S100A16与成骨分化的生物信息学分析及功能验证
2022-09-13沈亚非王朝旭谢亚争
辛 婧 沈亚非 王朝旭 谢亚争
S100A16是钙调蛋白S100家族的新成员,S100蛋白与细胞增殖、分化、凋亡与肿瘤的发生、发展密切相关。研究表明,S100A16在正常食道、脂肪、肺、甲状腺、胰腺、结肠等组织中高表达,且在不同的组织中参与了不同的细胞发育、细胞生长和细胞凋亡作用,有可能存在比S100家族其他成员更复杂的调控机制[1,2]。S100A16是笔者课题组在前期对肥胖相关基因研究中通过基因芯片筛查到的差异表达基因,并证实S100A16促进前体脂肪细胞分化、脂质合成从而促使肥胖发生[3,4]。脂肪细胞和成骨细胞均来源于间充质干细胞,两者存在此消彼长的关系,即S100A16促进脂肪细胞分化的同时可能抑制成骨细胞生成[5]。目前,S100A16与成骨分化的相关性研究鲜见报道。本研究利用GEO数据库的高通量测序数据对S100A16与成骨分化进行生物信息学相关性分析,并通过构建S100A16慢病毒载体转染骨髓间充质干细胞诱导成骨分化,观察S100A16对成骨分化的影响,初步探讨S100A16基因在成骨分化中的作用,为进一步研究骨质疏松发生的分子机制及寻找新的治疗靶点提供理论依据。
材料与方法
1.数据库及分析数据:采用来自美国国立生物技术信息中心(NCBI)的公共基因芯片数据库(Gene Expression Omnibus,GEO)和可视化平台GEO2R对S100A16与成骨分化进行生物信息学相关性分析。
2.材料:DMEM培养基、胎牛血清购自美国Hyclone公司;地塞米松、茜素红、FBS缓冲液、β-甘油磷酸钠购自美国Sigma公司;RNA提取试剂盒、反转录试剂盒、real-time PCR反应试剂盒购自日本TaKaRa公司;PLKO.1-sP6-GFP空病毒载体由南京医科大学分子遗传研究室李建民教授馈赠;引物由上海英骏生物技术有限公司合成;Pierce BCA蛋白浓度测定试剂盒购自美国Thermo Fisher公司;S100A16、CD14、CD34、CD45、CD44、CD90、CD105兔源一抗、PE偶联羊抗兔二抗均购自武汉三鹰生物技术有限公司;IgG-辣根过氧化物酶羊抗兔二抗、β-actin抗体购自美国CST公司。
3.生物信息学数据处理:根据GEO数据集中不同收录信息,通过转录组测序检测S100A16在正常C57BL6小鼠各主要组织器官中的表达情况(GEO测序数据登录号:GDS3142);检测4组正常MC3TC-E1成骨前体细胞(control)、3组曲古抑菌素 A诱导成骨分化的成骨前体细胞(TSA treated)中S100A16的表达情况(GEO测序数据登录号:GSE92470)。在GEO2R分析页面输入GEO测序数据登录号,确定样本分组后,使用默认参数进行S100A16的表达量分析,导出生成的柱状图。
4.大鼠骨髓间充质干细胞分离与培养:所有动物实验和操作顺序经漯河市中心医院动物伦理委员会批准通过(审批号:2020018),严格遵守动物实验伦理条例,并按照国家卫生研究院“实验动物保护和使用指南”的规定执行。颈椎脱臼处死大鼠后用75%乙醇溶液浸泡15min,无菌操作分离股骨与胫骨,用DMEM培养液吸取大鼠骨髓轻吹打使溶液混匀,选用22号注射针头滤制溶液制备成单细胞悬液,接种在细胞培养皿,置于37℃、5% CO2孵箱中培养,每2~3天换液。传至第3代流式细胞仪鉴定为骨髓间充质干细胞(BMSCs)。
5.大鼠骨髓间充质干细胞表面标志物的鉴定:选取第3代生长状态良好的大鼠骨髓间充质干细胞,胰酶消化3min,收集细胞悬液于1000r/min室温离心5min,弃上清后用无菌PBS重悬细胞并过200目筛网制成单细胞悬液。各管样本分别加入CD44、CD90、CD105、CD14、CD34、CD45的单克隆抗体,同时设立同型阴性对照,4℃避光孵育,用流式细胞仪进行检测。
6.慢病毒载体转染骨髓间充质干细胞并诱导成骨分化:笔者课题组早期构建了过表达S100A16慢病毒载体(PLJM1-S100A16-GFP),转染大鼠BMSCs并筛选稳定感染细胞株,建立S100A16过表达组、空载对照组和未转染载体的正常BMSCs组。分别加入成骨细胞诱导液(含10%PBS的DMEM培养基+10mmol/L β甘油磷酸钠+0.05mmol/L维生素C+100mmol/L地塞米松),每3天换液1次,持续诱导培养21天,检测相关指标。
7.Western blot法检测:分别抽提第3代正常BMSCs组、转入慢病毒载体PLJM1-S100A16-GFP的S100A16过表达组及转入空载质粒PLJM1的空载对照组的总蛋白质,测定蛋白质浓度,制备12%的分离胶和5%的浓缩胶,100V稳压电泳,半干转膜。一抗为浓度1∶500的S100A16多克隆抗体,二抗为1∶2000的羊抗兔IgG一辣根过氧化物酶,用免疫印迹化学发光法检测。以 GAPDH 作为内参,应用 Image J软件分析各组蛋白条带灰度比值的变化。
8.RT-PCR检测成骨细胞相关因子mRNA表达水平:取S100A16过表达组、空载对照组、正常BMSCs组的第3代BMSCs细胞,诱导成骨分化,培养14天提取总RNA,反转录得cDNA,利用实时定量PCR检测成骨分化相关因子碱性磷酸酶(alkaline phosphatase,ALP)、骨钙素(osteocalcin,OCN)、Runt 相关转录核心因子 2(Runx2)的表达,GAPDH 设为内参,结果以2-ΔΔCt表示,绘制柱状图。相关引物序列详见表1。

表1 实时定量PCR引物序列
9.茜素红染色检测成骨分化能力:在BMSCs向成骨分化过程中,细胞表面沉积钙盐形成钙结节,茜素红与钙发生显色反应,通过茜素红染色鉴定钙结节形成情况。BMSCs 成骨诱导分化21天后,弃培养液,PBS洗涤 2~3 次,甲醛缓冲液固定15min,蒸馏水洗涤3次,加入0.1%茜素红-Tris-HCL染液(pH值为8.3),37℃下染色30min,蒸馏水冲洗,显微镜下观察茜素红染色阳性细胞。

结 果
1.S100A16在各组织中的表达情况生物信息学分析:转录组测序检测S100A16在正常C57BL6小鼠各主要组织器官中的表达情况,分析发现S100A16基因在骨髓中的表达显著低于其他组织(图1),提示其表达与成骨呈负相关。
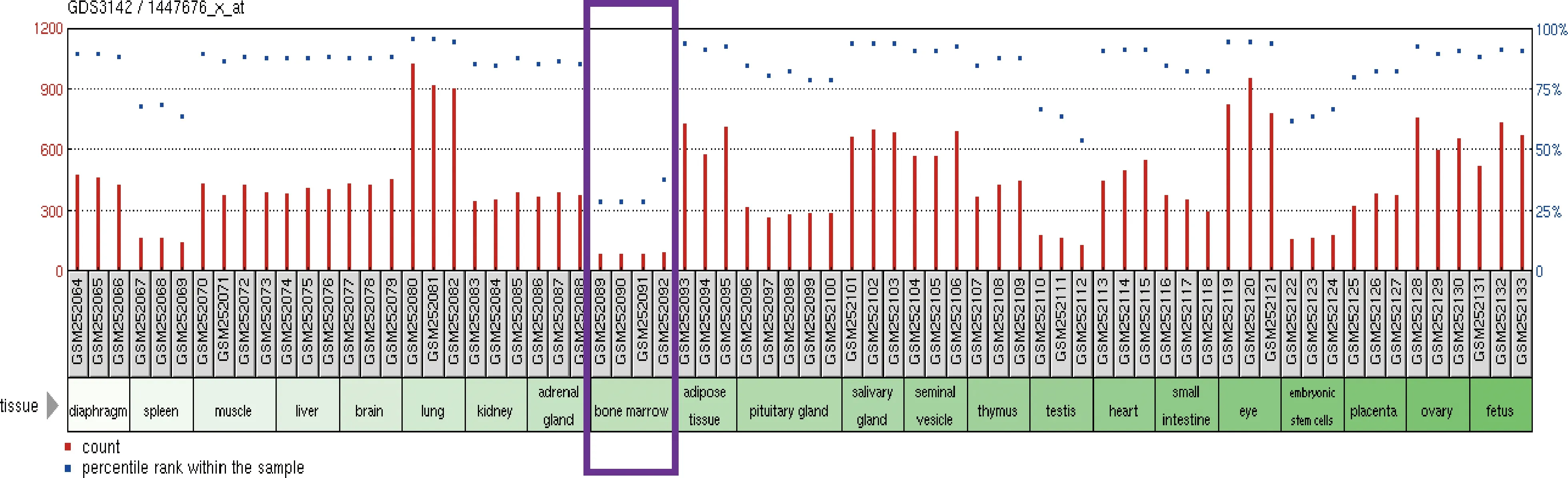
图1 S100A16在正常C57BL6小鼠各主要组织器官中的表达情况
2.S100A16在成骨细胞分化中的表达情况分析:已有文章表明曲古抑菌素 A可促进骨髓间充质干细胞的成骨分化[6,7]。转录组测序对比正常MC3TC-E1成骨前体细胞(control)和曲古抑菌素 A诱导成骨分化的成骨前体细胞(TSA treated)中S100A16的表达情况,分析发现S100A16基因在经TSA诱导成骨分化的细胞中表达量显著降低(图2),提示S100A16表达量与成骨分化呈负相关。

图2 S100A16在正常成骨前体细胞组和曲古抑菌素A诱导分化组中的表达情况
3.骨髓间充质干细胞流式鉴定:取生长状态良好的第3代大鼠骨髓间充质干细胞,经流式细胞仪鉴定细胞表面标志物,结果显示CD44、CD90和CD105的阳性率分别为96.6%、96.9%和97.7%。而CD14、CD34和CD45呈阴性表达(图3),符合骨髓间充质干细胞特征。

图3 流式鉴定骨髓间充质干细胞表面标志物骨髓间充质干细胞表面阳性标志物(CD44、CD90和CD105)表达均在96%以上,阴性标志物(CD44、CD34和CD45)表达均在2%以下,均符合要求
4.S100A16在各组细胞的表达情况:收集各组细胞蛋白,Western blot法检测结果显示,S100A16过表达组(PLJM1-S100A16-GFP-BMSCs)与空载对照组(PLJM1-BMSCs)及正常BMSCs组比较,S100A16蛋白表达显著升高(P<0.05,图4),表明S100A16稳定高表达细胞株构建成功。
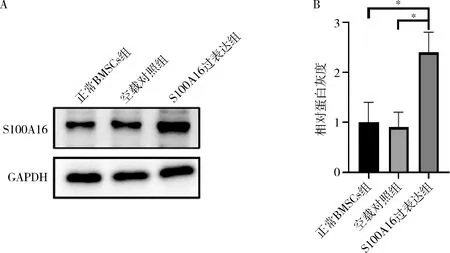
图4 S100A16蛋白表达情况A.S100A16蛋白表达的免疫印迹条带;B.S100A16相对于GAPDH蛋白表达水平的灰度值;*P<0.05
5.各组细胞成骨分化相关因子mRNA表达情况:实时定量PCR结果显示,S100A16过表达组(PLJM1-S100A16-GFP-BMSCs)细胞成骨分化相关因子ALP、OCN、Runx2的mRNA表达水平较正常BMSCs组及空载对照组表达明显下降(图5),说明S100A16抑制成骨分化。
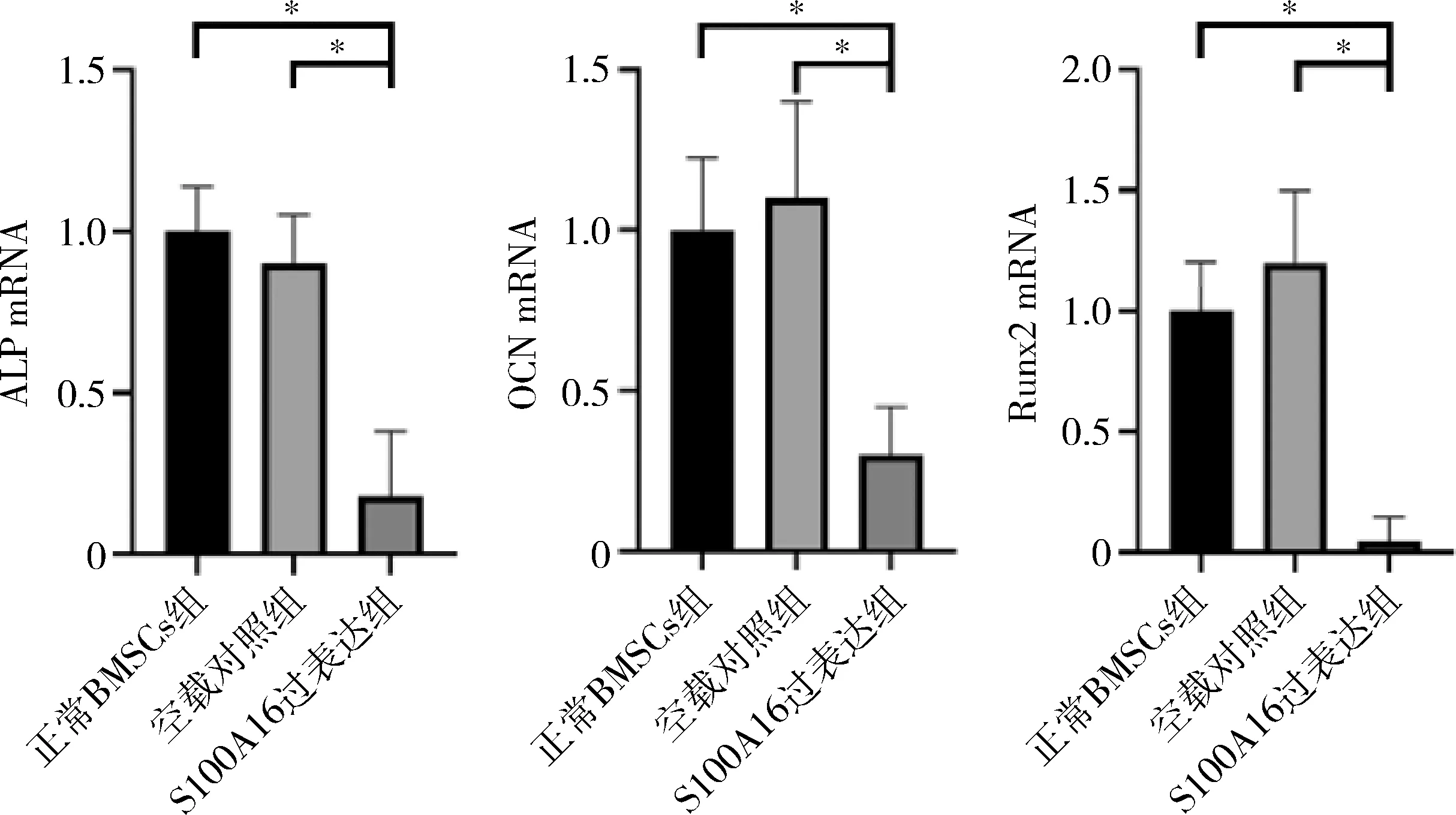
图5 细胞成骨分化相关因子mRNA的表达情况ALP.碱性磷酸酶;OCN.骨钙素;Runx2.Runt 相关转录核心因子2;*P<0.05
6.茜素红染色观察各组细胞骨矿化结节形成情况:各组BMSCs成骨诱导分化21天,茜素红染色显示S100A16过表达组(PLJM1-S100A16-GFP-BMSCs)矿化结节形成较正常BMSCs组和空载对照组明显减少(图6)。

图6 各组细胞成骨诱导21天后茜素红染色观察矿化结节形成情况(×200)A.正常BMSCs组;B.空载对照组;C.S100A16过表达组
讨 论
骨质疏松症(osteoporosis,OP)是一种与年龄、性别、环境、遗传等相关的退行性疾病,主要的危害就是发生骨质疏松性骨折[8,9]。近年来,国家重点宣传“全民健康,三减三健”的主题,其中包括健康骨骼,骨质疏松是全民需要重视的问题。骨质疏松导致的髋部骨折被称为是“人生最后一次骨折”,我国骨质疏松症患者髋部骨折的平均年龄是67.2岁,每年医疗费用估计需要150亿元人民币[10,11]。因此,骨质疏松给家庭和社会都带来了极大的负担和压力,这是现代人口老龄化社会的一个亟需解决的健康问题,需要有效的防治策略。骨质疏松的主要致病因素是骨吸收增强和骨形成能力减弱。成骨细胞分化在调控骨形成中发挥重要作用,也是目前研究的热点。本研究通过探讨S100A16与成骨分化的相关性为骨质疏松症的早期诊断分子标志物的筛选及分子靶向治疗提供新的思路。
骨髓间充质干细胞在一定条件下可分化为脂肪细胞、成骨细胞和软骨细胞,在BMSCs分化过程中,成脂分化和成骨分化呈负相关,成脂-成骨分化状态依赖于多种转录因子和信号通路共同调控[12]。S100A16是笔者课题组在前期对肥胖相关基因研究中通过基因芯片筛查到的差异表达基因。笔者课题组前期研究发现,S100A16能够促进前体脂肪细胞分化、脂质合成集聚从而导致肥胖发生[13,14]。另有研究显示,肥胖患者脂肪分化增加、成骨分化减少,成骨细胞被脂肪细胞替代、骨髓被脂肪填充致骨矿化密度下降可引发骨质疏松倾向[15,16]。而骨质疏松症患者骨量减少的同时伴有骨髓脂肪组织增加致骨矿化密度下降[17]。这些均提示S100A16在促进脂肪分化同时抑制成骨分化,笔者推测S100A16与成骨分化呈负相关。目前S100A16作为促进脂肪细胞分化和形成的关键因子,是否影响且如何影响成骨分化无相关报道。
为此,本研究首先进行了S100A16的表达与成骨分化的生物信息学相关性分析,结果显示,S100A16基因在骨髓中的表达显著低于其他组织,提示其与成骨呈负相关;S100A16基因在经曲古抑菌素A诱导成骨分化细胞中表达量较正常对照组显著降低,提示S100A16表达与成骨细胞分化呈负相关。为进一步验证S100A16与成骨分化的相关性,首先分离培养大鼠骨髓间充质干细胞,经流式细胞仪鉴定细胞表面标志物,结果显示,CD44、CD90和CD105的阳性率均大于90%,CD14、CD34和CD45呈阴性表达,符合骨髓间充质干细胞特征。笔者构建S100A16高表达慢病毒载体转染骨髓间充质干细胞并诱导成骨分化,研究S100A16对成骨分化的影响。从蛋白水平验证S100A16稳定转染细胞株构建成功。与正常BMSCs组和空载对照组比较,S100A16高表达下调骨髓间充质干细胞诱导成骨分化过程中成骨分化相关因子碱性磷酸酶、Runt相关转录核心因子 2的表达。矿化结节是成骨细胞分化成熟的形态学表现,通过茜素红染色笔者观察到S100A16高表达组矿化结节明显少于空载对照组,说明S100A16抑制成骨分化。因此,笔者推测S100A16通过调控成骨细胞的分化而促使骨质疏松发生、发展。目前缺乏关于S100A16参与成骨细胞分化及骨质疏松发生、发展的相关性研究,因此两者关系及其作用机制还需进一步基础研究及后续随机、多中心的大样本量分析加以论证和核实。
综上所述,生物信息学分析及实验研究结果显示,S100A16表达与成骨分化呈负相关,它作为全新的成骨分化呈负性调控因子可能是抗骨质疏松的潜在治疗靶点,值得进一步探索。本研究的意义在于丰富成骨分化和骨质疏松理论基础研究的同时,也将为骨质疏松症的筛查及防治提供新的研究方向。
