紫花苜蓿与不同生活型多年生禾本科牧草混播生长生理特征
2022-08-31张辉辉师尚礼李自立李小龙康文娟陈新栋
张辉辉, 师尚礼, 武 蓓, 李自立, 李小龙, 吴 芳, 康文娟, 陈新栋
(甘肃农业大学草业学院, 草业生态系统教育部重点实验室, 中-美草地畜牧业可持续研究中心, 甘肃 兰州 730070)
以单一作物耕作生产的传统农业是一种生产力低、不可持续的生产方式,用粮饲兼顾的草地农业可发挥4倍于土地资源传统耕作农业利用的潜力[1]。应用优良的豆科牧草与禾本科牧草混播建植人工草地能协同发挥豆科牧草和禾本科牧草的适应性和抗逆性[2],能再次提高自然资源利用率,达到增产[3-6]、提质[7-9]及改土培肥[10-13]的目的。上述混播优势在生产实践中能否持续存在,是豆禾混播系统具备优良生产性能的关键。在豆禾混播系统中草种组分对水分、养分、光资源和生态位的竞争以及高秆草种对矮杆草种的遮荫作用会使混播系统组分间产生逆境胁迫,牧草所受逆境胁迫超过其可调节范围时会限制各组分牧草的生长[14-15]。在混播系统中,植物所受到的逆境胁迫主要来自种间竞争,当受到逆境胁迫时,植物会通过调节自身的抗氧化物酶活性来清除体内积累的活性氧和自由基,减小氧化损伤[16],增加渗透物质含量以减轻或避免丙二醛等有害物质积累造成的组织或器官生物膜系统功能和结构的伤害,通过提高抗氧化能力和渗透调节能力,达到种间生理协同[17],同时,混播系统通过协调植物光合生理来协调种间对光资源的利用,达到提高干物质积累的目的[2]。
杨志超等[15]在紫花苜蓿分别与无芒雀麦、垂穗披碱草(Elymusnutans)、虉草(Phalarisarundinacea)和猫尾草(Phleumpratense)不同比例建植混播草地的研究中发现,混播组合和成分比例显著影响各组分牧草抗氧化酶活性和抗氧化剂含量。黄丽琴等[18]对苜蓿与虉草混播草地的研究发现,虉草根系可溶性糖含量在不同混播比例下均高于单播,在豆禾间行比为2∶1时最高;苜蓿根系可溶性糖含量在豆禾间行比为1:1时最高,董世魁等[19]发现混播草地光能利用率较单播显著提高。在豆禾混播系统中,生理调节是牧草响应竞争环境的主要生存策略,目前对混播草地牧草生理响应混播草种和比例的研究较少。本研究以紫花苜蓿分别与3类不同生活型多年生禾本科牧草建植的混播草地为研究对象,通过比较分析不同混播组合和成分比例下牧草抗氧化能力、膜脂过氧化物含量、渗透调节物质含量以及光合生理的差异,探讨牧草生长生理对混播系统种间协同效应的响应,为混播草种搭配及比例设置提供参考。
1 材料与方法
1.1 试验区自然概况
试验地位于甘肃农业大学兰州牧草实训基地,地理位置105°41′ E,34°05′ N,地处黄土高原西端向青藏高原过渡地带,海拔1 595 m,属温带半干旱大陆性气候,年平均气温10.3℃,年平均日照时数2 374 h,无霜期172 d,年降水量300 mm左右,集中分布于6—9月,年均蒸发量1 664 mm。土壤为黄绵土,肥力均匀,有机质含量为0.96%,碱解氮含量为65.92 mg·kg-1,速效钾含量为104.21 mg·kg-1,速效磷含量为10.06 mg·kg-1。
1.2 供试材料
供试紫花苜蓿品种为‘清水’(Medicagosativa‘Qingshui’),发芽率86.67%,由甘肃农业大学教育部草业生态系统重点实验室提供;不同生活型禾本科材料:丛生型-苇状羊茅,品种为‘雨姿’(Festucaarundinacea‘Yuzi’),发芽率85.56%,匍匐茎型-草地早熟禾,品种为‘盖世’(Poapratensis‘Gaishi’),发芽率81.11%,由甘肃创绿草业科技有限公司提供;根茎型-无芒雀麦,品种为‘原野’(Bromusinermis‘Yuanye’),发芽率82.22%,由北京正道草业有限公司提供。
1.3 试验设计
试验设3个混播组合,每个混播组合设3个混播比例(混播播种量按占单播量的比例计算),另设置4个单播处理作为对照(表1),共13个处理,小区面积15 m2(5 m×3 m),重复3次,总共39个小区,随机区组排列,小区间距0.4 m,行距0.2 m,采用同行条播的播种方式,播种深度2 cm。播种时间2020年5月25日,底肥施用量:磷酸二铵(P2O5≥46%)120 kg·hm-2;尿素(总氮≥46%)80 kg·hm-2;硫酸钾(K2O≥51%)50 kg·hm-2,底肥播种前均匀撒布于地表,翻耕入土,整地种植。试验地采用滴灌方式进行灌溉,生长期间不追肥,采用人工除草。2020年5月25日播种,2020年8月18日和2021年6月2日紫花苜蓿初花期取样并测定相关指标。

表1 试验处理Table 1 Experimental treatments
1.4 测定指标与方法
单播区每小区取3个1 m样段,称鲜重,然后每个样段取500 g鲜样;混播区每小区取3个1 m样段,取样后豆禾分开称鲜重,每个样段豆禾各取鲜样500 g;鲜样带回实验室后于105℃下杀青20 min,80℃下烘干至恒重,2年数据取平均值,通过测产面积和草样干鲜比换算获得混播草地产量,并根据产量计算各组分增产贡献率:
式中:YIR表示混播系统增产率,Yij和Yji分别表示混播系统中豆科牧草和禾本科牧草的产量,Yi和Yj分别表示豆科牧草和禾本科牧草的单播产量,Pi和Pj分别表示豆科牧草和禾本科牧草的混播比例,且Pi+Pj=1。

株高:用卷尺测量任意一株牧草从地面至叶尖或花序顶端的绝对高度,10株为一个重复,取其平均值,共3次重复。
紫花苜蓿初花期时于晴天9:30—11:00采用GFS-3000便携式光合仪(Walz,Germany)对紫花苜蓿功能叶片倒4叶的光合气体交换参数进行测定,空气中CO2浓度固定为400 μmol·mol-1,光照强度设定为1400 μmol·(m2·s)-1;对草地早熟禾顶第2片完全展开的功能叶的气体交换参数进行测定,空气中CO2浓度固定为400 μmol·mol-1,光照强度设定为1 000 μmol·(m2·s)-1;对无芒雀麦顶端第2片完全展开的功能叶的气体交换参数进行测定,空气中CO2浓度固定为400 μmol·mol-1,光照强度设定为2 000 μmol·m-2·s-1;对苇状羊茅顶端第2片完全展开的功能叶的气体交换参数进行测定,空气中CO2浓度固定为400 μmol·mol-1,光照强度设定为800 μmol·m-2·s-1;测定指标包括:净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。
于紫花苜蓿初花期随机选取单株,剪取从顶部向下第3~5片完整叶,每个小区10个重复单株。将取好的叶片装入锡箔纸袋在液氮中迅速冷冻,带回实验室后于-80℃超低温冰箱储藏[20],用于测定指标:超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)光还原法测定[21],过氧化物酶(POD)活性采用愈创木酚法测定[22],过氧化氢酶(CAT)活性采用试剂盒法(试剂盒由苏州格锐思生物公司提供),丙二醛(MDA)含量采用硫代巴比妥酸法测定[23],脯氨酸(Pro)含量采用酸性茚三酮法测定[24],可溶性糖(SS)含量采用蒽酮法测定[25],可溶性蛋白含量(SP)采用考马斯亮蓝染色法[26],并用牛血清白蛋白作标准曲线。
1.5 数据分析
采用Excel2010进行数据统计、计算及隶属函数分析,SPSS(IMB SPSS Statiatics 19)进行数据单因素方差分析,并用Duncan法对各测定数据进行多重比较。
利用下列公式计算不同混播处理标准化数据综合指标的隶属函数值,与植物生长生理协同能力呈正相关的参数,如SOD,POD,CAT,Pro,SS和SP等采用公式:
u(Xj)=(Xj-Xmin)/(Xmax-Xmin),
j=1,2,3,…,n
与植物生长生理协同能力呈负相关的参数如MDA采用公式:
u(Xj)=1-(Xj-Xmin)/(Xmax-Xmin),
j=1,2,3,…,n
式中,Xj表示第j个综合指标,u(Xj)表示j个综合指标的隶属函数值,Xmax和Xmin分别表示第j个综合指标的最大值和最小值。
权重采用客观赋权法进行计算:
式中,Wj表示第j个综合指标的权重,Pj表示第j个综合指标的贡献率。
综合评价D值计算公式:
2 结果与分析
2.1 混播成分与比例对牧草产量的影响
不同混播处理下牧草产量如表2所示。混播草地产量均较单播草地显著提高(P<0.05),随紫花苜蓿混播比例的下降和禾草混播比例的上升,紫花苜蓿产量下降,禾草产量上升,混播草地总产量下降。不同处理的混播草地各组分增产率和混播系统增产率均为正值,较单播草地增产。紫花苜蓿与草地早熟禾P1处理下紫花苜蓿增产率最大,P2处理下草地早熟禾增产率最大;紫花苜蓿与无芒雀麦B2处理下紫花苜蓿增产率最大,B3处理下无芒雀麦增产率最大;紫花苜蓿与苇状羊茅P1处理下紫花苜蓿与苇状羊茅增产率均最大。

表2 不同混播处理下牧草产量的差异Table 2 Effect of different mixed sowing treatment on yield of forage
2.2 混播成分与比例对牧草株高的影响
不同混播处理下牧草株高如图1所示,在紫花苜蓿与草地早熟禾混播草地中,混播紫花苜蓿株高2020年和2021年分别为75.94~77.51 cm和86.44~101.64 cm,在两年试验各混播处理下均显著低于单播(P<0.05);混播草地早熟禾株高2020年和2021年分别为40.33~44.78 cm和49.83~58.13 cm,各年份混播处理均高于单播,其中在2020年各混播处理及2021年P1处理下显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,混播紫花苜蓿株高2020年和2021年分别为68.07~72.17 cm和85.94~99.04 cm,在两年各混播处理下均显著低于单播(P<0.05);混播无芒雀麦株高2020年和2021年分别为44.17~47.83 cm和49.83~58.13 cm,在2020年B1和B2处理以及2021年各处理下显著高于单播(P<0.05)。在紫花苜蓿与苇状羊茅混播草地中,混播紫花苜蓿株高2020年和2021年分别为74.22~77.75 cm和85.42~96.99 cm,在两年F2和F3处理下均显著低于单播(P<0.05);混播苇状羊茅株高2020年和2021年分别为68.52~72.63 cm和82.70~92.40 cm,在两年各处理下显著高于单播(P<0.05)。
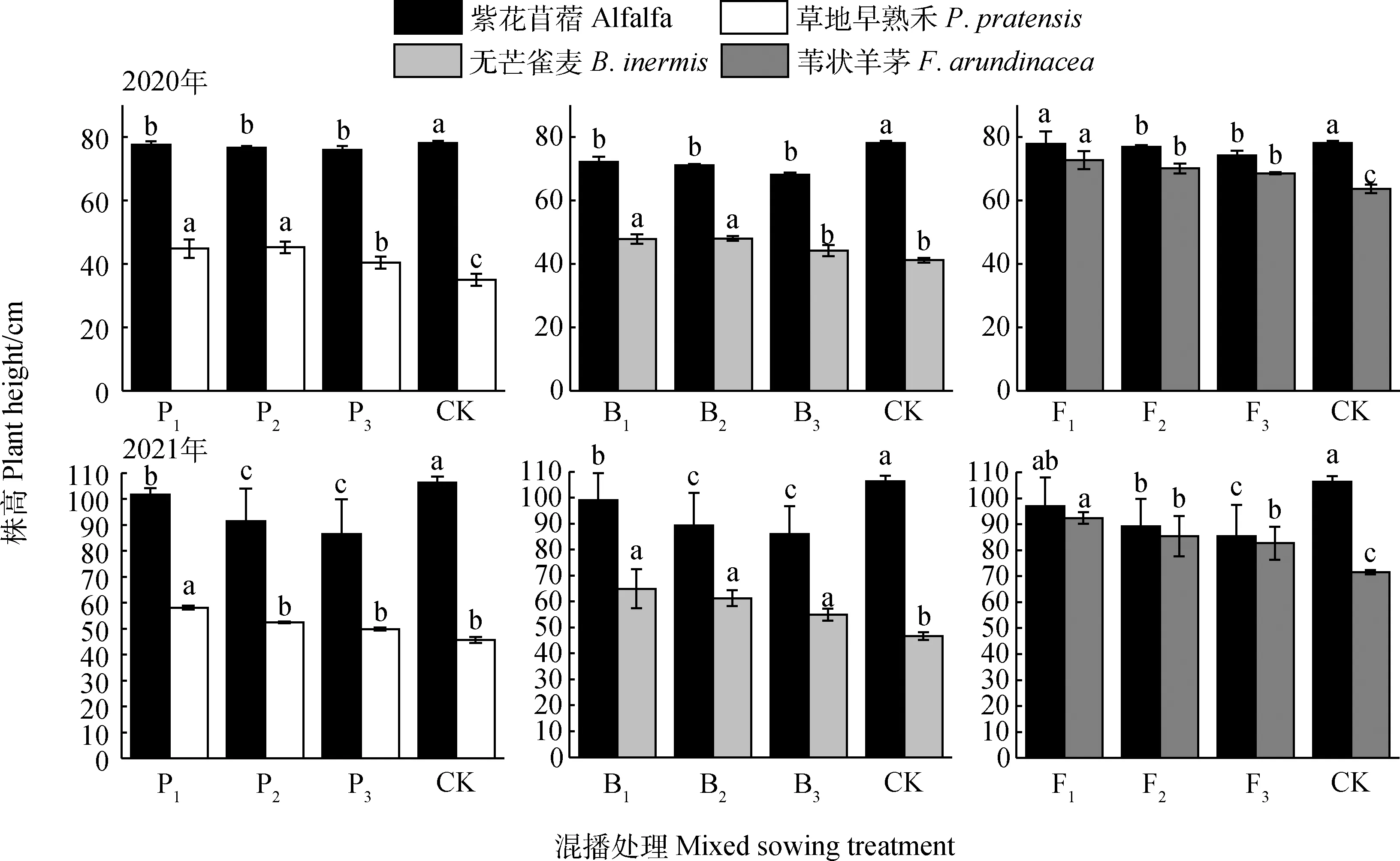
图1 不同混播处理对牧草株高的影响Fig.1 Effects of different mixed sowing treatment on plant height of forage
2.3 混播成分与比例对牧草叶片膜脂过氧化物含量的影响
如图2所示,不同混播处理下牧草叶片MDA含量具有差异。在紫花苜蓿与草地早熟禾混播草地中,紫花苜蓿叶片MDA含量在两年试验各混播处理下均显著低于单播(P<0.05);草地早熟禾叶片MDA含量在两年各混播处理下均显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,紫花苜蓿叶片MDA含量在两年试验P2和P3处理下均显著低于单播(P<0.05);无芒雀麦叶片MDA含量在2020年B2和B3处理下显著高于单播(P<0.05),在2021年各处理与单播差异不显著。在紫花苜蓿与苇状羊茅混播草地中,紫花苜蓿叶片MDA含量在2020年各处理以及2021年F2和F3处理下显著低于单播(P<0.05);苇状羊茅叶片MDA含量在2020年F2和F3处理以及2021年各处理下显著低于单播(P<0.05)。随紫花苜蓿比例的下降和禾草比例的上升,紫花苜蓿与3种禾草叶片MDA含量均呈下降趋势。

图2 不同混播处理对叶片丙二醛含量的影响Fig.2 Effects of different mixed sowing treatment on malondialdehyde content of leaf
2.4 混播成分与比例对牧草叶片抗氧化酶活性的影响
不同混播处理下牧草叶片SOD酶活性如图3所示。在紫花苜蓿与草地早熟禾混播草地中,紫花苜蓿叶片SOD酶活性在2020年P1和P2处理以及2021年各处理下均显著高于单播(P<0.05);草地早熟禾叶片SOD酶活性在2020年P2和P3处理以及2021年P1处理下显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,紫花苜蓿叶片SOD酶活性在2020年各处理及2021年P1和P2处理下均显著高于单播(P<0.05);无芒雀麦叶片SOD酶活性在2020年B2和B3处理以及2021年各处理下显著高于单播(P<0.05)。在紫花苜蓿与苇状羊茅混播草地中,紫花苜蓿叶片SOD酶活性在2020年各处理以及2021年F1和F2处理下显著高于单播(P<0.05);苇状羊茅叶片SOD酶活性在两年F1和F2处理下显著高于单播(P<0.05)。
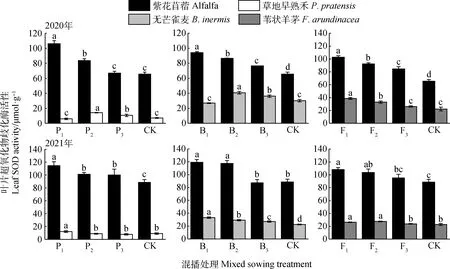
图3 不同混播处理对叶片超氧化物歧化酶活性的影响Fig.3 Effects of different mixed sowing treatment on SOD activity of leaf
如图4所示,在紫花苜蓿与草地早熟禾混播草地中,紫花苜蓿叶片POD酶活性在2020年P1和P2处理以及2021年各处理下均显著高于单播(P<0.05);草地早熟禾叶片POD酶活性在2020年P2和P3处理以及2021年P1处理下显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,紫花苜蓿叶片POD酶活性在2020年各处理及2021年P1和P2处理下均显著高于单播(P<0.05);无芒雀麦叶片POD酶活性在2020年B2和B3处理以及2021年各处理下显著高于单播(P<0.05)。在紫花苜蓿与苇状羊茅混播草地中,紫花苜蓿叶片POD酶活性在2020年各处理以及2021年F1和F2处理下显著高于单播(P<0.05);苇状羊茅叶片POD酶活性在两年F1和F2处理下显著高于单播(P<0.05)。
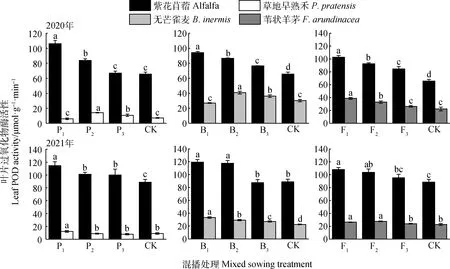
图4 不同混播处理对叶片过氧化物酶活性的影响Fig.4 Effects of different mixed sowing treatment on POD activity of leaf
如图5所示,在紫花苜蓿与草地早熟禾混播草地中,紫花苜蓿叶片CAT酶活性在2020年各处理以及2021年P1和P2处理下均显著高于单播(P<0.05);草地早熟禾叶片CAT酶活性在2020年各处理以及2021年P1和P2处理下显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,紫花苜蓿叶片CAT酶活性在2020年B1和B2处理及2021年各处理下均显著高于单播(P<0.05);无芒雀麦叶片CAT酶活性在2020年B2处理以及2021年各处理下显著高于单播(P<0.05)。在紫花苜蓿与苇状羊茅混播草地中,紫花苜蓿叶片CAT酶活性在2020年F1处理以及2021年各处理下显著高于单播(P<0.05);苇状羊茅叶片CAT酶活性在2020年F1和F2处理以及2021年各处理下显著高于单播(P<0.05)。

图5 不同混播处理下叶片过氧化氢酶活性的差异Fig.5 Effects of different mixed sowing treatment on CAT activity of leaf
总体而言,混播处理下多组分刺激了SOD酶活性、POD酶活性及CAT酶活性的变化,改变了植物清除体内活性氧的能力,以响应组分间竞争。
2.5 混播成分与比例对牧草叶片渗透调节物质含量的影响
如图6所示,不同混播处理下牧草叶片Pro含量具有差异。在紫花苜蓿与草地早熟禾混播草地中,紫花苜蓿叶片Pro含量在2020年各处理以及2021年P1和P2处理下均显著高于单播(P<0.05);草地早熟禾叶片Pro含量在2020年P1和P2处理以及2021年各处理下显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,紫花苜蓿叶片Pro含量在2020年各处理及2021年B1和B2处理下均显著高于单播(P<0.05);无芒雀麦叶片Pro含量在两年各处理下显著高于单播(P<0.05)。在紫花苜蓿与苇状羊茅混播草地中,紫花苜蓿叶片Pro含量在2020年F1和F2处理以及2021年各处理下显著高于单播(P<0.05);苇状羊茅叶片Pro含量在2020年F1处理以及2021年F1和F2处理下显著高于单播(P<0.05)。
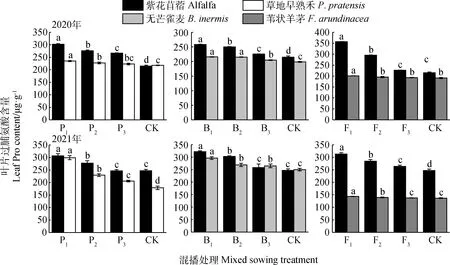
图6 不同混播比例处理下叶片脯氨酸含量的差异Fig.6 Effects of different mixed sowing ratio treatment on Pro content of leaf
如图7所示,在紫花苜蓿与草地早熟禾混播草地中,紫花苜蓿叶片SS含量在2020年P1和P2处理以及2021年P1处理下均显著高于单播(P<0.05);草地早熟禾叶片SS含量在2020年P2处理以及2021年P1处理下显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,紫花苜蓿叶片SS含量在两年B1和B2处理下均显著高于单播(P<0.05);无芒雀麦叶片SS含量在两年B1处理下显著高于单播(P<0.05)。在紫花苜蓿与苇状羊茅混播草地中,紫花苜蓿叶片SS含量在2020年F1处理以及2021年F1和F2处理下显著高于单播(P<0.05);苇状羊茅叶片SS含量在2021年F1处理下显著高于单播(P<0.05)。

图7 不同混播比例处理下叶片可溶性糖含量的差异Fig.7 Effects of different mixed sowing ratio treatment on SS content of leaf
如图8所示,在紫花苜蓿与草地早熟禾混播草地中,紫花苜蓿叶片SP含量在2020年P1和P2处理以及2021年各处理下均显著高于单播(P<0.05);草地早熟禾叶片SP含量在2020年P2处理以及2021年P1和P2处理下显著高于单播(P<0.05)。在紫花苜蓿与无芒雀麦混播草地中,紫花苜蓿叶片SP含量在两年各处理下均显著高于单播(P<0.05);无芒雀麦叶片SP含量在2021年B1处理以及2021年B1和B2下显著高于单播(P<0.05)。在紫花苜蓿与苇状羊茅混播草地中,紫花苜蓿叶片SP含量在两年各处理下显著高于单播(P<0.05);苇状羊茅叶片SP含量在2021年F1处理下显著高于单播(P<0.05)。

图8 不同混播比例处理下叶片可溶性蛋白含量的差异Fig.8 Effects of different mixed sowing ratio treatment on SP content of leaf
总体而言,混播较单播相比,多组分刺激了Pro含量、SS含量及SP含量的变异,调节了细胞渗透势平衡,以响应组分间激烈的竞争。
2.6 混播成分与比例对牧草光合生理的影响
不同混播处理下牧草光合生理变化如表3所示,不同混播处理下牧草光合生理具有差异。各混播处理下紫花苜蓿净光合速率、蒸腾速率、气孔导度和胞间CO2浓度均显著大于单播处理(P<0.05)。草地早熟禾净光合速率在两年P1混播处理下均显著小于单播(P<0.05),在2020年P2和P3混播处理以及2021年P2处理下显著大于单播(P<0.05);草地早熟禾蒸腾速率在2020年P1混播处理下显著小于单播(P<0.05),在两年P2和P3混播处理下均显著大于单播(P<0.05);草地早熟禾气孔导度在2020年P2和P3混播处理以及2021年P2混播处理下显著大于单播(P<0.05),在2021年P1混播处理下显著小于单播(P<0.05);草地早熟禾胞间CO2浓度在两年P2和P3混播处理下均显著大于单播(P<0.05),在P1混播处理下与单播差异不显著。无芒雀麦净光合速率在2020年B2和B3混播处理以及2021年各混播处理下均显著大于单播(P<0.05);无芒雀麦蒸腾速率在两年各混播处理下显著大于单播(P<0.05);无芒雀麦气孔导度在2020年B2和B3混播处理以及2021年各混播处理下显著大于单播(P<0.05);无芒雀麦胞间CO2浓度在2020年各混播处理以及2021年B2处理下显著大于单播(P<0.05)。苇状羊茅净光合速率在2020年F1和F2混播处理以及2021年各混播处理下均显著大于单播(P<0.05);苇状羊茅蒸腾速率在两年各混播处理下显著大于单播(P<0.05);苇状羊茅气孔导度在2020年F1和F2混播处理以及2021年各混播处理下显著大于单播(P<0.05);苇状羊茅胞间CO2浓度在2020年F1混播处理以及2021年各处理下显著大于单播(P<0.05)。

表3 不同混播处理下牧草光合指标的差异Table 3 Effects of different mixed sowing treatment on photosynthetic index of forage
2.7 紫花苜蓿与3种生活型禾草混播牧草生长生理指标综合评价
混播草地牧草通过调节多种生理物质来提高生理协同效应,单一的生理指标难以准确反映牧草的生理协同能力,本研究采用隶属函数法对紫花苜蓿与禾草的生长生理指标进行综合评价,综合评价D值越大,说明牧草生长生理表现越好,结果如表4所示。根据综合评价D值对各混播及单播处理牧草生理排序,结果显示混播紫花苜蓿综合评价D值均大于单播,综合评价D值由大到小的排列为F1>B2>P1=B1>F2>P2>B3>P3=F3>CKM,混播草地早熟禾综合评价D值大于单播,综合评价D值由大到小排列为P2>P3>P1>CKP,混播无芒雀麦综合评价D值在混播处理下均大于单播,综合评价D值由大到小排列为B2>B3>B1>CKB,苇状羊茅综合评价D值在混播处理下均大于单播,综合评价D值由大到小排列为F1>F2>F3>CKF。

表4 不同混播处理下牧草各生长生理指标隶属函数值及综合评价Table 4 Subordinate function values and comprehensive evaluation values of various mixed pasture treatments

续表4
3 讨论

MDA是植物体内自由基作用于膜脂而发生过氧化反应的主要产物,是加剧生物膜损伤的主要物质,其含量的高低可以间接反映膜脂过氧化程度[32]。已有研究表明逆境胁迫增加了植物体内MDA的含量,其含量与胁迫程度有关[33]。本研究发现,在两年试验中,紫花苜蓿与3种多年生禾草混播时紫花苜蓿叶片MDA含量均较单播降低,草地早熟禾和无芒雀麦叶片MDA含量均较单播增加,苇状羊茅叶片MDA含量均较单播降低,根据前人研究[34]可以看出混播紫花苜蓿和苇状羊茅所受逆境胁迫小于单播,混播草地早熟禾和无芒雀麦受到逆境胁迫大于单播。同时,本研究发现随着紫花苜蓿混播比例的减小和禾草混播比例增大,紫花苜蓿的MDA含量呈下降的趋势,推断混播紫花苜蓿种内对自然资源和生态位的竞争大于种间;草地早熟禾和无芒雀麦的MDA含量随这种混播比例的变化呈下降的趋势,推断在混播草地中株高较高的紫花苜蓿对光资源的竞争力大于地早熟禾和无芒雀麦,草地早熟禾和无芒雀麦在光资源竞争中处于劣势,同时矮秆、下繁草的草地早熟禾和无芒雀麦受到高秆、上繁草紫花苜蓿的遮荫作用,造成一定的逆境胁迫[34],随紫花苜蓿混播比例的减小,紫花苜蓿的竞争力减小,遮荫作用减小,草地早熟禾所受逆境胁迫减小,MDA含量下降,符合陈意兰等[35]研究发现的MDA含量随胁迫程度的增加而增加的结果,与付娟娟等[36]发现的遮荫程度越强,禾草所受逆境胁迫越大的结果一致。混播苇状羊茅叶片MDA含量随这种比例变化呈下降的趋势但始终低于单播处理,可能是因为紫花苜蓿与草地早熟禾株高差异较小,苇状羊茅不受紫花苜蓿遮荫竞争造成的压迫,两草种种间对资源的协同性良好,与秦燕[37]研究发现的燕麦与箭筈豌豆在2∶1混作下两种牧草对资源协同性最好的研究结果相似,但因草种不同而协同比例存在差异。混播草地牧草受到的逆境胁迫来源于多方面,本研究只对牧草是否受到逆境胁迫进行了分析,对于造成逆境胁迫的具体原因需要日后进一步探讨。
虽然ROS的累积能引起膜脂过氧化,进而破坏生物膜及细胞正常的生理过程,但生物体内存在抗氧化系统,主要由SOD,POD和CAT组成,能有效清除ROS[38],使得混播牧草种间生长生理达到正常水平。SOD是抗氧化系统清除ROS的核心酶,主要功能是催化超氧化物的歧化反应,将OH-和O2-转化形成H2O2,再由POD和CAT进一步将其转化为H2O[39-40]。吴明浩[41]以苜蓿分别与3种禾草以不同比例混播后发现,当豆禾比为1∶2时,禾草叶片中SOD和CAT活性显著高于其他处理,丙二醛含量低于其他处理,种间竞争胁迫较小。本研究发现紫花苜蓿与3种禾草混播时紫花苜蓿叶片SOD,POD和CAT酶活性大于单播,说明与禾草混播能有效提高紫花苜蓿的抗氧化能力,适当的MDA能上调紫花苜蓿抗氧化能力,草地早熟禾和无芒雀麦的SOD,POD和CAT酶活性在2020年5∶5和3∶7豆禾比下高于单播,在7∶3豆禾比下低于单播,2021年各混播处理下均大于单播,说明适当比例的紫花苜蓿增大了草地早熟禾和无芒雀麦的抗氧化能力,但建植初期过大比例的紫花苜蓿对草地早熟禾和无芒雀麦的竞争胁迫造成了两种禾草抗氧化系统损伤,建植第二年逐渐适应了竞争环境。两年试验中混播苇状羊茅的SOD,POD和CAT酶活性均大于单播,说明与紫花苜蓿混播增大了无芒雀麦抗氧化能力。本研究中,3种抗氧化酶活性的变化趋势基本一致,且与MDA含量的变化基本一致,符合蒋明义[42]提出的SOD,POD和CAT三种酶协同一致维持自由基平衡的说法,与张春平[43]发现的植物所受胁迫程度影响抗氧化物质含量结果一致。
另外,植物在逆境条件下会迅速积累某些小分子化学物质如Pro,SS和SP等来维持细胞的渗透势,维持体内水分平衡和细胞膨压,从而提高混播草地牧草种间协同性。Pro是一种具有重要渗透调节作用的电解质,在逆境条件下植物会升高Pro含量来维持细胞基质与环境之间的渗透平衡和减少水分散失,其含量可从一定程度上反映植物抗逆性的强弱[44]。可溶性糖是保障植物体内物质正常运输的主要物质[45],其也参与渗透调节过程,同时还是重要的能量来源。可溶性蛋白参与植物细胞的渗透调节,可以增强植物耐脱水能力,抗性越强,植物体内可溶性蛋白越高[46]。本研究发现混播草地牧草Pro含量,SS含量和SP含量的变化趋势与MDA基本一致,与陈意兰[35]在蒭雷草(ThuareaPers)对盐胁迫的生理响应的研究中发现的蒭雷草SP和Pro含量随胁迫程度增大而增大的结果相似,与贾新平[47]发现的植物通过提高渗透调节物质含量以响应逆境胁迫的研究结果一致,说明混播植物可在逆境条件下可通过提高渗透调节物质含量来维持渗透压平衡,但建植第一年7∶3豆禾比下草地早熟禾渗透调节物质含量低于单播,说明过大的紫花苜蓿比例造成的逆境胁迫使草地早熟禾过氧化物质清除能力下降,MDA积累,造成膜脂过氧化损伤,建植第二年逐渐适应了竞争环境,为造成膜脂过氧化损伤。
在水肥相对充足的条件下,光资源是植物种间竞争的主要因素,高秆作物对矮秆作物会造成遮荫胁迫以增强自身对光资源的竞争能力[34],同时,适度的遮荫可缓解植物光抑制现象[48],增大叶绿素含量,进而提高光合利用效率[49],过度的遮荫会引起植物体内养分供应障碍,光合速率受限[50]。已有研究发现禾本科牧草生长受到氮素的的限制,而豆科牧草生长往往依赖磷素[51],本研究结果显示,混播紫花苜蓿净光合速率、蒸腾速率、气孔导度和胞间CO2浓度均高于单播,且随紫花苜蓿比例的减小和禾草比例的增大呈增大的趋势,说明混播系统中高秆、上繁草的紫花苜蓿具有光竞争优势,同时禾本科牧草活化了土壤中的磷,促进了紫花苜蓿光合作用[52],草地早熟禾在7∶3豆禾比下光合作用低于单播,可能是因为过大的紫花苜蓿比例造成了草地早熟禾遮荫胁迫,引起了养分供应障碍,使光合作用受限,其他混播处理禾草光合能力均高于单播,说明紫花苜蓿给禾草提供了适度的遮荫,同时供给附近禾草氮素,进而缓解了禾草光制止现象,使禾草上调酶活性、酶量以及叶绿素等,提高了光合作用[53]。在本试验中,豆禾混播系统中紫花苜蓿表现为高光效、高蒸腾作用,禾草均表现为低光效、低蒸腾作用,光合生理互补优势明显[54]。
本研究对不同混播处理草地牧草生理指标应用隶属函数法进行综合评价,结果显示混播紫花苜蓿生理指标综合评价D值均大于单播且在F1处理下最大,同时混播紫花苜蓿较单播增产且在F1处理下紫花苜蓿增产率最高;P1处理下草地早熟生理综合评价D值小于单播,增产率最小,其他混播处理下草地早熟禾综合评价D值大于单播,草地早熟禾增产率较P1处理增大;B2处理下无芒雀麦综合评价D值最大,增产率最大;F1处理下苇状羊茅综合评价D值最大,增产率最大。
4 结论
混播紫花苜蓿和苇状羊茅均具有较低的MDA含量和较高的抗氧化能力、渗透调节物质含量和光合能力,2020年过大的紫花苜蓿比例降低了草地早熟禾的抗氧化能力、渗透调节物质含量和光合能力以及无芒雀麦的抗氧化能力,适当减小紫花苜蓿比例能使草地早熟禾和无芒雀麦的生长生理指标好于单播。混播草地中紫花苜蓿表现为高光效、高蒸腾作用,禾草均表现为低光效、低蒸腾作用,二者光合生理协同效应明显。紫花苜蓿与与丛生型苇状羊茅7∶3混播时两种牧草生理协同性最好,紫花苜蓿和苇状羊茅均具有最高的增产率,混播草地生产力最高。
