CRISPR/Cas9编辑芥菜型多心皮油菜BjROP10基因的研究
2022-09-03徐珂李爽赵歌孙振宇李海源李珂欣王效华徐平沈金雄
徐珂,李爽,赵歌,孙振宇,李海源,李珂欣,王效华,徐平*,沈金雄
(1.临沂大学农林科学学院,山东 临沂,276000;2.华中农业大学作物遗传改良国家重点实验室/国家油菜工程技术研究中心,湖北 武汉,430070)
油菜是世界上重要的油料作物之一,也是主要的食用植物油来源。油菜通常为两心皮角果,每角果粒数多为15~20 粒,但在近几年的研究中发现了多心皮油菜突变体,在单株角果数和千粒重相近的前提下,多心皮油菜每角果粒数可达30~40 粒[1],是一种有利于油菜高产的优良性状[2,3]。因此开展油菜心皮数发育分子机制的研究对揭示油菜心皮发育的机理和培育多心皮高产油菜具有非常重要的理论和实践意义。
已有研究发现,CLAVATA(CLV)信号途径相关基因CLV1、CLV2和CLV3的同源基因突变均可产生油菜多心皮性状[4,5],但对其调控机制尚不明确。G蛋白是所有真核生物中多种信号通路的关键调控因子。在植物中,G蛋白与多种农艺性状相关,例如产量、器官大小、生物和非生物胁迫反应、共生和氮利用效率等,因此该类蛋白是目前植物研究的热点[6]。研究发现,玉米中的Gα 蛋白(异源三聚体G蛋白的α 亚基)基因CT2通过传导CLV 信号来调控茎顶端分生组织中干细胞的生长[7],该基因的过量表达可使玉米产生小穗密度和籽粒行数增加、穗分生组织增大以及叶片直立等优异的农艺性状变异[8]。拟南芥中的Gβ 蛋白(异源三聚体G 蛋白的β亚基)基因AGB1也可通过与另一个类似CLV1的受体RPK2的互作调节分生组织的大小[9]。白菜型油菜中Gα 蛋白基因BraA.Gα1在花和茎顶端分生组织中高表达[10]。小G 蛋白是一类单体G 蛋白,而ROP(Rho-related GTPase from plants)是植物中特有的一类小G 蛋白,参与调控植物生命活动的众多生理过程[11,12]。拟南芥中共有11 种ROP,它们参与调控细胞极性的建立和维持[13,14]、细胞骨架的组织动态与囊泡运输[15~17]、响应激素信号[18,19]、保卫细胞的开放和闭合[20]、植物防御病原体的反应[21,22]等重要的生命活动。蛋白免疫共沉淀试验发现拟南芥ROP蛋白可与CLV1 蛋白相结合形成复合物,调控植物顶端分生组织的发育[23,24]。此外,研究表明ROPs 活性因子基因RopGEF7B参与调控水稻花器官发育,当RopGEF7B被T-DNA 插入以及RNAi 干涉以后,突变体均表现出第三轮和第四轮花器官(雄蕊和子房)数量增加的现象[25],类似于水稻中FON1(CLV1同源基因)的突变体表型,即雌蕊数量增加的现象[26],表明植物中ROP 家族基因在CLV 信号途径中发挥着重要的作用。
前期研究发现芥菜型油菜J163-4 植株中BjMcl基因(CLV1的同源基因)的突变使其产生4 个心皮[27],且小G 蛋白基因BjROP10的表达量相较于芥菜型两室油菜显著增加。本研究通过CRISPR/Cas9技术构建了BjROP10的双靶点载体,通过遗传转化芥菜型多心皮油菜J163-4,最终获得Bjrop10突变体,并鉴定分析了目标基因的突变位点。本研究为进一步解析BjROP10基因响应CLV 信号调控油菜心皮数发育的功能提供了材料基础,也为研究油菜多心皮性状发育的分子机制提供理论和实践参考。
1 材料与方法
1.1 材料
芥菜型多心皮油菜J163-4 由华中农业大学作物遗传改良国家重点实验室油菜组提供,种植于临沂大学实验田。人工气候培养箱里面培养油菜转基因再生苗,光照为20 000 lx,温度为22℃,黑暗培养8 h/光照培养16 h。人工气候箱培养2周左右后,炼苗,移栽到大田。
1.2 方法
1.2.1 油菜BjROP10的克隆 利用CTAB 法[28]提取芥菜型多心皮油菜J163-4 幼嫩叶片的基因组总DNA。以NCBI 数据库中芥菜型油菜基因组序列为参考,利用Primer3 设计BjROP10的扩增引物,引物序列为:ROP10-1L:5’-ATGGCTTCAAGTGCTTCGAAGTTC-3’和ROP10-1R:5’-CATCATCAATGCCGAGTCACCA-3’。使用Phusion 热启动II 高保真PCR 聚合酶混合物(Phusion Green Hot Start II High-Fidelity PCR Master Mix,Thermo Scientific™)扩增BjROP10,20µL PCR 反应体系包括:基因组DNA模板2 µL、ROP10-1L 引物1 µL、ROP10-1R 引物1µL、ddH2O 6 µL、Phusion 热启动II 高保真PCR 聚合酶混合物10 µL。PCR 反应程序为:94℃3 min,94℃30 s,55℃30 s,72℃1 min,35 个 循环,72℃10 min。PCR 产物利用天根科技有限公司的DNA纯化回收试剂盒回收后,利用TaKaRa 公司的pMDTM18-T Vector Cloning Kit 进行克隆测序。在BRAD(http://brassicadb.cn/)网站分析BjROP10在芥菜型油菜基因组中的拷贝位点及同源基因位点。
1.2.2 CRISPR/Cas9 基因编辑双靶点载体的构建 通过SoftBerry(http://linux1. softberry. com/berry. phtml)预测BjROP10的基因外显子区域和内含子区域。利用NCBIblast 比对获得BjROP10的保守区域。通过CRISPR-P 2.0(http://crispr. hzau. edu.cn/cgi-bin/CRISPR2/CRISPR)在BjROP10外显子的保守区域筛选靶点并设计2 条sgRNA 序列,分别为TD1-GCGCAAACGTTGTAGTTGA(AGG)和TD2-AGAAGACTATAACAGATTA(AGG)。设计4 条PCR引物,引物序列分别为ROP1-TD1-BsF: 5’-ATATATGGTCTCGATTGGCGCAAACGTTGTAGTT -GAGTT-3’,ROP1-TD1-F0: 5’-TGGCGCAAACGTTGTAGTTGAGTTTTAGAGCTAGAAATAGC-3’,ROP1-TD2-R0: 5’ -AACTAATCTGTTATAGTCTTCTCAATCTCTTAGTCGACTCTAC-3’,ROP1-TD2-BsR: 5’-ATTATTGGTCTCGAAACTAATCTGTTATAGTCTTCTCAA-3’。下划线部分为TD1 和TD2 的sgRNA 靶点序列,粗体部分为BsaI 酶识别位点。以pCBC-TD1TD2 为模板,进行四引物PCR,其中ROP1-TD1-BsF 和ROP1-TD2-BsR 引物的浓度为10 µmol/L,ROP1-TD1-F0 和ROP1-TD2-R0引物的浓度为0.01µmol/L,20µL PCR 反应体系包括:pCBC-TD1TD2 模板2 µL、ROP1-TD1-BsF 引物1 µL、ROP1-TD2-BsR 引物1 µL、ROP1-TD1-F0引物1 µL、ROP1-TD2-R0 引物1 µL、ddH2O 4 µL、Phusion 热启动II 高保真PCR 聚合酶混合物10 µL。PCR 反应程序为:94℃3 min;94℃30 s,55℃30 s,72℃1 min,35 个循环;72℃10 min。纯化回收PCR产物,建立10µL 酶切连接体系:PCR 片段(626 bp)2 µL、pKSE401 质粒2 µL、10 × NEB T4 Buffer 1.5µL、10 × BSA 1.5 µL、BsaI(NEB)1 µL、T4 Ligase(NEB)/高浓度1 µL、ddH2O 6 µL。PCR 片段通过BsaI 位点克隆到pSKE401 载体,构建含T1 和T2 两个 靶 点 的 CRISPR/Cas9 载 体 ROP10-TD1TD 2(图1)。
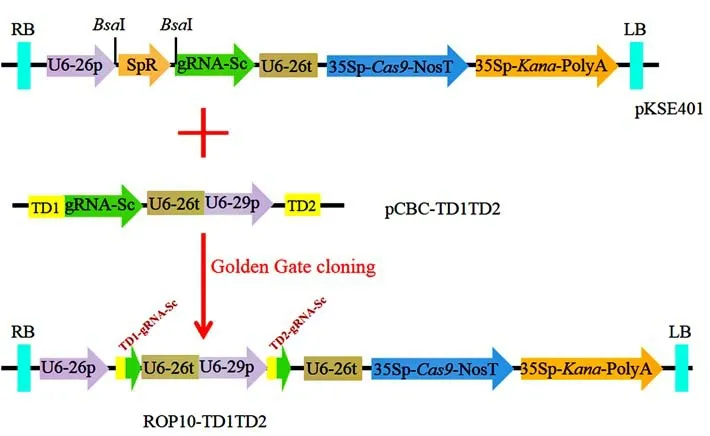
图1 基因编辑载体的构建Fig.1 Construction of genome editing vector
1.2.3 芥菜型油菜遗传转化 将ROP10-TD1TD2载体转化农杆菌GV3101 后,在芥菜型四心皮油菜中进行农杆菌介导的遗传转化,转化过程中使用卡那霉素进行阳性苗的筛选。ROP10-TD1TD2 在农杆菌中的遗传转化参照GV3101 Chemically Competent Cell 产品说明书,芥菜型油菜的组织培养方法参照已公开发布的国家发明专利[28]。
取移栽成活的油菜幼苗叶片,利用CTAB 法取基因组总DNA[29],根据pKSE401 载体上的Cas9基因序 列,设 计PCR 引 物CAS9-1L:5’-TCGCTGCCAAGAATCTGTCGG-3’和CAS9-1R:5’-TCCACGATGGCCTTCTTCTGCT-3’。以油菜幼苗基因组DNA 为模板,通过PCR 进行转基因验证,PCR 反应程序为:94℃3 min;94℃30 s,55℃30 s,72℃1 min,35个循环;72℃10 min。
以ROP10-TD1TD2 转基因油菜基因组DNA 为模 板,ROP10-1L 和ROP10-1R 为 引 物 进 行PCR 扩增。将扩增的DNA 片段进行TA 克隆后,挑选阳性单克隆进行测序,测序结果与野生型基因BjROP10进行BLAST分析,鉴定目标基因的突变位点。
2 结果与分析
2.1 芥菜型油菜中BjROP10及其同源基因序列分析
BLAST 分析发现,BjROP10在芥菜型油菜基因组中存在两个位点,分别为位于B04 染色体的BjuB000002和位于B06 染色体的Bju046507。测序结 果 证 明,BjROP10与BjuB000002和Bju046507的序列完全一致。BjROP10的基因全长为1576 bp,CDS序列长度为651 bp,包含8个外显子和7个内含子,编码包含216个氨基酸的蛋白。此外,测序结果显示BjROP10在芥菜型油菜基因组的A02 染色体上还存在一个同源基因BjuA02_ROP,该同源基因的基因全长为1506 bp,CDS 序列长度为615 bp,包含6个外显子和5 个内含子,编码包含204 个氨基酸的蛋白。BjROP10与BjuA02_ROP的CDS 序列存在27个位点的单碱基差异,且由于序列变异,BjuA02_ROP的CDS 序列相比BjROP10缺失最后一个外显子(图2)。为了进一步研究BjROP10的功能,且不受同源基因的干扰,分别在其第二个和第三个外显子位置(BjROP10在该位置的序列与BjuA02_ROP存在单碱基位点的差异)设计2 个sgRNA 靶点TD1 和TD2(图3),用于CRISPR/Cas9载体的构建。

图2 BjROP10和BjuA02_ROP的蛋白质序列比对Fig.2 Comparision of protein sequences between BjROP10 and BjuA02_ROP

图3 芥菜型油菜BjROP10的基因组和CDS序列分析及基因组编辑载体靶点的选择Fig.3 Alignment of genomic and CDS sequence of BjROP10 and the selection of CRISPR target sites in Brassica juncea
2.2 油菜遗传转化及转基因植株的获得
利用CRISPR 载体ROP10-TD1TD2 通过农杆菌介导遗传转化油菜下胚轴(图4A-D)。经过含卡那霉素的培养基筛选,共获得22株油菜再生植株。取油菜再生植株幼叶提取基因组DNA,利用Cas9序列引物,通过PCR 对转基因油菜植株进行验证发现,其中20 株为转基因阳性植株(图4E),转基因的阳性率约为91%。
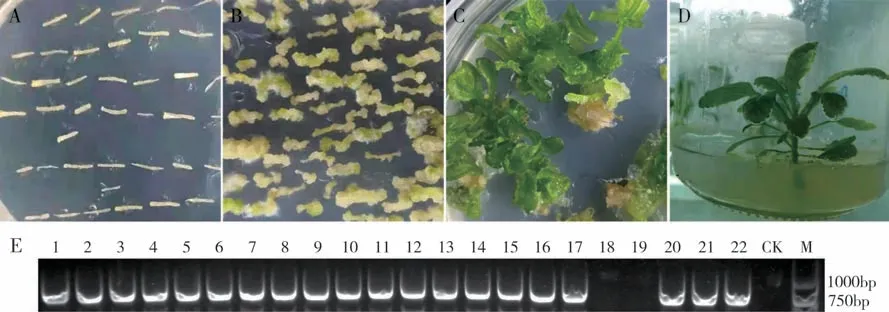
图4 ROP10-TD1TD2在芥菜型多心皮油菜J163-4中的遗传转化及遗传转化再生苗的PCR鉴定Fig.4 Agrobacterium-mediated genetic transformation of ROP10-TD1TD2 in J163-4 of multilocular Brassica juncea and PCR identification of regenerated seedlings of genetic transformation
2.3 转基因油菜心皮表型的鉴定及BjROP10 基因编辑位点的鉴定
在20 株油菜转基因阳性植株中,有13 株心皮数目发生改变,即角果由原来的4 心皮性状变为2心皮、3 心皮或者5 心皮性状(图5)。其中,2 株表现出2 心皮、3 心皮、4 心皮、5 心皮角果并存的现象,4株表现出3 心皮和4 心皮角果并存的现象,5 株表现出4 心皮和5 心皮角果并存的现象,有2 株表现出3心皮、4 心皮和5 心皮角果并存的现象,而对照组中的3株非转基因植株J163-4-1、J163-4-2、J163-4-3均表现为4心皮角果(表1)。

图5 芥菜型油菜不同心皮数角果Fig.5 Siliques with different carpel numbers in Brassica juncea
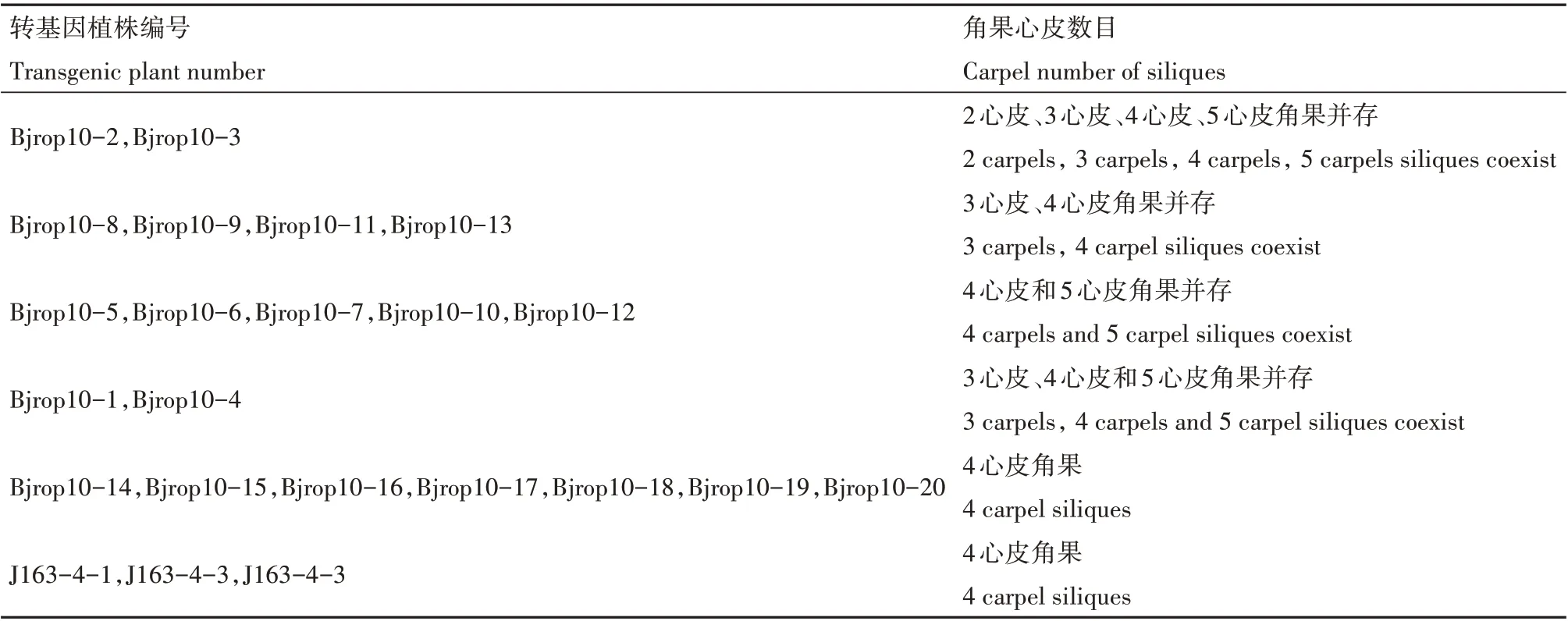
表1 转基因植株的角果性状Table 1 Silique traits of transgenic plants
为进一步鉴定基因组编辑载体ROP10-TD1TD2 对BjROP10的突变情况,在T0植株中,测序分析了7 株心皮数发生变化的转基因油菜,编号分别为Bjrop10-1~Bjrop10-7。结果显示,这7 株的BjROP10基因序列均发生变异,且变异序列均位于BjROP10第481-1283 个核苷酸之间的T1 和T2 靶点位置。其中,7 株植株在T1 靶点处均发生了序列变异,Bjrop10-2、Bjrop10-3、Bjrop10-4、Bjrop10-6 和Bjrop10-7 在T2 靶点处也发生了序列变异。除Bjrop10-3 植株仅检测到一个突变拷贝外,其它6 个转基因植株均检测到两个突变拷贝(图6)。经过编辑单株的基因型鉴定发现,Bjrop10-3植株为杂合突变,即BjROP10的1 个拷贝发生突变,而另外一个拷贝 没 有 发 生 突 变。 而Bjrop10-1、Bjrop10-2、Bjrop10-4、Bjrop10-5、Bjrop10-6 和Bjrop10-7 植株均发生了嵌合突变,即BjROP10的两个拷贝均发生了不同的序列变异,且在转基因植株中没有检测到BjROP10的野生型拷贝。
如图6 所示,Bjrop10-1 中的拷贝Bjrop10-1-1在T1 位点缺失1 个T 碱基,导致移码突变且翻译提前终止,拷贝Bjrop10-1-2 在T1 位点插入1 个T 碱基,导致该位点形成终止密码子UGA,翻译提前终止,而且第510位氨基酸由T碱基替换为C碱基为同义突变,突变前的密码子AUU 和突变后的密码子AUC 均编码异亮氨酸(Ile);Bjrop10-2 中,拷贝Bjrop10-2-1 在T1 和T2 靶点之间的序列发生了大片段的变异,导致第36-101 个氨基酸缺失,拷贝Bjrop10-2-2 在T1 和T2 靶点之间的序列发生了大片段的缺失,导致第52-101 个氨基酸发生移码突变;Bjrop10-3 中检测发现仅有一种变异拷贝,即在T1 靶点位置序列TG 突变为AACAGGACTACATACCAAAAGC 序列,且T2 靶点位置发生575 bp 的大片段缺失,但是T1靶点位置序列的变异产生终止密码子UGA,导致翻译的提前终止。 Bjrop10-4、Bjrop10-5 和Bjrop10-6 植株中的BjROP10拷贝均由于T1 靶点位置序列的缺失导致了移码突变及翻译提前终止。Bjrop10-7植株中的拷贝Bjrop10-7-1由于T1 靶点位置序列的大片段缺失,导致第44-71 个氨基酸发生移码突变,拷贝Bjrop10-7-2 由于T1 靶点位置序列的缺失导致移码突变及翻译提前终止。以上突变最终导致BjROP10 蛋白的结构被破坏(图7),转基因植株的心皮数目发生改变。

图6 不同转基因植株中BjROP10基因序列的变异分析Fig.6 Variation analysis of BjROP10 gene sequences in different transgenic plants

图7 不同转基因植株中Bjrop10的蛋白序列Fig.7 Protein sequences of Bjrop10 in different transgenic plants
3 讨论与结论
在油菜育种中,多心皮性状是一种有着潜在利用价值的优异的高产性状。近年来,已有较多的研究探讨其形成的分子机制。目前在三大类型油菜(白菜型油菜、芥菜型油菜和甘蓝型油菜)中均发现了多心皮性状。已有研究认为CLV 信号途径相关基因调控了油菜多心皮性状的形成,例如,控制白菜型油菜多心皮性状的BrCLV3是拟南芥CLV3的同源基因[4],而控制芥菜型油菜的多心皮性状的两个基因BjMc1和BjCLV1均为拟南芥CLV1的同源基因[5,27,30];将甘蓝型油菜中CLV 信号途径的三个重要成员CLV1、CLV2和CLV3的同源基因进行基因编辑后,均获得了多心皮性状[31]。本研究中,芥菜型多心皮油菜J163-4 为CLV1同源基因的突变体,其为四心皮性状,且没有嵌合现象,是当前报道中唯一没有嵌合现象的多心皮油菜突变体[32],也是研究油菜多心皮性状形成的分子机制的优异材料。通过基因编辑技术将J163-4 中的BjROP10基因突变以后,突变体植株的心皮数发生变化,形成了两心皮角果、三心皮角果、四心皮角果以及五心皮角果,证明芥菜型油菜中小G 蛋白基因参与CLV 途径调控其心皮数的发育,但是究竟是通过何种分子机制进行调控,小G 蛋白基因BjROP10和CLV1存在怎样的互作关系,还需要进一步的研究。本研究中所获得的突变体为后续实验提供了材料基础。另外,在本研究中将小G 蛋白基因BjROP10突变后,突变体植株出现了多种心皮数目的现象,即嵌合现象,推测由于本研究调查的是基因编辑的T0突变体,同一个突变体植株中存在多个BjROP10基因的突变拷贝,从而导致了同一植株出现多种心皮数目的现象。后续试验中,将观察T0代自交后代的表型并鉴定其突变基因型,从而确定小G 蛋白基因BjROP10的各种突变拷贝对油菜心皮数发育的影响,这将为油菜多心皮发育的分子机制的研究奠定理论和实践基础。
CRISPR/Cas9 基因编辑技术能够快速准确地修饰基因组位点,在作物分子育种领域有着较高的发展潜力。当前,花生中通过CRISPR/Cas9 技术编辑了FAD2基因,获得了其突变体,为高油酸花生育种提供了育种材料[33];玉米中通过CRISPR/Cas9 技术对香味基因BADH2进行编辑,最终获得了具有香味的玉米突变体,填补了当前玉米种质资源库没有香味玉米品种的空白[34]。水稻中通过CRISPR/Cas9 技术对香味基因Badh2和粒长基因GS3进行了定向编辑,成功获得了长粒型香米品种[35];此外,大豆中通过CRISPR/Cas9技术编辑了与重金属转运相关的蛋白基因GmHIPP26,获得了目标基因的突变体,为研究GmHIPP26在大豆镉胁迫中的功能提供了实验材料[36]。本研究中也通过CRISPR/Cas9 基因编辑技术获得了油菜的小G 蛋白基因Bjrop10突变体植株,序列分析显示BjROP10的突变主要为碱基的插入和缺失,且多个突变拷贝存在大片段缺失的现象,其中突变拷贝Bjrop10-2-2、Bjrop10-3 和Bjrop10-6-1中,T1 和T2 靶点之间的序列全部缺失,突变拷贝Bjrop10-2-1 的T1 和T2 靶点之间的序列出现了大片段变异,此外,本研究中BjROP10其它的突变拷贝也均在T1 或T2 处存在碱基的替换或缺失,证明本研究中使用的CRISPR/Cas9双靶点系统的基因编辑能力很强。这将为后期油菜的分子育种提供实践基础和技术支撑,所获得的突变体材料也将为油菜的多心皮育种提供新的种质资源。
