独脚金内酯与激素互作调控根系生长的研究进展
2022-09-14沈月陶宝杰华夏吕冰刘立军陈云
沈月 陶宝杰 华夏 吕冰 刘立军 陈云
(1. 江苏省作物遗传生理重点实验室,扬州 225009;2. 扬州大学生物科学与技术学院,扬州 225009)
根系作为植物的重要器官,不仅能固定植株,还具有从土壤中吸收、贮藏与运输水分和养分,以及合成多种有机化合物等功能。植物根系生长发育(包括主根的伸长、侧根的发生、不定根的形成等)受到多种植物激素、外界环境信号和遗传物质的调控。植物激素是植物自身微量合成且能调节其生理过程的有机化合物的总称,主要包括生长素(indole-3-acetic acid,IAA)、细胞分裂素(cytokinin,CK)、赤霉素(gibberellin,GA)、脱落酸(abscisic acid,ABA)和乙烯(ethylene,ETH)。独脚金内酯(strigolactone,SL)是一种在植物根部合成的新型植物激素,调控植物的生长发育。虽然激素调控植物根系生长研究较为广泛,但在激素调节的网络途径中,独脚金内酯的作用尚不明确。本文阐述了独脚金内酯的结构、生物合成过程及功能,聚焦独脚金内酯在植物激素互作调控根系生长中的作用,旨为独脚金内酯调控植物根系生长的深入研究提供参考。
1 独脚金内酯
1.1 独脚金内酯的发现
Cook等[1]最先从独脚金的非寄主植物棉花的根际分泌物分离得到独脚金醇。1998年,Yokota等[2]从小列当四叶草中提取列当醇,并确定列当醇与独脚金醇互为异构体。独脚金内酯可以诱导独脚金属和列当属种子的萌发,其次,还可以诱发丛枝菌根真菌的菌丝分枝,促进其与植物建立良好的共生关系[3]。2008年,Gomez-Roldan等[4]和Umehara等[5]同时发现独脚金内酯与生长素和细胞分裂素协同调控植物茎分枝,因而将其确定为调控植物分枝的新型植物激素。近年来,独脚金内酯的生物学功能成为植物学界研究的重要方向。
1.2 独脚金内酯的化学结构
独脚金内酯是一类萜类小分子化合物。天然SL包含1个三环内酯(ABC环)和1个由烯醇醚键连接的单环内酯(D环)[6]。不同形式的天然SL的区别主要是ABC环上取代基的种类和位置不同,但D环高度保守[7]。根据ABC环是否完整,SL分为典型和非典型两类(图1)[8-9]。典型SL具有完整的ABC环,根据C环立体构型差异可划分为两类:具有β取向的独脚金醇类和具有α取向的列当醇类[10-11],进一步修饰则可产生多种典型SL。非典型SL的B环和C环则是未闭合状态,目前,已经发现的非典型SL如图1所示。

图1 独脚金内酯的化学结构Fig. 1 Structures of strigolactone
1.3 独脚金内酯的合成
独脚金内酯的合成包括体内生物合成及体外人工合成。SL体内合成的主要部位是植物根部,合成前体是类胡萝卜素[12]。SL合成途径主要包括5个酶,分别是类胡萝卜素异构酶(DWARF27,D27)、类胡萝卜素裂解双加氧酶7(carotenoid cleavage dioxygenase 7,CCD7)、CCD8、细胞色素P450单加氧酶(cytochrome P450 monooxygenase,P450)和侧分枝氧化还原酶(lateral branching oxidoreductase,LBO),以及一个关键的中间产物——己内酯(carlactone,CL)。在水稻中,玉米黄质经D27、CCD7、CCD8作用产生的3-羟基-己内酯(3-hydroxy-carlactone,3-OH-CL)是CL形成的新途径[13]。CL在不同物种中经不同酶的作用生成己内酯酸(carlactonoic acid,CLA),进而转变为不同形式的独脚金内酯(图2)[14-18]。在独脚金内酯体内生物合成过程中,尚有部分合成步骤未彻底搞清楚,如己内酯酸转变为甲基己内酯酸,最终生成独脚金内酯,参与此过程的酶未知。

图2 独脚金内酯的生物合成通路Fig.2 Biosynthesis pathway of strigolactone
目前,已经分离的天然独脚金内酯有36种,但由于微生物降解及C环和D环之间的烯醇醚键易断裂,造成其在土壤中的稳定性较差,很难获得[19]。因此,体外人工合成是获得独脚金内酯类似物的重要方式。Bromhead等[20]以茚为底物,经臭氧化、witting反应及加成反应合成了独脚金内酯类似物GR24。
1.4 独脚金内酯的生物学功能
独脚金内酯可以促进独脚金和列当等根寄生杂草种子的萌发[1]。进一步研究表明,SL与其他激素存在协同作用[4-5],是促进高等植物根系分枝及与真菌产生共生关系[3]的关键激素。独脚金内酯能够调控植物的整体形态结构[4-5],包括参与拟南芥、水稻、豌豆、矮牵牛等植物根系的发育过程;促进叶片衰老,调节植物次生生长、光形态建成;参与生物与非生物胁迫响应等(表1)。SL主要在根中合成,茎、芽等组织中合成量极低,根系合成的SL通过短距离转运到土壤,促进菌根形成,增强根系养分吸收能力。因此,研究独脚金内酯有助于调控根部构型、改善营养吸收状况、促进植物生长。

表1 独脚金内酯的功能及其生理或分子反应Table 1 Functions and physiological/molecular responses of strigolactone
2 独脚金内酯与生长素互作调控植物根系
独脚金内酯与生长素互作,调节根系对生长素的敏感性以及根组织中的生长素含量,进而影响根系发育[40,54-55]。将受IAA抑制主根伸长的番茄幼苗在GR24中培养时,其主根长度显著增加,表明GR24可以调节根系对生长素的敏感性,削弱IAA对主根伸长的抑制效应[41]。与野生型相比,拟南芥独脚金内酯合成突变体max1和max4以及信号转导突变体max2的主根长度缩短。独脚金内酯处理可以增加野生型、max1和max4的分生区皮层细胞数目而促进主根伸长,max2的主根长度无显著差异。进一步研究发现,野生型根尖原形成层中PIN1、PIN3和PIN7融合的绿色荧光蛋白密度分别下调了38%、50%和73%[27]。因此,独脚金内酯抑制生长素转运,增加分生区和过渡区大小,促进主根伸长。
侧根生长对独脚金内酯的响应与主根相反。经GR24处理,降低侧根原基中生长素的含量,抑制野生型和max4的侧根生长。野生型中,PIN1的表达显著下调。Ruyter-Spira等[27]将侧根原基的发育划分为7个时期(I-VII)。max4中,处于V期的侧根原基中生长素含量显著降低[27,32-33],表明独脚金内酯通过抑制生长素外流,调控根组织中生长素含量,抑制侧根原基的发生,从而抑制侧根的发育。
GR24降低了番茄幼苗根毛的长度和数量。施用IAA或萘乙酸后,根毛生长不能恢复至正常水平。施用生长素外流抑制剂萘氨甲酰苯甲酸(naphthylphthalamic acid,NPA),可以恢复幼苗根毛的长度和数量,表明独脚金内酯可以促进生长素外流[41]。但GR24促进拟南芥根毛伸长[31]。在低磷条件下,IAA处理增加了max4-1、max2-1的根毛密度[40]。经GR24处理后,生长素受体突变体tir1-1根毛的长度显著低于野生型,表明当缺乏生长素时,根毛对独脚金内酯的敏感性降低,独脚金内酯通过诱导生长素受体TIR1表达正调控根毛的伸长过程[39]。
综上所述,独脚金内酯通过调节主根和侧根生长素的含量、根毛中生长素转运和TIR1表达来调控其发育过程(图3)。
3 独脚金内酯与细胞分裂素互作调控植物根系
独脚金内酯是侧根形成的负调控因子[27,31]。与野生型相比,拟南芥SL合成突变体max3和max4及信号转导突变体max2均表现侧根密度增加[31]。不同的拟南芥细胞分裂素受体(Arabidopsishistidine kinase,AHK)突变体对GR24响应不同。与野生型 相 比,突 变 体ahk2、ahk4和ahk2;ahk4的 侧根密度均显著降低;突变体ahk3、ahk2;ahk3和ahk3;ahk4差异不显著[34]。表明细胞分裂素受体中,AHK3的缺失削弱了GR24对侧根密度的负调控作用。
ARR1和ARR12是AHK3的下游正调控因子,二者协同参与AHK3调节的根系伸长过程[56]。经GR24处理后,野生型和单突变体arr1、arr12侧根密度均显著降低,双突变体arr1;arr12侧根密度无显著差异[34]。AHK3-ARR1/ARR12-SHY2共同调节GR24抑制生长素转运,进而影响侧根发育[34,57]。
与野生型相比,单突变体max2的主根较短,双突变体ahk2;ahk3的主根较长。三突变体ahk2;ahk3;max2的主根比双突变体长,表明细胞分裂素和独脚金内酯在调节主根发育过程中存在相互作用[58]。
总之,独脚金内酯与细胞分裂素互作调控主根发育过程,与生长素互作抑制的侧根生长过程依赖于细胞分裂素受体AHK3(图3)。
4 独脚金内酯与乙烯互作调控植物根系
根毛伸长受SL和乙烯正调控[31,59-60]。Kapulnik等[31]发现,经GR24处理后,野生型、乙烯信号突变体ein2-1根毛的长度均显著增加,但ein2-1根毛的长度比野生型短,max2-1、etr1-1根毛的长度无显著变化,表明乙烯和独脚金内酯共同正调控根毛的伸长。施用乙烯合成抑制剂氨氧乙基乙烯基甘氨酸(aminoethoxyvinylglycine,AVG),抑制根毛伸长,同时施用GR24和AVG,根毛长度未发生显著变化[31]。经GR24处理后,乙烯合成过程中的关键基因acs的表达量上调,乙烯产生增加[39]。因此,乙烯和独脚金内酯协同促进根毛伸长(图3)。
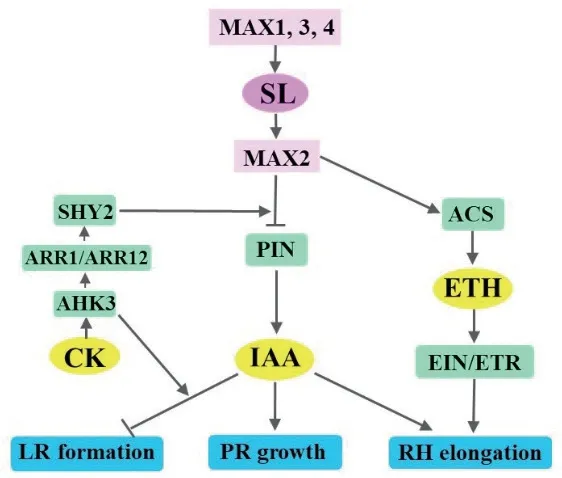
图3 独脚金内酯、生长素、细胞分裂素和乙烯对根系生长的协同调节Fig.3 Synergetic regulation of the root growth by strigolactone,auxin,cytokinin and ethylene
5 展望
独脚金内酯是由类胡萝卜素衍生的一类萜类小分子化合物,调节植物生长发育的多个过程。本文聚焦于独脚金内酯与生长素、细胞分裂素和乙烯互作调控植物根系生长,总结了独脚金内酯在激素调节根系的网络途径中的作用。关于对独脚金内酯的研究,未来应加强以下几个方面:
5.1 加强独脚金内酯生物合成过程的研究
从类胡萝卜素到独脚金内酯过程复杂,并非通过几个酶催化就能完成此过程,完整的合成过程需进一步研究,关键步骤不能突破,加上天然的独脚金内酯化合物很难获得,很大程度阻止了独脚金内酯的应用。因此,完整的独脚金内酯生物合成途径还需深入探索,为未来其在实际生产中广泛应用提供科学依据。
5.2 完善调节植物生长发育的独脚金内酯的生物活性形式
SL调节植物生长发育(包括种子萌发、枝条分枝、叶片衰老、根系发育等),但作用于不同过程的SL的生物活性形式仍然不确定[57]。通过进一步对SL前体进行生化和遗传分析,鉴定出SL生物合成过程中所包含的新的酶,将是全面了解SL作用机制的关键。
5.3 完善激素信号网络途径及其调控植物根系的机制
SL参与调控植物根系的生长发育,通过外源施用SL来调控作物根部构型和营养吸收状况,提高作物对水分及养分的吸收效率,对促进植物生长意义重大。研究发现,SL、IAA、ETH和CK等激素间很可能存在一个动态平衡,它们相互协同来调控植物的根系构型,此过程可能涉及一个极其复杂的网络系统,对该系统的深入解析可能是未来研究SL的一个重要方向[61]。
此外,SL与ABA的研究多集中于两者协同调节干旱或渗透胁迫下植物叶片气孔的闭合,但两者在调控根系发育过程中是否存在互作有待于解决。虽然已经假设GA处理对水稻根系中SL的生物合成、基因表达和分泌物有害[62],但这只发生在GA处理24 h后,而不是对GA处理的直接反应[62-63]。因此,在植物根系中SL与GA是否存在直接相互作用也有待于研究。
总之,独脚金内酯作为一种新型植物激素,其为植物生物学领域开拓了一片新的研究领域,但相关的研究启动较晚而又意义重大。因此,其中存在的科学问题有待于不同领域的科学家深入探讨。
