熟地黄对6-羟基多巴胺诱导的多巴胺能神经细胞氧化应激的影响
2022-08-25王家鹏孙晓杰
王家鹏 孙晓杰
(山东中医药高等专科学校 1中医系,山东 烟台 264199;2招生就业处)
帕金森病是一种神经退行性疾病,其病理特征为脑黑质致密区域多巴胺能神经元缺失,神经元缺失可诱导纹状体多巴胺异常减少,诱导基底神经节异常,进而诱发行动迟缓、平衡功能障碍、肢体僵硬等运动症状,氧化应激被认为是多巴胺能神经细胞功能紊乱的重要原因〔1〕。目前常见的帕金森病治疗药物有多巴胺受体激动药、抗胆碱药等,这些药物多数为对症治疗,只能部分改善运动缺陷,且不能阻止病情进展,研发有效药物改善帕金森病备受关注〔2〕。熟地黄是玄参科植物地黄块根经过炮制加工而成,具有抗衰老、抗氧化、改善记忆力、调节免疫等作用〔3〕。有研究〔4〕显示,熟地黄能够缓解帕金森病模型大鼠异动症状。现阶段尚不清楚熟地黄对6-羟基多巴胺(OHDA)诱导的多巴胺能神经细胞的影响。本实验研究熟地黄对6-OHDA诱导的多巴胺能神经细胞氧化应激的影响和机制。
1 材料与方法
1.1材料 细胞:多巴胺能神经细胞SH-SY5Y(货号:258028)购自宁波明舟生物科技有限公司。多巴胺能神经细胞SH-SY5Y培养在含有10%胎牛血清的DMEM细胞培养液内,细胞培养条件为37℃,饱和湿度,5% CO2培养箱。药物:熟地黄(北京索莱宝科技有限公司,编号:DYS8270,规格:1 g)。试剂与仪器:6-OHDA(Sigma公司,美国,货号:H4381-100MG);p38MAPK激活剂Anisomycin(Sigma公司,美国,货号:A5862-0.5ML);兔抗p-p38MAPK抗体(Cell Signaling Technology公司,美国,货号:4511);活性氧(ROS)检测试剂盒(北京百奥莱博科技有限公司,货号:HR8786);兔抗剪切的含半胱氨酸的天冬氨酸蛋白水解酶(酶切Caspase)-3抗体(Abcam公司,美国,货号:ab2302);兔抗p38MAPK抗体(北京百奥莱博科技有限公司,货号:YT820);兔抗Ki-67抗体(Abgent公司,美国,货号:A-AJ1427c);丙二醛(MDA)检测试剂盒(北京索莱宝科技有限公司,货号:BC0020);DMEM培养基(Sigma公司,美国,货号:DF-042-B);兔抗Bcl-2相关X蛋白(Bax)抗体(Abcam公司,美国,货号:ab53154);超氧化物歧化酶(SOD)检测试剂盒(碧云天生物技术研究所,货号:S0101S);胎牛血清(浙江天杭生物科技股份有限公司,货号:11011-8611);乳酸脱氢酶(LDH)检测试剂盒(碧云天生物技术研究所,货号:C0017);流式细胞仪(Becton,Dickinson and Company公司,美国,LSRFortessa X-20型);酶标仪(北京普凯瑞生物科技有限公司,SpectraMax iD5型)。
1.2分组及给药方法 取处于对数生长期的多巴胺能神经细胞,根据不同的实验需求接种到细胞培养板内,24 h后,添加不同浓度的含药培养液,对照组细胞培养液不添加任何药物,6-OHDA组、熟地黄低剂量组、熟地黄中剂量组、熟地黄高剂量组、熟地黄高剂量+Anisomycin组细胞培养液中均添加100 μmol/L的6-OHDA〔5〕,同时熟地黄低、中、高剂量组细胞培养液中分别添加120、240、480 μmol/L的熟地黄,熟地黄高剂量+Anisomycin组细胞培养液中添加480 μmol/L的熟地黄和2 ng/ml的p38MAPK激活剂Anisomycin〔6〕。各组细胞培养24 h后进行相关检测。
1.3检测指标与方法 (1)CCK-8实验检测细胞增殖:多巴胺能神经细胞按照每个孔内4 000个细胞接种到96孔板内,细胞培养24 h后,分别在每个孔内添加10 μl CCK-8溶液,继续孵育10 min,用酶标仪测定波长为450 nm的吸光度(OD)值,以OD值表示细胞增殖活性。(2)Western印迹检测相关蛋白表达:多巴胺能神经细胞按照每个孔内接种5×104个细胞接种到24孔板内,24 h后,用磷酸盐缓冲液(PBS)洗涤细胞2次,添加含10%苯甲基磺酰氟(PMSF)的放射免疫沉淀(RIPA)溶液,放在冰上裂解,30 min后,在4℃条件下高速离心,收集蛋白上清,经二喹啉甲酸(BCA)方法检测蛋白浓度后,与2倍结合缓冲液混合煮沸5 min。在每个十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)上样孔内添加40 μg蛋白样品,先以90 V的电压电泳30 min后,再以120 V的电压电泳1.5 h。以200 mA恒流转膜1.5 h。将NC膜放置5%牛血清白蛋白溶液中封闭2 h。NC膜放在一抗溶液(p-p38MAPK、酶切Caspase-3抗体以1∶800稀释,p38MAPK抗体以1∶1 500稀释,Ki-67、Bax抗体以1∶1 000稀释)中,4℃摇床过夜。NC膜放在1∶2 000稀释后的二抗溶液内,在室温中结合2 h。ECL发光。Image J分析比较灰度值,以GAPDH校正。(3)流式细胞术检测凋亡:多巴胺能神经细胞按照每个孔内接种5×104个细胞接种到24孔板内,24 h后,收集细胞,用PBS洗涤细胞2次,每个测定样品取1×106个细胞,悬浮于500 μl的结合缓冲液中,分别添加5 μl的膜连蛋白(Annexin)V-FITC和碘化丙啶(PI)溶液,转移至避光条件下结合20 min,用流式细胞仪检测凋亡变化。(4)ROS、MDA、SOD含量和培养液上清中LDH检测:多巴胺能神经细胞按照每个孔内接种5×104个细胞接种到24孔板内,24 h后,收集细胞和培养液上清,以ROS检测试剂盒(CellROX Green荧光探针法)、MDA检测试剂盒(可见分光光度法)、SOD检测试剂盒(黄嘌呤氧化法)分别测定细胞中ROS、MDA、SOD含量,ROS检测结果以对照组为参照,分析其余各组ROS含量变化百分比;以LDH检测试剂盒(比色法)测定培养液上清中LDH含量,步骤均按照试剂盒说明书进行。
1.4统计学方法 采用SPSS23.0软件进行单因素方差分析。
2 结 果
2.1熟地黄对多巴胺能神经细胞增殖影响 与0 μmol/L(0.56±0.06)比较,120、240、480 μmol/L的熟地黄处理后多巴胺能神经细胞增殖活性(0.51±0.08、0.49±0.06、0.47±0.07)没有变化(P>0.05);960、1 920 μmol/L熟地黄处理后多巴胺能神经细胞增殖活性(0.40±0.06、0.32±0.04)显著降低(P<0.05)。选择对多巴胺能神经细胞没有毒性的120、240、480 μmol/L的熟地黄进行后续实验。
2.2熟地黄可抑制6-OHDA诱导的多巴胺能神经细胞中p38MAPK信号激活 与对照组(0.15±0.03)比较,6-OHDA组多巴胺能神经细胞中p-p38MAPK/p38MAPK表达量(0.89±0.09)显著升高(P<0.05);与6-OHDA组比较,熟地黄低、中、高剂量组表达量(0.63±0.05、0.41±0.03、0.29±0.02)逐渐显著降低(P<0.05);与熟地黄高剂量组比较,熟地黄高剂量+Anisomycin组表达量(0.45±0.06)显著升高(P<0.05)。见图1。
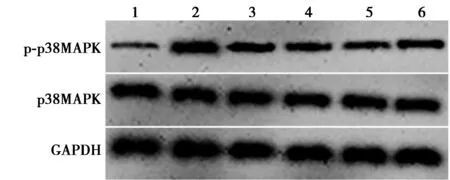
1~6:对照组、6-OHDA组、熟地黄低剂量组、熟地黄中剂量组、熟地黄高剂量组、熟地黄高剂量+Anisomycin组;同下图图1 各组多巴胺能神经细胞中p38MAPK信号 相关蛋白表达
2.3熟地黄调控p38MAPK对6-OHDA诱导的多巴胺能神经细胞增殖、凋亡影响 与对照组比较,6-OHDA组多巴胺能神经细胞增殖活性显著降低,凋亡率显著升高(P<0.05);与6-OHDA组比较,熟地黄低、中、高剂量组多巴胺能神经细胞增殖活性逐渐显著升高,凋亡率逐渐显著降低(P<0.05);与熟地黄高剂量组比较,熟地黄高剂量+Anisomycin组多巴胺能神经细胞增殖活性显著降低,凋亡率显著升高(P<0.05)。见图2,表1。
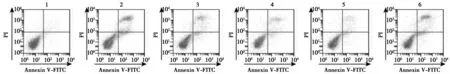
图2 各组多巴胺能神经细胞凋亡情况
表1 各组多巴胺能神经细胞增殖活性和凋亡率及Ki-67、Bax、酶切Caspase-3蛋白表达量比较
2.4熟地黄调控p38MAPK对6-OHDA诱导的多巴胺能神经细胞增殖、凋亡相关蛋白表达影响 与对照组比较,6-OHDA组多巴胺能神经细胞中Ki-67蛋白表达量显著降低,Bax、酶切Caspase-3蛋白表达量显著升高(P<0.05);与6-OHDA组比较,熟地黄低、中、高剂量组多巴胺能神经细胞中Ki-67蛋白表达量逐渐显著升高,Bax、酶切Caspase-3蛋白表达量逐渐显著降低(P<0.05);与熟地黄高剂量组比较,熟地黄高剂量+Anisomycin组多巴胺能神经细胞中Ki-67蛋白表达量显著降低,Bax、酶切Caspase-3蛋白表达量显著升高(P<0.05)。见表1,图3。
2.5熟地黄调控p38MAPK对6-OHDA诱导的多巴胺能神经细胞氧化应激影响 与对照组比较,6-OHDA组多巴胺能神经细胞中SOD含量显著降低,ROS、MDA含量显著升高,培养液上清中LDH含量升高(P<0.05);与6-OHDA组比较,熟地黄低、中、高剂量组多巴胺能神经细胞中SOD含量逐渐显著升高,ROS、MDA含量逐渐显著降低,培养液上清中LDH含量逐渐显著降低(P<0.05);与熟地黄高剂量组比较,熟地黄高剂量+Anisomycin组多巴胺能神经细胞中SOD含量显著降低,ROS、MDA含量显著升高,培养液上清中LDH含量显著升高(P<0.05)。见表2。

图3 各组各组多巴胺能神经细胞中增殖和凋亡 相关蛋白表达
表2 各组多巴胺能神经细胞中ROS、MDA、SOD含量和培养液上清中LDH含量比较
3 讨 论
帕金森病属于神经退行性疾病,多发生于中老年人,也是中老年残疾的重要诱因〔7〕。帕金森病常见病理变化为脑黑质致密带多巴胺神经细胞脱落、缺失,目前尚不清楚帕金森病发生的原因,遗传因素、兴奋性因素、自身免疫因素、氧化应激、环境因素等均与其有关,其中氧化应激因素研究最多〔8,9〕。6-OHDA是常见的体外构建帕金森病细胞模型的诱导因子〔10〕。帕金森病条件下,多巴胺能神经细胞中合成大量ROS,这些ROS不能被抗氧化酶SOD等及时清除,诱导ROS聚集,而过量的ROS可促进脂质发生过氧化,生成MDA;脂质是细胞膜的重要组成部分,其过氧化后可诱导细胞膜完整性破坏,导致原本多存在于细胞内的LDH释放至细胞外,因此检测MDA、LDH水平可间接反映氧化应激水平〔11,12〕。过量的ROS还可以激活细胞凋亡蛋白如Bax、酶切Caspase-3,促进细胞凋亡发生〔13〕。Ki-67是细胞增殖标志蛋白,其表达变化与细胞增殖水平有关〔14〕。本实验结果表明,6-OHDA诱导多巴胺神经细胞氧化应激,激活细胞凋亡,成功构建了体外帕金森病细胞模型。
熟地黄是常见中药材,具有滋阴补肾的功效,现代药理学证明熟地黄有调节免疫力、改善骨质疏松、降血糖、抗衰老、抗氧化、改善肾功能等作用〔15〕。熟地黄对神经系统有保护作用,可以抑制阿尔茨海默病大鼠海马神经细胞损伤,减少Caspase-3活化〔16〕;熟地黄治疗后的注意缺陷多动障碍大鼠神经元发育明显改善〔17〕。熟地黄对帕金森病可能有治疗功效,复方地黄处理后的帕金森病动物模型运动障碍症状减轻〔18〕;熟地黄处理后的MPP+诱导的神经细胞凋亡减少,Bax和酶切Caspase-3蛋白表达减少〔19〕。本实验结果说明熟地黄能够减轻6-OHDA诱导的多巴胺能神经细胞氧化应激,进而减少细胞凋亡,与以前的研究报道结果一致〔12〕,均提示熟地黄可能是帕金森病治疗的有效药物。
熟地黄作用机制与信号途径的转导有关〔20〕。本实验发现,熟地黄处理可降低多巴胺能神经细胞中p38MAPK磷酸化水平,熟地黄作用机制可能与p38MAPK有关。p38MAPK是MAPK信号通路的重要分支,在细胞增殖、氧化应激、细胞凋亡、细胞衰老、炎症、能量代谢过程中发挥作用〔21〕。有研究〔22〕表明,p38MAPK在帕金森病进展中过度激活,p38MAPK具有促进帕金森病发生的作用。p38MAPK在6-OHDA诱导的多巴胺能神经细胞中过度磷酸化,且抑制p38MAPK可改善多巴胺能神经细胞损伤〔23〕。本文以p38MAPK激活剂进一步验证熟地黄的作用机制,结果发现,p38MAPK激活剂能够逆转熟地黄对6-OHDA诱导的多巴胺能神经细胞增殖、凋亡和氧化应激的作用,提示熟地黄通过抑制p38MAPK信号发挥作用。
综上,熟地黄可能是帕金森病治疗的潜在药物,其能够通过抑制p38MAPK信号改善6-OHDA诱导的多巴胺能神经细胞氧化应激,进而减少细胞凋亡。关于熟地黄通过何种具体靶向机制影响p38MAPK信号进而发挥作用还有待进一步研究。
