多酚-膳食纤维相互作用及其影响多酚生物利用率研究进展
2022-09-09刘甜甜吴晓娟
刘甜甜,吴晓娟,吴 伟
(中南林业科技大学食品科学与工程学院;稻谷及副产物深加工国家工程实验室,长沙 410004)
多酚是一种广泛存在于植物源食品中的生物活性物质,具有良好的抗氧化、抗肿瘤、抗病毒、缓解抑郁、调节胆固醇水平和延缓心脑血管疾病等生理功能[1-3]。多酚只有在胃肠道消化过程中从食品基质中释放出来,通过肠道消化吸收进入人体代谢,才能充分发挥其健康功效,因而生物利用率对于评估多酚的生理功能具有重要意义[4]。天然多酚绝大多数以结合态存在,与膳食纤维、蛋白质、脂质以及其他小分子物质形成复合结构。在储藏加工和机体消化过程中,多酚与其他组分会发生复杂的反应,多酚的存在形态可能会发生改变,影响其生物利用率[5]。膳食纤维是目前已知的与多酚结合最为广泛,且紧密的膳食组分之一。多酚与膳食纤维可通过酯键、醚键或碳-碳键等形成共价化合物,还可通过氢键、疏水作用力、静电作用力等产生非共价相互作用。膳食纤维的结合可提高多酚的化学稳定性,有利于保持其生物活性,但另一方面,膳食纤维的束缚作用会减少多酚在胃肠道中释放,降低其生物可及性。近年来有许多关于提高多酚生物利用率的策略,例如:通过化学修饰提高多酚的生物可及性,破坏阻碍多酚释放的大分子结构,或将多酚装载于微米或纳米载体等,但从膳食纤维结合作用的角度分析多酚生物利用率的研究才刚刚起步[6,7]。因此,本文将重点讨论多酚与膳食纤维相互作用的机理及其对多酚生物利用率的影响,为多酚-膳食纤维复合物在食品领域的应用提供参考。
1 膳食纤维与多酚的相互作用
多酚是一类含有一个芳香环以及一个或多个羟基的植物次生代谢物,根据羟基的数目和位置,可分为酚酸、黄酮、二苯乙烯和木质素等[8]。目前,研究最为广泛的酚类物质主要为酚酸和黄酮[8]。酚酸是一类含有酚醛环和至少1个羧基的物质,主要有水杨酸、没食子酸、苹果酸、阿魏酸和咖啡酸等;黄酮包括2个苯环和1个杂环,根据杂环的氧化程度,可以进一步表征为黄酮、黄酮醇、黄烷酮、黄烷醇、异黄酮和花色苷等[9]。
膳食纤维是一种多糖,可分为可溶性膳食纤维和不溶性膳食纤维。可溶性膳食纤维包括果胶、β-葡聚糖、树胶、低聚糖、菊粉等,不溶性膳食纤维主要包括纤维素、半纤维素、甲壳素和抗性淀粉[10]。可溶性膳食纤维具有良好的抗氧化性质,可作为天然抗氧化剂调节机体氧化应激水平,改善氧化损伤[11];不溶性膳食纤维有利于润肠通便[12]、预防肥胖[13]和减少有害物质蓄积对人体造成损伤[14]等。通常不溶性膳食纤维中酚类物质含量要明显高于可溶性膳食纤维含量,特别是在谷物中,酚类物质大多分布在由不溶性膳食纤维构成的麸皮中,并且多酚与纤维素基质结合的比例要显著高于其他基质[15,16]。
膳食纤维和多酚之间的相互作用可以分为共价结合和非共价结合2种形式,其表征方法主要有傅里叶变换红外分光光度法、紫外分光光度法、核磁共振波谱分析、差示扫描量热法分析、等温滴定量热法和X射线光电子能谱分析等[9]。共价结合过程是不可逆的,作用很强,而非共价结合过程是可逆的,作用较弱。
1.1 共价结合
多酚可通过酯键、醚键或碳-碳键等与膳食纤维共价键合。如图1所示,膳食纤维可通过羟基与酚酸羧基形成酯键,或与芳香环羟基形成醚键,还可通过碳原子与酚类化合物的碳原子相连。根据不同的共价结合方式,陈彩薇等[17]发现米糠中4种不同存在形态酚类物质的含量从高到低依次为:不溶性酯型结合酚、不溶性苷型结合酚、可溶性酯型和可溶性苷型结合酚。其中,不溶性酯型结合酚占脱脂米糠中总酚质量的32.48%,明显高于其他存在形态结合多酚含量,说明脱脂米糠中的多酚多以酯键形式与不溶性大分子物质(如膳食纤维)结合。杜小燕等[18]将小麦麦麸中的结合酚类物质分为酯苷结合型、糖苷结合型、碱解束缚型和酸解束缚型结合酚,发现碱解束缚型酚类物质的含量最高,占总酚的63.00%,与脱脂米糠中不同存在形态结合多酚含量分布的结论基本一致。此外,贾亮[19]发现杏仁种皮中酸水解可溶性结合酚含量显著高于碱水解可溶性结合酚,而碱水解不溶性结合酚含量和种类均高于酸水解不溶性结合酚,推测杏仁种皮中可溶性结合酚主要以糖苷键与膳食纤维结合,而不可溶性结合酚主要以酯键与膳食纤维结合。Wang等[20]发现枣子酚类物质主要以糖苷结合型和不溶性结合酚形式存在,其中果核和果皮中的酚类物质主要为不溶性结合酚,而果肉中的多酚主要为糖苷结合型酚。除天然存在的多酚-膳食纤维共价结合复合物,多酚与膳食纤维间的共价相互作用还可通过氧化反应介导、多酚氧化酶催化或在热酸条件下生成。Fernandes等[21]以抗坏血酸/过氧化氢氧化还原对作为自由基引发剂,发现海带多糖上的羟基与没食子酸的羧基形成了酯键结合。Suyeon等[22]以漆酶为催化剂促使儿茶素的氧化产物邻醌和壳聚糖上的氨基成功结合。Olive等[23]在酸性条件下观察到儿茶素与葡聚糖酐产生了共价缩合。
1.2 非共价结合
如图2所示,多酚与膳食纤维非共价相互作用主要包括氢键、疏水相互作用和静电或离子相互作用。多酚和膳食纤维中含有许多羟基基团,这为氢键的形成提供了可能。氢键通常在多酚的羟基和膳食纤维的糖基键或羟基之间形成。如果先形成氢键,多酚和膳食纤维分子之间的距离就会缩短,从而产生范德华力。当一些不溶性的多酚和膳食纤维暴露在水中时,为了减少与水的接触面积,多酚和膳食纤维自发的聚集在一起,就会产生疏水相互作用[4]。表没食子儿茶素没食子酸酯与燕麦β-葡聚糖的相互作用以氢键为主[24];槲皮素与β-葡聚糖的相互作用以疏水作用力为主[25];白藜芦醇与羧甲基化(1,3/1,6)-β-D-葡聚糖间以弱静电相互作用为主[26]。天然多酚与膳食纤维间通常同时存在多种不同的非共价相互作用,如:茶多酚与燕麦β-葡聚糖间为氢键和范德华力[27];苹果细胞壁与原花青素存在疏水相互作用和氢键作用[28];Koh等[29]发现花青素的风味阳离子与蓝莓果胶的自由羧基之间同时存在氢键、疏水相互作用和静电相互作用。虽然与共价结合相比,非共价结合作用强度较弱,但是非共价结合也可以提高多酚的稳定性,避免光、热导致的多酚降解[30, 31]。
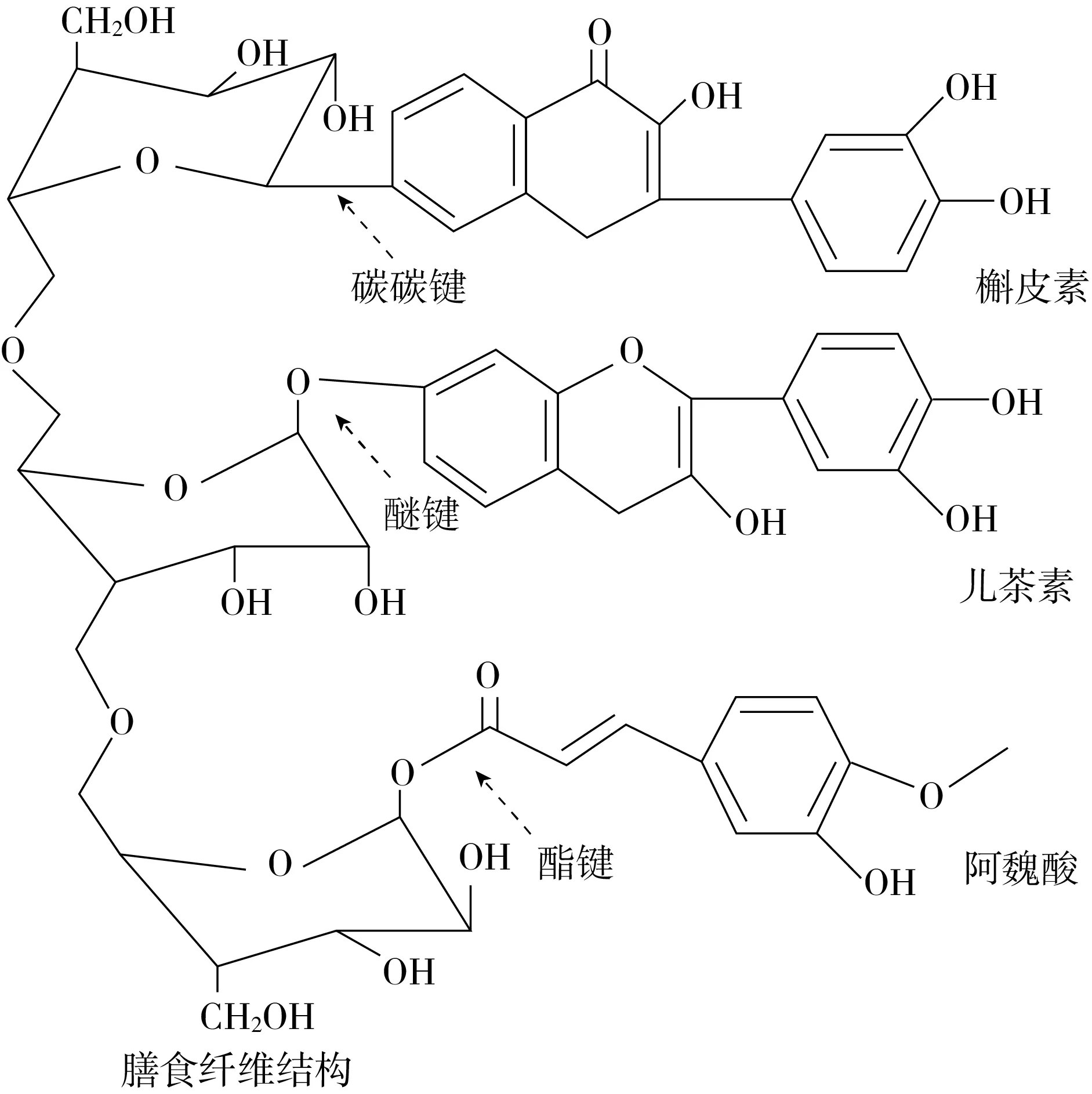
图1 多酚与膳食纤维的共价结合方式
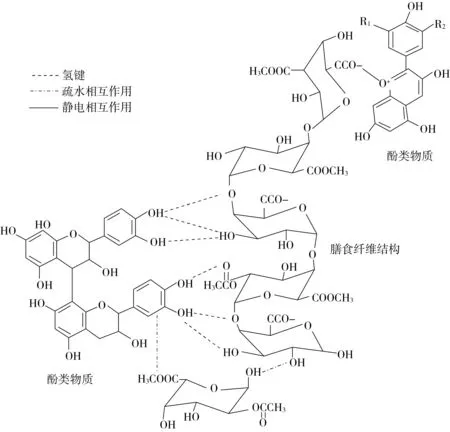
图2 多酚与膳食纤维的非共价结合方式
2 影响多酚-膳食纤维相互作用的因素
不同来源膳食纤维与多酚的结合方式以及作用力大小主要由膳食纤维和多酚的结构决定,并受温度、pH值、离子强度和氧化应激等环境因素影响[8],具体如表1所示。
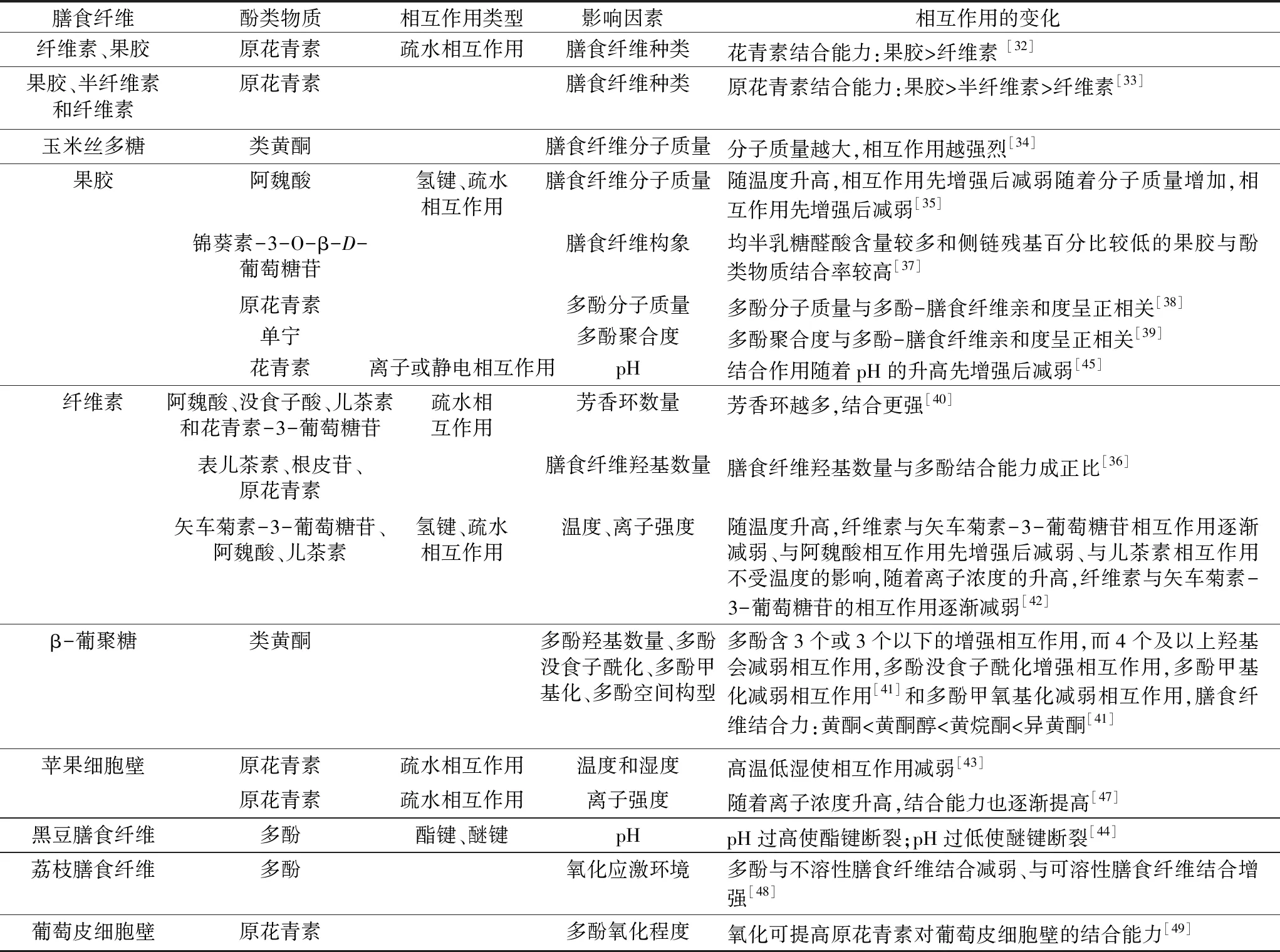
表1 不同因素对多酚-膳食纤维相互作用的影响
2.1 膳食纤维结构对多酚-膳食纤维相互作用的影响
膳食纤维结构复杂,影响多酚-膳食纤维结合作用的因素主要是膳食纤维的种类、分子质量以及分子结构和构象等。有研究表明,与其他膳食纤维相比,果胶对多酚具有相对较高的亲和力,因为在果胶分子形成凝胶的过程中,可形成疏水的袋状结构,这有利于产生疏水相互作用[32]。Ruiz等[33]报道,葡萄皮中的膳食纤维对原花青素的吸附能力从大到小依次为:果胶、半纤维素和纤维素。膳食纤维的分子质量影响多酚与膳食纤维结合的机制复杂。一些认为,分子质量高的膳食纤维结构更复杂,分支更多,存在更多的吸附位点,因此更容易与酚类物质相互作用[34]。Zhang等[35]发现果胶对阿魏酸的吸附量随着果胶分子质量的增加先增加后减少,可能是分子质量适度增加可以使膳食纤维分子链变长,膳食纤维的吸附能力增加,但分子质量过大会导致膳食纤维聚集度增大、分子间空隙变小,导致吸附能力减弱。此外,Liu等[36]发现,纤维素结构中可用的羟基数量与多酚结合能力成正比。Fernandes等[37]发现,均半乳糖醛酸含量较多和侧链残基百分比较低的葡萄皮果胶多糖与锦葵素-3-O-β-D-葡萄糖苷有较高的结合率,这可能与膳食纤维更高的线性度和灵活性有关。
2.2 多酚结构对多酚-膳食纤维相互作用的影响
多酚的聚合度、酚环数量、空间构型和某些特定基团的存在等都会影响多酚-膳食纤维的相互作用。分子质量和聚合度与原花青素对膳食纤维的亲和度呈正相关性,因为分子质量较大的多酚上有更多可以成键的基团[38]。Mamet等[39]发现,高聚合单宁与低聚合单宁相比,与果胶的结合能力更强,因为高聚合单宁中没食子酸酯部分的存在可以增强与高度甲基化果胶的亲和力。分子结构中芳香环较多的酚类化合物与纤维素的结合更强,因为越多芳香环的多酚与膳食纤维形成疏水相互作用的可能性越大[40]。在类黄酮异构体中,与燕麦β-葡聚糖的吸附量按黄酮、黄酮醇、黄烷酮和异黄酮的顺序依次增加;邻香豆酸对燕麦β-葡聚糖的吸附能力高于对香豆酸和间香豆酸;3个或3个以下的羟基可以增强黄酮类多酚对β-葡聚糖的吸附能力,而4个及以上羟基会减弱黄酮类多酚对β-葡聚糖的吸附能力;没食子酰化会提高儿茶素对β-葡聚糖的的吸附能力;而甲基化或甲氧基化会降低酚酸对β-葡聚糖的吸附能力[41]。
2.3 环境因素对多酚-膳食纤维相互作用的影响
在食品储藏和加工的过程中,多酚与膳食纤维之间的结合强度会受到各种环境因素影响,主要的环境因素有:温度、pH值、离子强度和氧化应激环境。
温度对于多酚与膳食纤维之间的氢键和疏水相互作用具有重要意义。形成氢键是一个释放能量的放热过程,而形成疏水相互作用是一个需要能量的吸热过程,多酚与膳食纤维间通常同时存在氢键与疏水键,不同的多酚-膳食纤维结构中氢键与疏水键比例不同,因此其亲和度随温度变化的趋势并不一致[4]。例如,随着温度的上升,纤维素对矢车菊素-3-葡萄糖苷的吸附能力逐渐减弱,对阿魏酸吸附能力先增强后减弱,而对儿茶素吸附能力不受温度的影响[42]。在阿魏酸与果胶的相互作用中,温度的升高增加了吸附量,可能意味着疏水键的生成;温度的进一步升高降低了吸附量,可能与氢键的变化相关[35]。在食品加工过程中,温度常与湿度同时影响膳食纤维对多酚的吸附能力,干燥通过减小膳食纤维的表面积、降低孔隙率减少了膳食纤维与多酚的结合[43]。
pH值可以显著影响多酚与膳食纤维之间的共价和非共价相互作用,并使多酚的结构发生变化。pH值过高会使多酚-膳食纤维间的酯苷键断裂,而pH值过低则会使多酚-膳食纤维间的醚苷键断裂,因此人们常用碱水解和酸水解的方法来提取膳食纤维中的结合多酚[44]。Lin等[45]发现花青素与果胶的结合作用随着pH值的升高先增强后减弱,因为pH值适度增加会使多酚-膳食纤维间离子或静电相互作用增强,而pH值过高会导致多酚发生降解。
溶液的离子强度对于多酚-膳食纤维的疏水相互作用至关重要。有机分子间的疏水相互作用随着离子强度的增加而增加[46]。离子浓度上升使苹果膳食纤维与花青素的结合能力提高,可能是由于离子强度的增加导致多酚-膳食纤维间疏水相互作用增加[47]。Phan等[42]发现,纤维素对矢车菊素-3-葡萄糖苷的吸附能力随着离子浓度的升高而降低;对儿茶素的吸附能力不受离子浓度的影响,说明纤维素与矢车菊素-3-葡萄糖苷和儿茶素间可能不存在疏水相互作用。
在食品储藏加工过程中,会产生许多具有强氧化性的自由基。当环境中产生和清除自由基的能力不能达到平衡时会生成氧化应激环境。Xu等[48]发现,荔枝“Guiwei”经过一年时间的储藏后,荔枝果肉中总膳食纤维中结合酚含量基本不变,但不溶性膳食纤维中的结合酚含量降低,而可溶性膳食纤维中的结合酚含量增加。荔枝储藏使得多酚与膳食纤维的结合方式发生了变化,这种变化可能是通过氧化使多酚和膳食纤维结构改变来实现的。Bautista-等[49]研究了6种不同种类的原花青素类单宁与葡萄皮不溶性细胞壁物质(膳食纤维)的相互作用,发现氧化可提高原花青素对葡萄皮细胞壁的结合能力,表没食子儿茶素没食子酸酯相较于其他5种酚类物质表现出了较高的膳食纤维亲和力。
3 多酚-膳食纤维相互作用对多酚生物利用率的影响
3.1 多酚-膳食纤维相互作用对多酚生物可及性的影响
多酚的生物可及性是食物基质中释放的酚类物质经消化后可被胃肠道黏膜吸收的百分比。结合多酚在胃肠道中会发生一系列的酶促反应,以游离和可溶性结合的方式释放到胃和小肠的消化液中。多酚-膳食纤维相互作用会严重阻碍人体胃肠道内酯酶对不溶性多酚的水解。当食物基质中含有菊粉微粒时,橄榄苦苷经胃肠道消化后的生物可及性降低了27.5%[50]。Carrillo等[51]证明在没有细胞壁物质的情况下,花青素的生物可及性更强。米糠膳食纤维经胃肠道消化后,仅释放19.91 mg /100 g DW的酚类物质,占米糠膳食纤维中总结合酚类物质的2.68%[52]。膳食纤维的结构(如粒径大小)对结合多酚的生物可及性有重要影响。不溶性膳食纤维结合多酚的生物可及性与膳食纤维粒径的大小呈反比,减小膳食纤维颗粒的大小可以提高多酚在小肠中的生物可及性[53]。膳食纤维的性质(如溶解性)对结合多酚的生物可及性也有较大影响。Zhao等[54]发现,米糠不溶性膳食纤维(IDFDRB)结合酚的生物可及性远低于米糠可溶性膳食纤维(SDFDRB)结合酚,IDFDRB的多酚生物可及性仅为18.8%,而SDFDRB的多酚生物可及性达87.4%,IDFDRB中的酚类物质主要在胃中释放;而SDFDRB中的酚类物质主要在小肠中释放。
3.2 多酚-膳食纤维相互作用对多酚生物利用率的影响
多酚的生物利用率是多酚在食用后被吸收到血液循环系统的程度[54]。结合多酚被人体利用的过程如图3,人摄入食物后,胃和小肠消化将酚类物质从食物基质中释放出来,然后胃肠道表皮细胞将游离酚吸收进入血液循环,血浆将它们分布到达人体不同靶向器官,发挥不同的生物活性,最后经代谢排出体外。多酚在胃和小肠中生物可及性很低,大多数的结合多酚会到达消化道下部——大肠,大肠微生物如双歧杆菌和乳酸菌产生的糖酶、酯酶和其他胞外酶可以水解多酚和膳食纤维的共价键,释放多酚并将其发酵成各种分解代谢物。Merve等[55]发现,添加果胶和菊粉可以有效提高黑莓泥中某些酚类物质在体外结肠发酵后释放的含量,并产生了一些低分子质量代谢物,如酪醇、没食子酸和苯甲酸,这些简单酚类化合物可以调节免疫系统、调节肠道菌群激活短链脂肪酸排泄、促进肠道健康。酚类物质的吸收可以通过协助扩散和主动运输来实现[56,57]。多酚被上皮细胞吸收后进入血液循环。Pimpao等[58]在摄入富含酚类的浆果果泥的志愿者的血浆中发现了几种酚硫酸盐,这些代谢物在血液循环下可跨越血脑屏障内皮细胞,发挥神经保护作用。与脂肪和蛋白质相比,增加碳水化合物的摄入可以显著增加血浆中表儿茶素和儿茶素黄烷醇的含量[59]。目前有关膳食纤维结合多酚生物利用率的研究报道主要集中于结合多酚在胃肠道消化环境的释放和利用方面,而关于真实的贮藏加工条件下多酚-膳食纤维相互作用的变化,以及这些变化如何影响多酚生物利用率的研究较少。

图3 膳食纤维结合多酚在胃肠道中的吸收和代谢
4 展望
目前对于多酚-膳食纤维相互作用的研究几乎都仅限于简单的环境因素对于多酚和膳食纤维亲和性的影响。但在复杂的食品贮藏加工或者机体消化条件下,多酚和膳食纤维相互作用具体发生怎样的变化研究还不够深入。在不同的环境条件下,到底有哪些非共价作用增强、哪些非共价作用减弱,不同加工强度是否导致多酚和膳食纤维结合作用不同程度的变化,这些变化又怎么影响多酚的消化和吸收尚不清楚。因此,将食品加工和人体消化环境下多酚-膳食纤维具体的结构变化规律阐明,找到多酚-膳食纤维相互作用变化规律与多酚生物利用率之间的关联,是未来研究的重点。这对于改善多酚-膳食纤维的功能性质和提高多酚的生物利用率具有重要意义。
