克隆值的特征向量值较大;第3主成分的特征值为1.941,贡献率为12.133%,果枝数、有效铃数、果枝始节位的特征向量值较大;聚类分析将630份棉花种质材料分成了6大类,其中第II类包含2份衣分低、纤维品质差的材料(佳荣棉7、三江大花);第III类包含10份纤维上半部平均长度较长、纤维比强度较大的材料(东兰花、南丹、平阳棉、里湖1、里当–陆5、罗甸铁籽、平塘棉、榕江棉、三都棉、开棉),可作为改良棉花纤维比强度、产量构成及纤维品质的材料加以利用。
陆地棉;种质资源;表型性状;遗传多样性分析;主成分分析;聚类分析
棉花是重要的经济作物,是纺织工业天然纤维的主要来源,同时也是食用油和植物蛋白的重要来源[1]。随着中国棉花产业战略西移,新疆已经成为中国最大的产棉区。根据国家统计公报[2],2021年新疆棉花播种面积2.506 1×106hm2,皮棉产量5.129×106t,分别占全国的78.9%和87.3%。毫无疑问,棉花产业已经成为新疆经济的重要支柱和棉区农民收入的主要来源。但随着棉花骨干亲本的集中使用以及转基因技术的应用,培育出的棉花新品种种间的遗传差异越来越小[3],导致棉花品质和产量难以获得更大突破。
优良的棉花品种是棉花生产的先决条件,种质资源是研究棉花遗传性状、发掘优异等位变异基因位点及培育新品种的重要基础[4];因此,开展棉花种质资源多样性研究,了解种质间的亲缘关系和遗传组成对优异种质的筛选、资源的合理高效利用和品种改良具有重要意义。有学者利用变异分析、相关性分析、聚类分析和主成分分析等统计方法对棉花的产量和纤维品质[5–8]、早熟性[9–11]、抗病性[12–13]、抗旱耐盐性[14–17]等进行了大量研究。孔清泉等[18]对92份陆地棉种质资源的8个性状进行了遗传变异、相关性和聚类分析,发现92份材料的成铃数和单铃质量的差异较大,资源类型丰富,有利于种质材料的比较和筛选。董承光等[19]运用相关性、主成分及聚类分析对429份陆地棉种质资源性状进行综合评价,指出陆地棉品种选育应集中在纤维品质优良、高衣分和单株铃数多的品种。邓艳凤等[20]以自主选育的57份棉花品系材料为研究对象,对其12个农艺性状进行遗传变异、相关性和聚类分析,通过综合评价筛选出5份早熟高产优质棉材料。赵云雷等[21]对125份陆地棉优异种质进行抗病性鉴定,共筛选到40个抗黄萎病优异等位基因。刘娜等[22]运用主成分分析和聚类分析等综合评价分析了319份陆地棉材料的遗传多样性,筛选出可用于耐盐育种的耐盐性资源材料。郑巨云等[23]运用抗旱综合指数、抗旱综合度量值等评价方法,结合灰色关联度及聚类分析,对272份棉花品种资源进行抗旱性综合评价,筛选出4份强抗旱、24份抗旱、134份耐旱、100份较敏旱及10份敏旱材料,为棉花抗旱遗传改良提供了特异资源。为更有效地利用棉花种质资源丰富的遗传背景,构建高效可育的棉花选育体系,本研究中,笔者对国内外630份陆地棉种质资源进行了农艺性状、品质性状指标的调查研究,通过分析其遗传多样性,旨在揭示其表型规律,筛选相对合理的表型评价指标,为棉花特异种质资源的创制和优质新品种的选育和生产提供亲本材料和理论依据。
1 材料与方法
1.1 供试材料
供试材料为新疆农垦科学院生物技术研究所多年来收集的国内外630份陆地棉种质资源。其中江苏省59份、河北省80份、山东省59份、湖南省11份、山西省6份、贵州省31份、甘肃省1份、河南省92份、广西壮族自治区32份、辽宁省8份、上海市2份、四川省14份、安徽省8份、湖北省63份、云南省4份、江西省3份、浙江省17份、陕西省9份、福建省1份、台湾省1份、新疆维吾尔自治区115份;国外种质14份。
1.2 方法
试验于2018—2019年在新疆农垦科学院生物技术研究所试验基地进行。完全随机区组排列。重复播种2年,每年2次重复。试验区肥力中等,均匀一致。小区面积9.12 m2,行长4 m,株距12 cm。人工点播,膜下水肥一体化滴灌栽培,常规大田管理。
调查各材料的出苗期及吐絮期,了解各材料的生育期;参照棉花种质资源描述规范和数据标准[24]进行性状调查。在棉花成铃后期,选取行中间长势均匀一致的10株材料调查株高、果枝始节高、果枝始节位、果枝数、单株铃数。吐絮后期,每小区收集中部20朵花,室内轧花考种(单铃质量、衣分);每份材料称取皮棉约10~12 g,采用印度普瑞美HFT9000纤维检测仪检测纤维品质,主要考察纤维上半部平均长度、纤维比强度、马克隆值、纤维整齐度及纤维伸长率。
1.3 数据处理与分析
采用SPSS 25.0计算各性状的平均值、标准差及变异系数等;根据平均数()和标准差()将每个性状分为10级,第1级≤–2,第10级>+2,中间每级相差0.5[5, 25];运用R 4.0.1软件ggplot2[26]及ggtree[27]绘制主成分分析图和聚类分析图;采用非加权类平均法[28]进行聚类分析,种质间距离为欧式距离[29]。表型性状的遗传多样性指数采用Shannon’s 信息多样性指数()进行分析。
–∑PlnP (1)
式中,P表示某一性状第级别出现的概率。
2 结果与分析
2.1 陆地棉种质资源表型性状的遗传多样性
630份陆地棉种质资源材料的变异分析结果(表1)表明,种质资源不同性状间的变异范围为1.92%~ 25.22%。其中,有效铃数的变异系数最大,为25.55%,变异范围为4.50~24.50个;纤维整齐度的变异系数最小,为1.92%,变异范围为76.15%~ 90.85%;果枝始节位、短纤维率、果枝始节高、果枝数、衣分、纤维伸长率、单铃质量、株高、纺纱一致性指数、马克隆值、纤维比强度、纤维上半部平均长度、生育期、纤维成熟度的变异系数依次减小。16个性状的遗传多样性指数最高的是生育期,其多样性指数为3.89;最低的是有效铃数,其多样性指数为1.63,平均遗传多样性指数为2.11。综上可以看出630份陆地棉种质资源的遗传性状差异较大,表现出较高的遗传多样性。
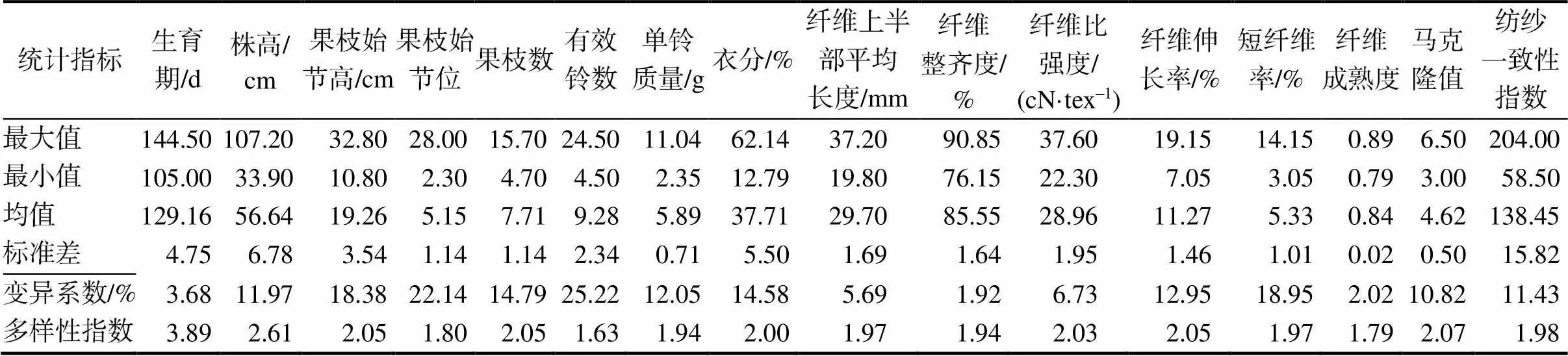
表1 630份陆地棉种质资源的表型性状
2.2 陆地棉种质资源表型性状的相关性分析
对630份陆地棉种质资源的16个表型性状进行相关性分析,结果(表2)表明:生育期与果枝始节位、纤维伸长率呈极显著正相关;株高与果枝始节高、果枝数、有效铃数、纤维整齐度、纤维比强度、纤维成熟度、马克隆值呈极显著正相关,与短纤维率呈极显著负相关;果枝始节高与果枝始节位、单铃质量、衣分、纤维上半部平均长度、纤维整齐度、纤维比强度、纤维成熟度、马克隆值、纺纱一致性指数呈极显著正相关,与果枝数、有效铃数、短纤维率呈极显著负相关;果枝始节位与果枝数、有效铃数呈极显著负相关;果枝数与有效铃数呈极显著正相关,与单铃质量、纤维上半部平均长度、纺纱一致性指数呈极显著负相关;有效铃数与单铃质量、纤维上半部平均长度、纤维比强度、纺纱一致性指数呈极显著负相关;单铃质量与衣分、纤维上半部平均长度、纤维整齐度、纤维比强度、纺纱一致性指数呈极显著正相关,与短纤维率呈极显著负相关;衣分与纤维上半部平均长度、纤维整齐度、纤维比强度、纤维成熟度、马克隆值、纺纱一致性指数呈极显著正相关,与短纤维率呈极显著负相关;纤维上半部平均长度与纤维整齐度、纤维比强度、纺纱一致性指数呈极显著正相关,与短纤维率、纤维成熟度、马克隆值呈极显著负相关;纤维整齐度与纤维比强度、纺纱一致性指数呈极显著正相关,与短纤维率、马克隆值呈极显著负相关;纤维比强度与纤维成熟度、纺纱一致性指数呈极显著正相关,与纤维伸长率、短纤维率呈极显著负相关;纤维伸长率与纤维成熟度、马克隆值、纺纱一致性指数呈极显著负相关;短纤维率与纺纱一致性指数呈极显著负相关;纤维成熟度与马克隆值呈极显著正相关,与纺纱一致性指数呈极显著负相关;马克隆值与纺纱一致性指数呈极显著负相关。从相关性分析可以看出陆地棉各性状间的相关关系比较复杂,相互制约。
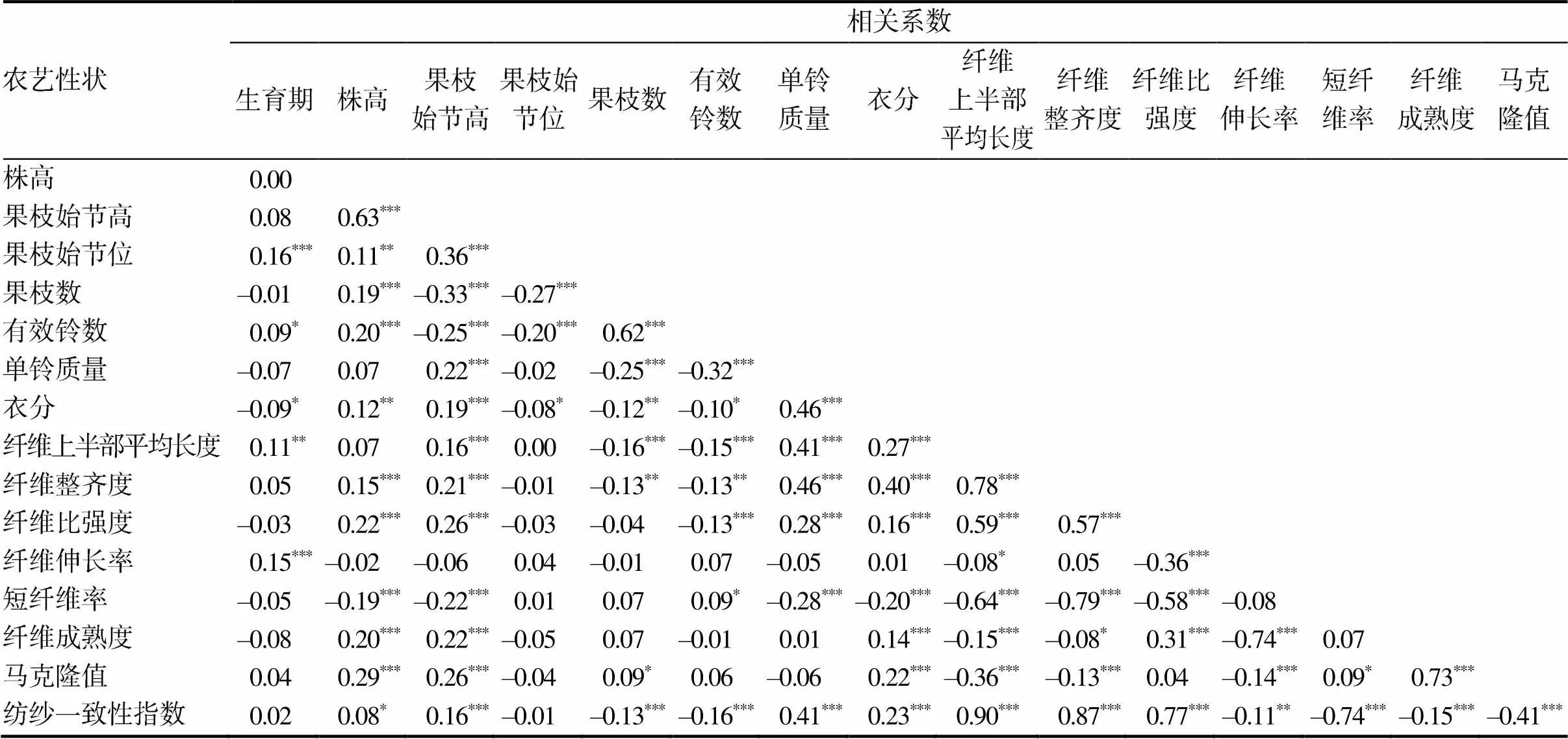
表2 630份陆地棉种质资源各农艺性状间的相关系数
“***”“**”“ *”分别表示在0.001、0.01及0.05水平下相关性显著。
2.3 陆地棉种质资源表型性状的主成分分析
2.3.1主成分个数的选取及因子载荷矩阵分析
对16个性状进行主成分分析,以特征值大于1为标准提取主成分。由图1可知,棉花表型性状中前5个主成分的特征值均大于1,说明这5个主成分在棉花表型变异构成中作用较大,累计反映了总信息量的74.798%。从表3可以看出,第1主成分的特征值为4.536,贡献率为28.353%,第1主成分中特征向量值最大的是纺纱一致性指数(0.440),说明纺纱一致性指数对第1主成分的影响最大;其后依次是纤维整齐度(0.423)、纤维上半部平均长度(0.409)。第2主成分的特征值为2.574,贡献率为16.088%,第2主成分中特征向量最大的是纤维成熟度(–0.569),其次是马克隆值(–0.500)。第3主成分的特征值为1.941,贡献率为12.133%。第3主成分中特征向量值最大的是果枝数(0.574),其次是有效铃数(0.520)。第4主成分的特征值为1.618,贡献率为10.110%,特征向量值最大的为株高(0.510),其次为纤维伸长率(0.434)。第5主成分的特征值1.298,贡献率为8.114%,特征向量最大的为纤维上半部平均长度(–0.613),其次是单铃质量(–0.391)。
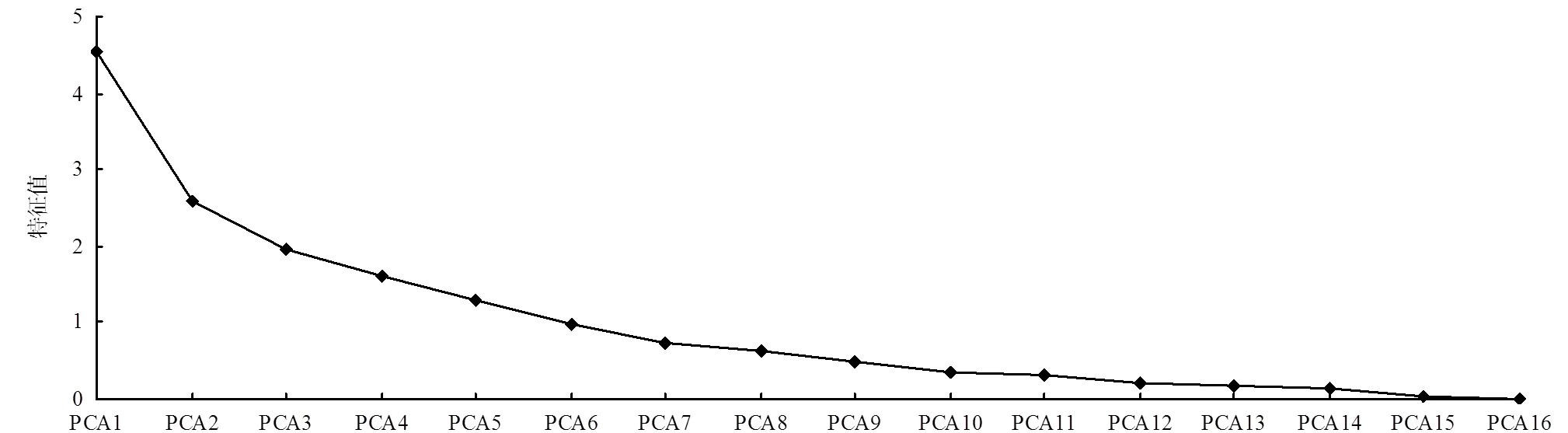
主成分
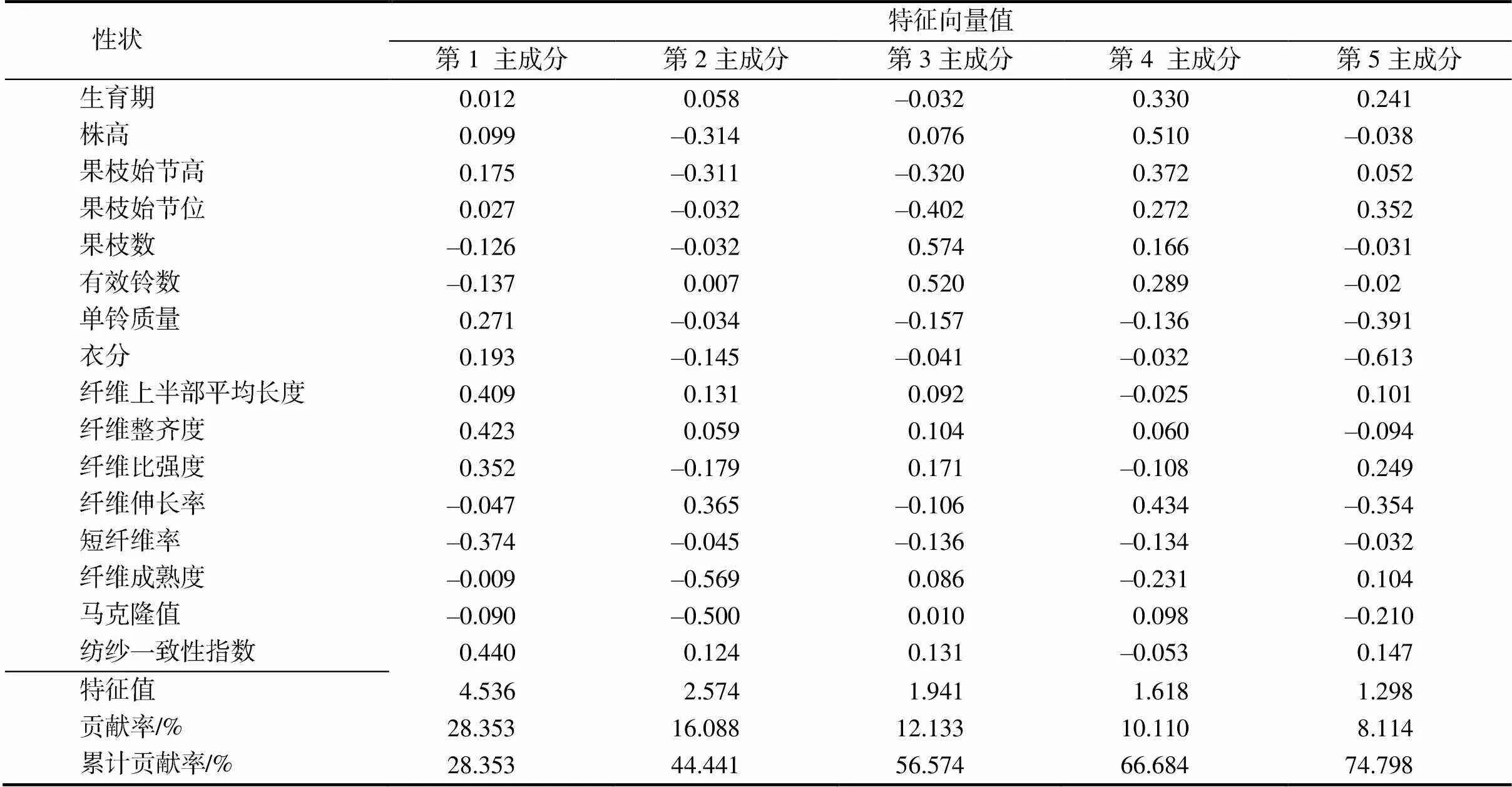
表3 棉花表型性状的主成分矩阵与特征向量和贡献率
2.3.2 陆地棉种质资源主要性状之间的主成分分析
对630份陆地棉种质资源的16个表型性状进行主成分分析,结果如图2所示。其中,横、纵坐标是主成分,各个向量在主成分上的投影可以代表两者的相关程度;不同种质资源的位置和距离不同,距离的远近反映品种之间的差异大小,而品种在单个向量上的投影表示在该属性向量上的表现。从图2可以看出,与第1主成分密切相关的为纤维品质性状,与第2主成分密切相关的性状为纤维成熟度。630份种质资源在图上的分布比较分散,表明630份种质资源间农艺性状及品质性状具有较大的差异,品种之间的相似性较小。
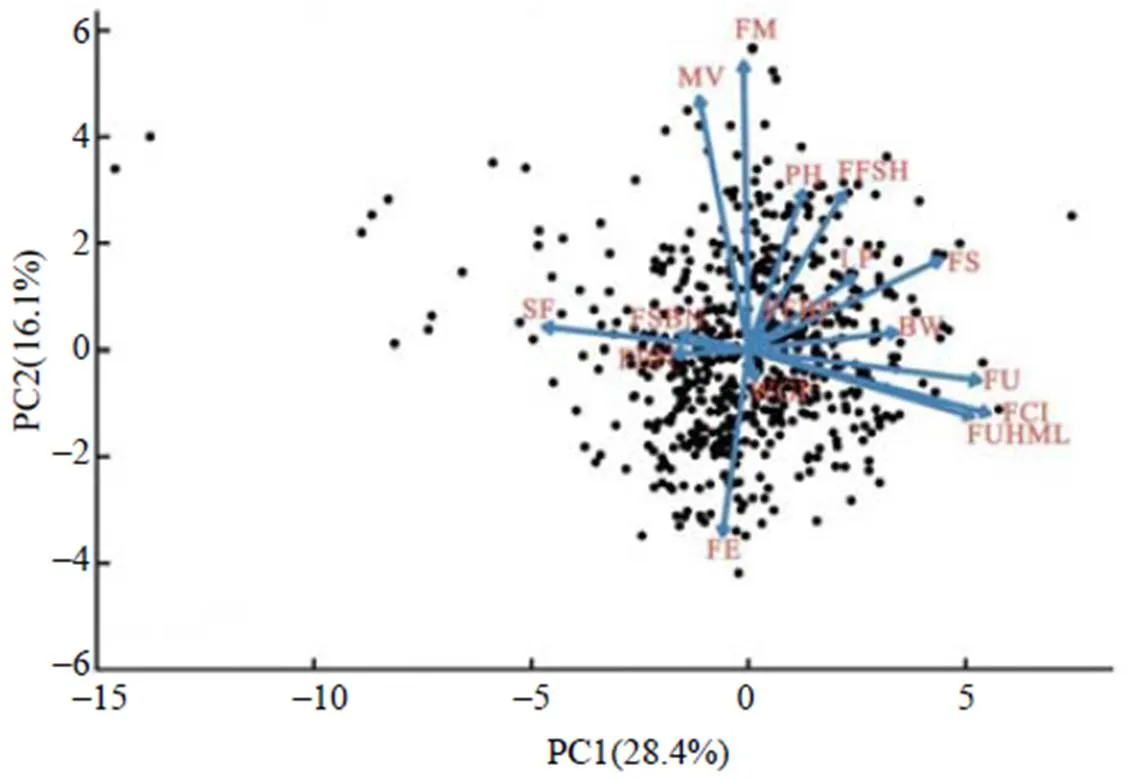
WGP示生育期;PH示株高;FFSH示果枝始节高;FFBP示果枝始节位;FSBN示果枝数;FBN示有效铃数;BW示单铃质量;LP示衣分;FUHML示纤维上半部平均长度;FU示纤维整齐度;FS示纤维比强度;FE示纤维伸长率;SF示短纤维率;FM示纤维成熟度;MV示马克隆值;FCI示纺纱一致性指数。
2.4 陆地棉种质资源表型性状的聚类分析
以欧式距离为遗传距离,采用类平均法对表型性状数据进行聚类分析。从图3可以看出,630份种质资源可以分为6大类群,第I类包含1份材料,为贵州的那佐–陆材料,属于单铃质量大(6.5 g)的材料;第II类包含2份材料(佳荣棉7,三江大花),均为贵州的材料,属于衣分低、纤维品质差的材料,纤维上半部平均长度只有20 mm,衣分为10%左右;第III类包含10份材料(东兰花、南丹、平阳棉、里湖1、里当–陆5、罗甸铁籽、平塘棉、榕江棉、三都棉、开棉),属于纤维上半部平均长度、纤维强度较好的材料,可作为改良棉花纤维比强度、产量构成及纤维品质的材料加以利用。第IV类包含57份材料,属于生育期较长的材料(130 d),其中有11份来源于贵州,10份来源于广西,还有1份来源于美国,大部分来源于华南棉区;第V类包含3份材料,属于衣分较高,纤维品质较差的材料;第VI类包含557份材料,属于铃数、纤维上半部平均长度、纤维比强度适中的材料。

图3 630份陆地棉种质资源聚类结果
3 结论及讨论
深入研究作物种质资源的遗传多样性,在作物遗传育种的环节中具有重要意义[30–31]。变异系数是体现表型性状多样性程度的指标,变异系数越大,性状间个体差异越大,一般变异系数大于10%表示样本间差异较大[32]。本研究中,11个性状的变异系数均大于10%,其中,有效铃数和果枝始节位的变异系数最大。说明630份种质资源间的差异大,资源类型丰富,有利于特异种质材料的比较和筛选。
研究数量性状之间的相关性对评价2种或2种以上性状联合选择的可行性具有重要意义。本研究对630份陆地棉种质资源的16个数量性状进行了相关性分析,结果显示株高与果枝数呈极显著正相关,这与李慧琴等[8]的研究结果一致。但大多数产量性状与品质性状呈显著负相关,表明同步改良棉花品质及产量性状有一定的难度。
对不同种质进行分类,可以针对不同的育种目标进行定向选育,在遗传改良中优势互补,从而有效指导作物的遗传育种。本研究对16个陆地棉性状进行聚类分析,将630份种质资源分为6个类群,筛选出2份产量低、品质差的材料,10份纤维上半部平均长度、纤维比强度大的材料(东兰花,南丹,平阳棉,里湖1,里当–陆5,罗甸铁籽,平塘棉,榕江棉,三都棉,开棉)。本研究拟对这10份品质较好的材料进一步进行田间棉花育种应用,以期选育出高产、优质的棉花新品种。
[1] CAMPBELL B T,SAHA S,PERCY R,et al.Status of the global cotton germplasm resources[J].Crop Science,2010,50(4):1161–1179.
[2] 国家统计局.国家统计局关于2021年棉花产量的公告[N].中国信息报,2021–12–15(1).
[3] 匡猛,杨伟华,许红霞,等.中国棉花主栽品种DNA指纹图谱构建及SSR标记遗传多样性分析[J].中国农业科学,2011,44(1):20–27.
[4] ESBROECK G V,BOWMAN D T.Cotton germplasm diversity and its importance to cultivar development[J]. The Journal of Cotton Science,1998,2:121–129.
[5] 王林辉,余正军,刘少卿,等.27个棉花育种材料的产量及部分纤维品质差异分析[J].湖南农业大学学报(自然科学版),2009,35(1):9–12.
[6] 代攀虹,孙君灵,何守朴,等.陆地棉核心种质表型性状遗传多样性分析及综合评价[J].中国农业科学,2016,49(19):3694–3708.
[7] 钱玉源,刘祎,崔淑芳,等.基于表型的棉花种质资源遗传多样性分析及核心种质的抽提[J].华北农学报,2019,34(S1):29–35.
[8] 李慧琴,崔建强,王潭刚,等.24份陆地棉材料表型性状遗传多样性分析[J].新疆农垦科技,2019,42(5):10–13.
[9] 喻树迅,王寒涛,魏恒玲,等.棉花早熟性研究进展及其应用[J].棉花学报,2017,29(S1):1–10.
[10] 高进,蔡立旺,宋锦花,等.早熟棉花品种产量品质性状的综合评价与聚类分析[J].西南农业学报,2015,28(6):2425–2431.
[11] 宿俊吉.陆地棉早熟与产量纤维品质性状的全基因组关联分析及候选基因筛选[D].杨凌:西北农林科技大学,2017.
[12] 赵俊兴,徐小鸿,吴静,等.2009—2015年陕西棉花品种抗病性鉴定及分析[J].西北农业学报,2016,25(12):1816–1820.
[13] 赵沛,杨兆光,吴洁,等.近十年来参加江西省棉花区试品种的抗病性分析[J].棉花科学,2021,43(1):37–42.
[14] 郑巨云,曾辉,王俊铎,等.陆地棉品种资源萌发期耐盐性的隶属函数法评价[J].新疆农业科学,2018,55(9):1579–1592.
[15] 韩贝,王旭文,李保奇,等.陆地棉种质资源抗旱性状的关联分析[J].作物学报,2021,47(3):438–450.
[16] 李菲,龚记熠,李欲柯,等.抗旱耐盐植物功能基因发掘及其在棉花育种中的应用[J].分子植物育种,2019,17(22):7395–7400.
[17] 郑巨云,曾辉,王俊铎,等.陆地棉品种资源盛蕾期抗旱性状的综合评价[J].分子植物育种,2021,19(19):6512–6528.
[18] 孔清泉,杨兆光,吴振江,等.陆地棉92份种质资源主要性状的遗传多样性分析[J].棉花科学,2020,42(3):33–40.
[19] 董承光,王娟,周小凤,等.基于表型性状的陆地棉种质资源遗传多样性分析[J].植物遗传资源学报,2016,17(3):438–446.
[20] 邓艳凤,肖水平,柯兴盛,等.长江流域早熟棉新品系主要产量、品质及农艺性状的分析[J].江西农业大学学报,2019,41(5):861–872.
[21] 赵云雷,王红梅,陈伟,等.基于优异等位基因的棉花抗黄萎病性状的分子鉴定[J].中国农业科学,2017,50(2):216–227.
[22] 刘娜,高文伟.陆地棉耐盐表型数据的遗传多样性研究[J].种子,2019,38(11):57–62.
[23] 郑巨云,王俊铎,龚照龙,等.棉花品种资源花铃期抗旱性鉴定与评价[J].干旱地区农业研究,2021,39(2):150–163.
[24] 杜雄明,周忠丽.棉花种质资源描述规范和数据标准[M].北京:中国农业出版社,2005.
[25] 周瑜,李泽碧,黄娟,等.高粱种质资源表型性状的遗传多样性分析[J].植物遗传资源学报,2021,22(3):654–664.
[26] ITO K,MURPHY D.Application ofto pharmacometric graphics[J].CPT:Pharmacometrics & Systems Pharmacology,2013,2(10):e79.
[27] YU G C,SMITH D K,ZHU H C,et al.Ggtree:an R package for visualization and annotation of phylogenetic trees with their covariates and other associated data[J]. Methods in Ecology and Evolution,2017,8(1):28–36.
[28] HUANG C Q,LONG T,BAI C J,et al.Establishment of a core collection ofbased on morphological data[J].Tropical Grasslands-Forrajes Tropicales,2020,8(3):203–213.
[29] WANG J C,HU J,XU H M,et al.A strategy on constructing core collections by least distance stepwise sampling[J].Theoretical and Applied Genetics,2007,115(1):1–8.
[30] 杨学乐,何录秋,张璐,等.湖南芝麻种质资源农艺性状的遗传多样性[J].湖南农业大学学报(自然科学版),2019,45(6):577–582.
[31] 彭嘉熹,周仲华,马肖,等.陆地棉主推品种种质资源的评价与分析[J].湖南农业大学学报(自然科学版),2021,47(1):23–29.
[32] 李慧琴,于娅,王鹏,等.270份陆地棉种质资源农艺性状与品质性状的遗传多样性分析[J].植物遗传资源学报,2019,20(4):903–910.
Phenotypic traits genetic diversity analysis of upland cotton germplasm resources
LI Youzhong1,2,ZHAO Zengqiang1,2,WANG Zhijun1,2,DONG Yongmei1,2,XIE Zongming1,2*
(1.Biotechnology Research Institute, Xinjiang Academy of Agricultural Reclamation Sciences, Shihezi, Xinjiang 832000, China; 2.Xinjiang Production & Construction Group Key Laboratory of Crop Germplasm Enhancement and Gene Resources Utilization, Shihezi, Xinjiang 832000, China)
In this study, genetic diversity, trait correlation, principal component and cluster analysis were performed on 630 upland cotton germplasm resources. The results showed that among the 16 traits, the effective boll number and the starting node of the fruit branch had the largest variation coefficient; the genetic diversity of each trait was higher, with an average of 2.11; the correlation analysis showed that the average length of the upper half of the fiber was correlated with the specific strength of the fiber. Fiber uniformity showed a significant positive correlation and the growth period, fruiting branch start node position and fiber elongation also showed a significant positive correlation. The principal component analysis showed that the eigenvalues of the first five principal components of cotton phenotypic traits were all greater than 1, which cumulatively reflected 74.798 of the total information. The eigenvalue of the first principal component was 4.536, the contribution rate was 28.353%, and the eigenvector values of the spinningconsistency index, fiber uniformity and fiber upper half mean length were larger.The second principal component was 2.574, the contribution rate was 16.088%, and the eigenvector values of the fiber maturity andmicronaire value were larger.The third principal component was 1.941, the contribution rate was 12.133%, and the eigenvector values of the fruit branch number, effective boll number and first fruit branch position were larger. The cluster analysis divided the 630 cotton germplasm materials into 6 categories, of which category II contained 2 materials(Jiarong cotton 7, Sanjiang Dahua), which were the materials owing low lint content and poor fiber quality; the Category III contained 10 materials(Donglanhua, Nandan, Pingyang cotton, Lihu 1, Lidang-Lu 5, Luodiantiezi, Pingtang cotton, Rongjiang cotton, Sandu cotton, Kaimian), which belonged to the materials having the longest average length of the upper half of the fiber and the highest fiber specific strength, which could be used as materials for improving cotton fiber specific strength, output composition and fiber quality.
upland cotton; germplasm resources; phenotypic traits; genetic diversity analysis; principal component analysis; cluster analysis
S562.024
A
1007-1032(2022)04-0394-07
李有忠,赵曾强,王志军,董永梅,谢宗铭.基于陆地棉种质资源表型性状的遗传多样性分析[J].湖南农业大学学报(自然科学版),2022,48(4):394–400.
LI Y Z,ZHAO Z Q,WANG Z J,DONG Y M,XIE Z M.Phenotypic traits genetic diversity analysis of upland cotton germplasm resources[J].Journal of Hunan Agricultural University(Natural Sciences),2022,48(4):394–400.
http://xb.hunau.edu.cn

2021–10–24
2022–07–21
国家重点研发计划项目(2016YFD0100203–6);八师石河子市中青年科技创新领军人才计划项目(2020RC06);兵团农业科技创新工程专项(NCG202224);国家科技资源共享服务平台项目(NCGRC–2022–051)
李有忠(1981—),男,甘肃武威人,硕士,副研究员,主要从事棉花抗逆种质资源创制与应用研究,2529629871@qq.com;*通信作者,谢宗铭,博士,研究员,主要从事棉花遗传育种研究,xiezmchy@163.com
责任编辑:毛友纯
英文编辑:柳正
