松果菊苷对人肺癌HCC827 细胞增殖、凋亡和迁移的影响*
2022-08-24朱建勇尹义平郭雅君
李 猛,杨 力,朱建勇,尹义平,郭雅君
(湖北省十堰市人民医院·湖北医药学院附属人民医院呼吸与危重症医学中心,湖北 十堰 442000)
肺癌是最常见癌症,其中约有83%为非小细胞肺癌(NSCLC)[1]。虽然手术、化学药物治疗(简称化疗)、放射治疗(简称放疗)、免疫治疗等方案均可在一定程度上改善NSCLC 患者的预后,但对于肿瘤发生转移者,其5 年相对生存率仅约15%[2]。松果菊苷属苯乙醇苷类化合物,具有清除氧自由基、抑制炎症和调节免疫等药理活性[3]。松果菊苷能诱导乳腺癌细胞凋亡,同时抑制急性髓系白血病细胞增殖,但其对肺癌作用的研究较少[4-5]。上皮 -间充质转化(EMT)与恶性肿瘤的迁移和侵袭密切相关,虽有大量研究揭示了EMT 在NSCLC 进展中的作用,但调控EMT 的分子机制尚未明确[6]。转化生长因子-β1(TGF-β1)通过经典的Smad 信号或非典型信号传导,成为NSCLC EMT 和转移的驱动因素[7]。在TGF - β1信号通路中,TGF - β1通过结合TGF - βⅠ型受体(TβRⅠ)激活信号,进而通过磷酸化激活Smad 2和Smad 3,被激活的Smad2 和Smad3 进一步与Smad 4相互作用,激活或抑制其靶基因(如SNAIL)[8]。本研究中探讨了松果菊苷对人肺癌HCC827 细胞增殖和迁移的影响,以及TβRⅠ/ Smad 信号通路在整个过程中的作用。现报道如下。
1 材料与方法
1.1 仪器、试药与细胞
仪器:3000T 型 CO2培养箱(美国 Thermo Revco 公司);Sunrise型全自动酶标仪(瑞士Tecan公司);CKX53型倒置显微镜(日本Olympus 公司);BD FACSCanto 型流式细胞仪、Stepone 型Real - time PCR 扩增仪和Mini Protean 3型BD垂直电泳仪(美国ABI公司)。
试药:松果菊苷(苏州润新生物科技有限公司,批号为82854-37-3,纯度 98%);DMEM 培养基、胎牛血清(美国Hyclone 公司);四氮唑蓝(MTT)、二甲基亚砜(DMSO)均购自美国 Amresco 公司;AnnexinV/ PI 细胞凋亡检测试剂盒(美国BD 公司,批号为0220496);Trizol(美国Invitrogen 公司,批号为170933);基质金属蛋白酶 - 9(MMP - 9)、mRNA 反转录试剂盒和实时荧光定量聚合酶链式反应(qPCR)试剂盒(宝生物工程<大连>有限公司,批号分别为20210376,20211208);RIPA裂解液(北京索莱宝生物技术公司,批号为SL059332);TβRⅠ、Smad 2、Smad 3 和β - actin(美国 Santa Cruz 公司,批号分别为210383,210315,210564,211206);辣根过氧化物酶(HRP)标记山羊抗鼠免球蛋白(IgG,美国Abcam公司,批号为AB21054372)。
细胞:人肺癌HCC827细胞(中国科学院细胞库)。
1.2 方法
1.2.1 细胞培养与分组
在37 ℃及5% CO2条件下,用含10%胎牛血清的DMEM 培养基培养HCC827 细胞,隔天换液。实验分为阴性对照组(等体积培养基)、阳性对照组(顺铂80µmol/L)及松果菊苷低、中、高剂量组(实验Ⅰ,Ⅱ,Ⅲ组,20,40,80 µmol/L)。
1.2.2 观察指标及检测
细胞增殖能力:采用MTT 法。取对数生长期细胞,接种于96 孔板(每孔3× 103个),在37 ℃及5%CO2条件下,每孔以0(对照),5,10,20,40,80,160 µmol/L 松果菊苷或0(对照),5,10,20,40,80,160,320 µmol/ L 顺铂孵育24 h,加入MTT(每孔20 µL),继续培养4 h,弃上清液,加入DMSO(每孔200 µL)振荡10 min,以酶标仪在570 nm波长处测定吸光度值(OD值),平行测定3次,并计算细胞增殖率。细胞增殖率(%)=OD用药组/OD对照组×100%。以增殖率为50%时的药品质量浓度为半数致死浓度(LC50)。
细胞迁移能力:采用细胞划痕法。取各组对数生长期细胞,接种于6 孔板中(每孔1× 106个),以100µL 灭菌枪头在单层细胞上呈“一”字划痕,将细胞用磷酸盐缓冲液(PBS)冲洗划痕。各组以相应药液孵育24 h,从每个划痕伤口中随机选择5个视野,显微镜下观察,以图像分析仪测量划痕宽度。划痕越宽,表明细胞迁移能力越弱。
细胞凋亡情况:采用流式细胞术。取各组对数生长期细胞,接种于 6 孔板中(每孔1 × 106个),孵育细胞24 h,预冷PBS 洗涤,离心弃上清液,用Annexin V Binding Solution(1 ×)调整细胞密度为1 × 106/mL,悬浮于400µL V-FITC 溶液中,黑暗中室温孵育15 min,加入PI 10 µL,4 ℃避光孵育5 min,以流式细胞仪检测细胞凋亡情况,并计算凋亡率。
MMP-9,TβRⅠ,Smad 2,Smad 3 mRNA表达水平:采用qPCR 法。取各组对数生长期细胞,接种于6孔板中(每孔1×106个),孵育细胞24 h,用预冷PBS洗涤,离心弃上清液,用Trizol提取总RNA,反转录制备cDNA。PCR反应条件为,95 ℃预变性10 s;94 ℃变性15 s,40个循环;55 ℃退火30 s;70 ℃延伸30 s 终止反应。以 2-△△Ct法计算MMP-9,TβRⅠ,Smad 2,Smad 3 mRNA 的相对表达量。各组设3个复孔,实验重复3次。引物序列见表1。

表1 引物序列Tab.1 Primer′s sequence
TβRⅠ,Smad 2,Smad 3 蛋白表达水平:采用Western blot法。取各组对数生长期细胞,接种于6孔板中(每孔1× 106个),孵育细胞24 h,用预冷PBS 洗涤,离心弃上清,加入RIPA 裂解液裂解2 h,离心取上清,电泳,转到聚偏二氟乙烯膜上,然后用5%脱脂牛奶在室温下封闭 1 h,将膜与 TβRⅠ(1∶400)、Smad 2(1∶200)、Smad 3(1∶300)和 β - actin(1∶1 000)抗体孵育,4 ℃过夜,将HRP 标记山羊抗鼠 IgG(1∶10 000)在室温下孵育30 min,显色,采集图像,利用Image J 软件分析目的条带与内参条带灰度比值,即为各目的蛋白相对表达量。
1.3 统计学处理
2 结果
2.1 细胞增殖能力和LC50
随着松果菊苷浓度的增加(10~160 µmol / L)与0 µmol/L 比较,细胞增殖率显著降低(P< 0.05)。当松果菊苷浓度为40µmol/L 时,增殖率约为50%,LC50为(47.50±6.21)µmol/L,故选择40µmol/L为松果菊苷干预浓度。随着顺铂浓度的增加(10~320 µmol/ L)与0 µmol/L 比较,细胞增殖率显著降低(P< 0.05),当顺铂浓度为 80 µmol/ L 时,增殖率约为 50%,LC50为(76.85 ± 14.89)µmol/L,故选择80 µmol/L 为顺铂干预浓度。结果见表2。
表2 松果菊苷及顺铂对HCC827细胞增殖的影响(,%,n=3)Tab.2 Effects of echinacoside and cisplatin on the proliferation of HCC827 cells(,%,n=3)
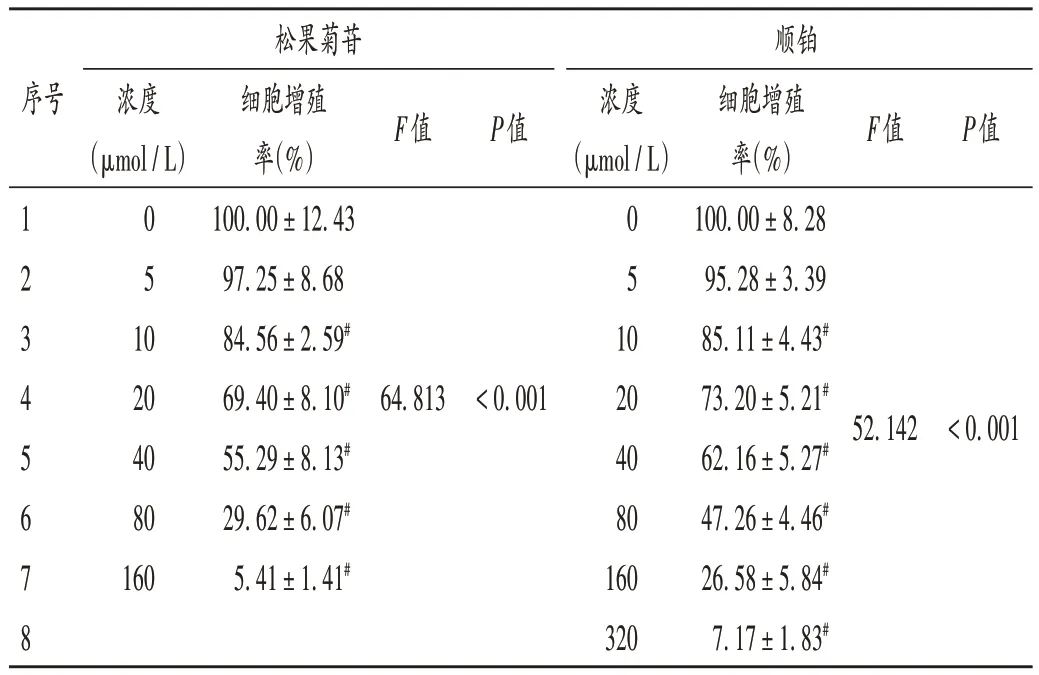
表2 松果菊苷及顺铂对HCC827细胞增殖的影响(,%,n=3)Tab.2 Effects of echinacoside and cisplatin on the proliferation of HCC827 cells(,%,n=3)
注:与0 µmol/L比较,#P < 0.05。Note:Compared with those at 0 µmol/L,#P < 0.05.
序号松果菊苷浓度(µmol/L)F值P值顺铂浓度(µmol/L)F值P值1 2 3 4 5 6 7 8 0 5 1 0 0 5 1 0 20 40 80 160细胞增殖率(%)100.00±12.43 97.25±8.68 84.56±2.59#69.40±8.10#55.29±8.13#29.62±6.07#5.41±1.41#64.813<0.00120 40 80 160 320细胞增殖率(%)100.00±8.28 95.28±3.39 85.11±4.43#73.20±5.21#62.16±5.27#47.26±4.46#26.58±5.84#7.17±1.83#52.142<0.001
2.2 细胞迁移和凋亡情况
与阴性对照组比较,阳性对照组及实验Ⅰ,Ⅱ,Ⅲ组细胞迁移距离缩短,细胞凋亡率显著升高(P<0.05)。详见表3和图1、图2。

图1 HCC827细胞迁移情况Fig.1 The migration of HCC827 cells

图2 HCC827细胞凋亡情况Fig.2 The apoptosis of HCC827 cells
2.3 细胞中mRNA 及蛋白表达水平
与阴性对照组比较,阳性对照组及实验Ⅰ,Ⅱ,Ⅲ组细胞中TβRⅠ,Smad 2,Smad 3 mRNA 及蛋白表达水平均显著降低(P< 0.05)。详见表3。
表3 各组HCC827细胞迁移、凋亡情况及细胞中mRNA和蛋白表达水平比较(,n=3)Tab.3 Comparison of migration,apoptosis and the mRNA and protein expression levels of HCC827 cells in each group (,n=3)
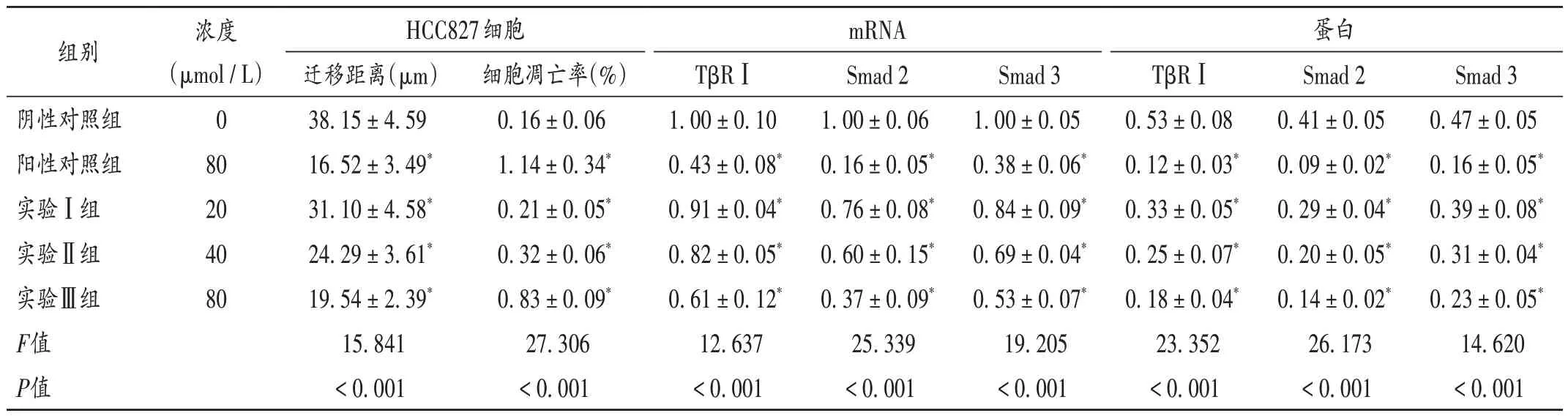
表3 各组HCC827细胞迁移、凋亡情况及细胞中mRNA和蛋白表达水平比较(,n=3)Tab.3 Comparison of migration,apoptosis and the mRNA and protein expression levels of HCC827 cells in each group (,n=3)
注:与阴性对照组比较,*P <0.05。Note:Compared with those in the negative control group,*P < 0.05.
组别mRNA 蛋白浓度(µmol/L)0 80 20 40 80 HCC827细胞迁移距离(µm)38.15±4.59 16.52±3.49*31.10±4.58*24.29±3.61*19.54±2.39*15.841<0.001 Smad 3 0.47±0.05 0.16±0.05*0.39±0.08*0.31±0.04*0.23±0.05*14.620<0.001细胞凋亡率(%)0.16±0.06 1.14±0.34*0.21±0.05*0.32±0.06*0.83±0.09*27.306<0.001阴性对照组阳性对照组实验Ⅰ组实验Ⅱ组实验Ⅲ组F值P值TβRⅠ1.00±0.10 0.43±0.08*0.91±0.04*0.82±0.05*0.61±0.12*12.637<0.001 Smad 2 1.00±0.06 0.16±0.05*0.76±0.08*0.60±0.15*0.37±0.09*25.339<0.001 Smad 3 1.00±0.05 0.38±0.06*0.84±0.09*0.69±0.04*0.53±0.07*19.205<0.001 TβRⅠ0.53±0.08 0.12±0.03*0.33±0.05*0.25±0.07*0.18±0.04*23.352<0.001 Smad 2 0.41±0.05 0.09±0.02*0.29±0.04*0.20±0.05*0.14±0.02*26.173<0.001
3 讨论
松果菊苷可通过诱导肝癌细胞凋亡抑制肝癌进展,对胰腺癌、乳腺癌等肿瘤细胞也有抑制作用[9]。研究发现,松果菊苷通过抑制EMT,进而抑制急性髓系白血病细胞体外迁移[4],但松果菊苷对HCC827 细胞的抗转移作用尚未见报道。本研究中,松果菊苷以剂量依赖的方式抑制HCC827 细胞的迁移和增殖,并诱导其凋亡;同时,松果菊苷可抑制TβRⅠ/Smad 信号通路,作用虽无顺铂强,但潜在副作用较低。
EMT 在肿瘤细胞转移早期起着至关重要的作用,因为细胞失去黏附,获得了更强的迁移和侵袭能力,向远处组织扩散。EMT 的调控涉及多种信号通路,其中TGF- β 信号通路较典型[10]。TGF- β 主要由白细胞介素4(IL-4)或白细胞介素13(IL-13)刺激免疫细胞分泌,也可由TGF - β 激活的Ⅱ型肺泡上皮细胞、成纤维细胞、细胞外基质释放[11]。TβRⅠ与TβR Ⅱ结合形成一种能识别并结合配体的复合物(TGF - β)。TGF - β 可上调TβRⅠmRNA 和蛋白表达,诱导细胞内TβRⅠ肽磷酸化,激活细胞内信号转导,包括Smad 通路和非Smad 通路的信号转导[12]。在 TβRⅠ / Smad 通路中,激活TβRⅠ使Smad 2/3 磷酸化水平升高,并与Smad 4 结合形成 Smad 2/3-4 复合物[13]。Smad 7 属 Smad 通路拮抗剂,它可抑制Smad 2/3 的磷酸化和Smad 2/3-4 复合物的形成,并且抑制TGF - β 下调[14]。Smad 2/ 3 - 4复合物随后易位进入细胞核,作为转录因子,导致纤维化转录因子上调[15]。靶向 TβRⅠ的抑制剂,如 LY -2157299、EW7197、SD - 208 和 SB431542,成功地抑制了临床前模型和Ⅰ~Ⅲ期临床试验中的肿瘤进展[16]。本研究结果显示,松果菊苷可降低HCC827细胞中TβRⅠmRNA 和蛋白表达水平,并抑制Smad 2 和Smad 3 mRNA 和蛋白表达,表明松果菊苷具有抑制HCC827 细胞增殖和迁移的作用。
此外,TβRⅠ也可直接激活非Smad信号通路,包括丝裂原活化蛋白激酶/ c - Jun 氨基末端激酶(MAPK/JNK)、MAPK/p38和磷脂酰肌醇3激酶(PI3K)/蛋白激酶B(Akt)信号通路,可直接或间接调控细胞凋亡[17]。在MAPK/ JNK 和MAPK/ p38 信号通路中,体外结果显示,经 SP - 600125 处理 24 h 后也降低了 JNK 和 p38 的磷酸化水平,诱导肿瘤细胞凋亡[18]。同时,SD - 208 对MAPK/ JNK 和MAPK/ p38 信号通路的抑制作用被SB203580 处理逆转[19]。本研究结果显示,松果菊苷可诱导HCC827 细胞凋亡,这可能与TβRⅠ可直接激活非Smad 信号通路有关,但本研究中未对TβRⅠ非smad 信号通路进行研究,后续将持续完善。
综上所述,松果菊苷可抑制HCC827 细胞增殖及迁移,并促进细胞凋亡,机制可能与其抑制TβRⅠ/Smad信号通路激活有关。
