光叶丁公藤及其3个潜在代用品治疗类风湿性关节炎差异性成分和作用机制研究
2022-08-16杨媛媛崔小敏陈志永
胡 静,杨媛媛,任 慧,崔小敏,李 宁,曲 彤,陈志永*
光叶丁公藤及其3个潜在代用品治疗类风湿性关节炎差异性成分和作用机制研究
胡 静1,杨媛媛2,任 慧1,崔小敏1,李 宁1,曲 彤1,陈志永1*
1. 陕西省中医药研究院,陕西 西安 710061 2. 西安市食品药品检验所,陕西 西安 710054
目的 研究光叶丁公藤与3个潜在代用品多花丁公藤、凹脉丁公藤、大果飞蛾藤治疗类风湿性关节炎(rheumatoid arthritis,RA)的差异性成分和潜在作用机制。采用超高效液相色谱-四极杆/静电场轨道阱高分辨质谱法(UHPLC-Q Exactive Focus MS/MS)分析光叶丁公藤及3个潜在代用品的差异成分;结合Swiss Target Prediction、DisGeNET、OMIM、TTD等数据库获得差异成分治疗RA的作用靶点;采用STRING数据库构建共有靶点互作(protein-protein interaction,PPI)网络模型,采用Cytoscape 3.6.0软件进行可视化处理并分析网络拓扑参数筛选出核心靶点;通过DAVID数据库和R语言软件对靶点进行基因本体(gene ontology,GO)分析和京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)信号通路富集分析,采用Cytoscape 3.6.0软件构建“药材-差异成分-靶点-通路(H-C-T-P)”网络图并进行分析,探讨光叶丁公藤及3个潜在代用品差异成分治疗RA的潜在作用机制。从光叶丁公藤及3个潜在代用品中共鉴定60个成分,其中差异成分35个,共涉及348个靶点。与RA相关的差异成分有18个,得到差异成分-RA共同靶点153个。GO和KEGG富集分析结果显示光叶丁公藤及3个潜在代用品可能通过芒柄花素、丁公藤黄素E、圣草酚、-反式-对香豆酰酪胺、秦皮乙素共5个活性差异成分作用于RAC-α丝氨酸/苏氨酸蛋白激酶(RAC-alpha serine/threonine-protein kinase,AKT1)、双特异性丝裂原活化蛋白激酶激酶1(dual specificity mitogen-activated protein kinase kinase 1,MAP2K1)、磷脂酰肌醇4,5-二磷酸3-激酶催化亚单位α亚型(phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit alpha isoform,PIK3CA)等32个靶点,调节PI3K-Akt信号通路、Rap1信号通路、Ras信号通路等5个信号通路发挥治疗RA的作用。初步明确了光叶丁公藤及3个潜在代用品治疗RA的差异成分、关键靶点和核心通路;光叶丁公藤和大果飞蛾藤所鉴定的共有化学成分数量和H-C-T-P网络图中度值较其他2个潜在代用品高,说明大果飞蛾藤更适合做光叶丁公藤代用品;为濒危中药代用品开发提供了相对可靠的新策略。
丁公藤;差异性成分;网络药理学;类风湿性关节炎;多花丁公藤;凹脉丁公藤;大果飞蛾藤;芒柄花素;丁公藤黄素E;圣草酚;-反式-对香豆酰酪胺;秦皮乙素
类风湿性关节炎(rheumatoid arthritis,RA)是一种慢性、以炎性滑膜炎为主的全身性自身免疫性疾病。其以手、足小关节的多关节、对称性、侵袭性关节炎症为特征,常伴有关节外器官受累及血清类风湿因子阳性,可以导致关节畸形及功能丧失[1]。Yamamoto等[2]研究发现,机体的免疫网络失衡,成纤维样滑膜细胞的类肿瘤样增殖和细胞内大量分泌金属蛋白酶和促炎性细胞因子等,是造成RA关节内炎性细胞聚集、滑膜组织增生、骨及软骨降解和破坏的主要因素。目前临床上常规治疗RA主要为西药,如非甾体抗炎药(nonsteroidal antiinflammatory drugs,NSAIDs)、慢作用抗风湿药物(slow-acting antirheumatic drugs,SAARDs)、糖皮质激素、生物制剂等[3]。但是,化学药所带来的一系列不良反应[4]使得具有辨证施治、系统调节及不良反应小等特点[5]的中医药治疗越来越受到人们的重视。
丁公藤药材最早见于《本草拾遗》,其在《中国药典》2020年版收载品种为旋花科丁公藤属植物丁公藤Benth. 或光叶丁公藤Craib的干燥藤茎,具有祛风胜湿、消肿止痛的功效,主治风湿痹痛、风湿性关节炎、跌打损伤等症[6-7]。近年来,由于生态环境恶化以及临床需求量的快速增长,丁公藤和光叶丁公藤资源已经近于枯竭,无法满足人们的用药需求,大果飞蛾藤Henmsl. 现已成为丁公藤的主流替代品种及事实上《中国药典》所载丁公藤的代用品[8-9]。另据文献报道,旋花科植物多花丁公藤Merr.、凹脉丁公藤Merr. et Chun.与丁公藤、光叶丁公藤成分近似,提示具有作为丁公藤药材新基原/代用品的潜力[10]。本课题组在前期研究中发现光叶丁公藤和大果飞蛾藤、多花丁公藤、凹脉丁公藤等3个潜在代用品主成分近似,并建立了4种药材中8个共同成分的HPLC含量测定方法[11-12]。但光叶丁公藤与3个潜在替代品的化学成分差异,及差异性成分对治疗RA的影响尚未得到系统阐释。
本研究构建了有公开发表文献支撑的旋花科丁公藤属和飞蛾藤属植物化学成分数据库,在该数据库的基础上采用超高效液相色谱-四极杆/静电场轨道阱高分辨质谱(UHPLC-Q Exactive Focus MS/MS)联用技术对光叶丁公藤及3个潜在代用品的化学成分进行快速鉴定和确证,得到光叶丁公藤及3个潜在代用品的化学成分并分析差异成分;采用网络药理学的方法预测与筛选差异性成分治疗RA的作用靶点,系统阐释成分差异可能造成的机制差异。本研究为丁公藤药材潜在代用品开发提供了实验支持,研究技术与方法为濒危中药代用品开发提供了示范。
1 仪器与材料
1.1 仪器
UPLC-Q Exactive Focus液质联用系统:Thermo Q Exactive型质谱仪(美国赛默飞世尔科技公司),Ultimate3000超高效液相色谱系统(美国戴安公司),Xcalibur 4.0工作站(美国赛默飞世尔科技公司),CD(Compound discovery,2.1)化合物分析鉴定软件(美国赛默飞世尔科技公司);色谱柱:Thermo Accucore aQ RP18(美国赛默飞世尔科技公司);KQ-100型超声波清洗机(昆山市超声仪器有限公司);BS210S型万分之一、BT25S型十万分之一电子分析天平[赛多利斯科学仪器(北京)有限公司]。
1.2 药材与试剂
对照品东莨菪内酯、绿原酸、新绿原酸、伞形花内酯、秦皮乙素和咖啡酸(上海圻明生物科技有限公司,批号分别为161208、1701904、17062003、18010202、18092803、17122804,质量分数均≥98%),东莨菪苷、隐绿原酸、异绿原酸A、异绿原酸B、异绿原酸C、(四川维克奇生物科技有限公司,批号分别为wkq20021510、wkq20082705、wkq20020403、wkq20021003、wkq20031101,质量分数均≥98%)。色谱纯甲醇(Thermo Fisher Scientific,美国)、色谱纯甲酸(Sigma-Aldrich,美国),均为LC-MS级,水为超纯水(屈臣氏蒸馏水),其他试剂均为分析纯。
光叶丁公藤(采自广东省阳西县,批号20111020;购自缅甸克钦邦,批号20121207;采自海南,批号20101102)、多花丁公藤(采自海南,批号20110918)、凹脉丁公藤(采自海南,批号20111023;采自海南,批号20131121)、大果飞蛾藤(购自南宁药材市场,批号20130717;佛山冯了性药业有限公司提供,批号20110829;购自广西,批号20130120;佛山冯了性药业有限公司提供,批号20130916;购自海南,批号20120825)清洗、切片、干燥并保存备用,经陕西省中医药研究院陈志永副研究员分别鉴定为旋花科植物光叶丁公藤Craib、多花丁公藤Merr.、凹脉丁公藤Merr. et Chun.和大果飞蛾藤Henmsl.的干燥藤茎。
2 方法
2.1 光叶丁公藤及3个潜在代用品的成分分析
2.1.1 样品的制备
(1)供试品溶液制备[13]:精密称取光叶丁公藤及3个潜在代用品的样品粉末(过40目筛)0.5 g,置100 mL具塞锥形瓶中。精密加入80%甲醇水50 mL,称定质量。常温下超声处理30 min(40 kHz,400 W),放冷,用80%甲醇水补足损失的质量。摇匀,滤过,取续滤液5 mL转移至10 mL量瓶中,40%甲醇水定容至刻度,经0.22 μm微孔滤膜滤过,即得供试品溶液。
(2)对照品溶液制备[13]:取各对照品约1.0 mg,精密称定并置于10 mL量瓶中,加入甲醇溶解并稀释至刻度,摇匀制备成0.1 mg/mL的单一对照品储备液;分别量取上述储备液适量,混合后用纯甲醇稀释,最终制备成各对照品质量浓度均为1 μg/mL的混合对照品溶液。以上对照品溶液均置于4 ℃冰箱保存,备用。
2.1.2 UPLC-Q-Orbitrap分析条件 色谱条件[13]:Thermo Accucore aQ RP18色谱柱(150 mm×2.1 mm, 2.6 μm);柱温为30 ℃;进样量为3 μL;体积流量0.3 mL/min;二元梯度洗脱:流动相为甲醇(A)- 0.1%的甲酸水溶液(B),梯度洗脱:0~12 min,5%~25% A;12~20 min,25%~30% A;20~28 min,30%~38% A;28~40 min,38%~42% A。
质谱条件[13]:离子源为HESI,正负离子检测模式;鞘气流量为45 arb,辅助气流量为15 arb,喷雾电压为3500 V;毛细管温度为350 ℃;雾化温度为320 ℃;扫描模式为Full MS/dd-MS,Full MS分辨率为70 000,dd-MS分辨率为17 500,扫描范围为/100~1200。MS/MS模式时所用碰撞能梯度为20、40 eV。
2.1.3 LC/MS数据分析 数据采集在Xcalibur 4.0工作站进行。检索国内外文献自建丁公藤属、飞蛾藤属植物化学成分数据库,根据化合物精确相对分子质量、二级碎片信息(差异设定<5×10−6),通过在线检索、自建成分库检索及对照品比对,进行化学成分结构鉴定。
2.2 网络药理学研究
2.2.1 光叶丁公藤及3个潜在代用品的差异成分整理 对5批大果飞蛾藤、3批光叶丁公藤、2批凹脉丁公藤、1批多花丁公藤进行了成分鉴定,成分表征过程中成分的选取均尽量综合多批次样品质谱分析结果。将以上批次光叶丁公藤及3个潜在代用品鉴定的化学成分进行归类,找出共有成分和差异成分,采用Cytoscape 3.6.0软件展示,成分度值(degree)<4为差异成分。
2.2.2 关键靶点的富集 利用PubChem数据库(https://pubchem.ncbi.nlm.nih.gov/search/)获取差异成分的2D结构并下载其sdf文件保存备用,将sdf文件导入Swiss Target Prediction(http://www. swisstargetprediction.ch/)数据库中预测差异成分的靶点,去掉重复靶点后即得差异成分潜在靶点。在TTD(http://db.idrblab.net/ttd/)、Drugbank(https:// www.Drugbank.ca/)和DisGeNET(https://www.d- isgenet.org/)数据库查询RA相关靶点,去掉重复值即得RA相关靶点。取每一个药材的差异成分作用靶点与疾病相关靶点交集,获得差异成分治疗风湿性关节炎的潜在关键靶点。
2.2.3 蛋白-蛋白互作(protein-protein interaction,PPI)网络构建 将“2.2.2”项差异成分和RA相映射得到的共同靶点在STRING平台进行PPI网络分析,并将基本设置(Basic Settings)中的网络边缘设置为置信度(confidence);互作资源(active interaction sources)设置为文本挖掘(Textmining)、实验(Experiments)、数据库(Databases)、共表达(Co-expression)、邻近(Neighborhood)、基因融合(Gene Fusion)和共存(Co-occurrence);所需的最低互作分数(minimum required interaction score)设置为最高置信度(highest confidence)0.900进行PPI分析。应用Cytoscape 3.6.0软件绘制共同靶点互作网络图,并进行分析。
2.2.4 基因本体(gene ontology,GO)分析和京都基因与基因组百科全书(Kyoto encyclopedia of genes and genomes,KEGG)通路富集分析 为了探索光叶丁公藤及3个潜在代用品差异成分治疗RA潜在靶点的相关信号通路,将差异成分治疗RA的靶点导入DAVID数据库(https://david.ncifcrf. gov/home.jsp),限定物种为人,设定阈值<0.05,进行GO分析和KEGG通路富集分析,筛选出光叶丁公藤及3个潜在代用品的差异成分治疗RA的重要生物进程和信号通路。
2.2.5 “药材-差异成分-靶点-通路”网络构建与分析 由“2.2.4”项筛选出KEGG通路富集分析中基因富集数量前20的通路,通过查阅文献,再筛选出可能与治疗RA相关的通路,找出富集在这些通路上的差异成分治疗RA的潜在靶点,并找出对应的差异成分及归属药材。将药材-差异成分、差异成分-靶点、靶点-通路数据表导入Cytoscape 3.6.0软件,运用“Merge”功能得到3个数据表的交集,即药材- 差异成分-靶点-通路网络图。
3 结果
3.1 差异成分的获取
UPLC/Q Exactive Focus MS/MS分析得到光叶丁公藤及3个潜在代用品的总离子流图,见图1。从光叶丁公藤及3个潜在代用品中共预测了60个成分,其中有11个化合物通过与对照品比对明确识别。光叶丁公藤、凹脉丁公藤、多花丁公藤和大果飞蛾藤分别鉴定49、37、39和47个成分,各化合物信息见表1。图2为光叶丁公藤及3个代用品各成分含量示意图,颜色越深代表相应成分质谱峰面积越大。在样品称样量相差小于0.005 0 g条件下,相应成分质谱峰面积越大代表该成分在该药材中的含量越高。其中化合物6、8、12、15、24、35、42、51颜色较深、峰面积较大,分别为新绿原酸、东莨菪苷、绿原酸、隐绿原酸、东莨菪内酯、异绿原酸B、异绿原酸A、异绿原酸C,与文献报道的丁公藤属植物主成分一致[11-12]。共有成分和差异成分用Cytoscape 3.6.0软件展示出来,共有成分25个,差异成分35个,见图3。
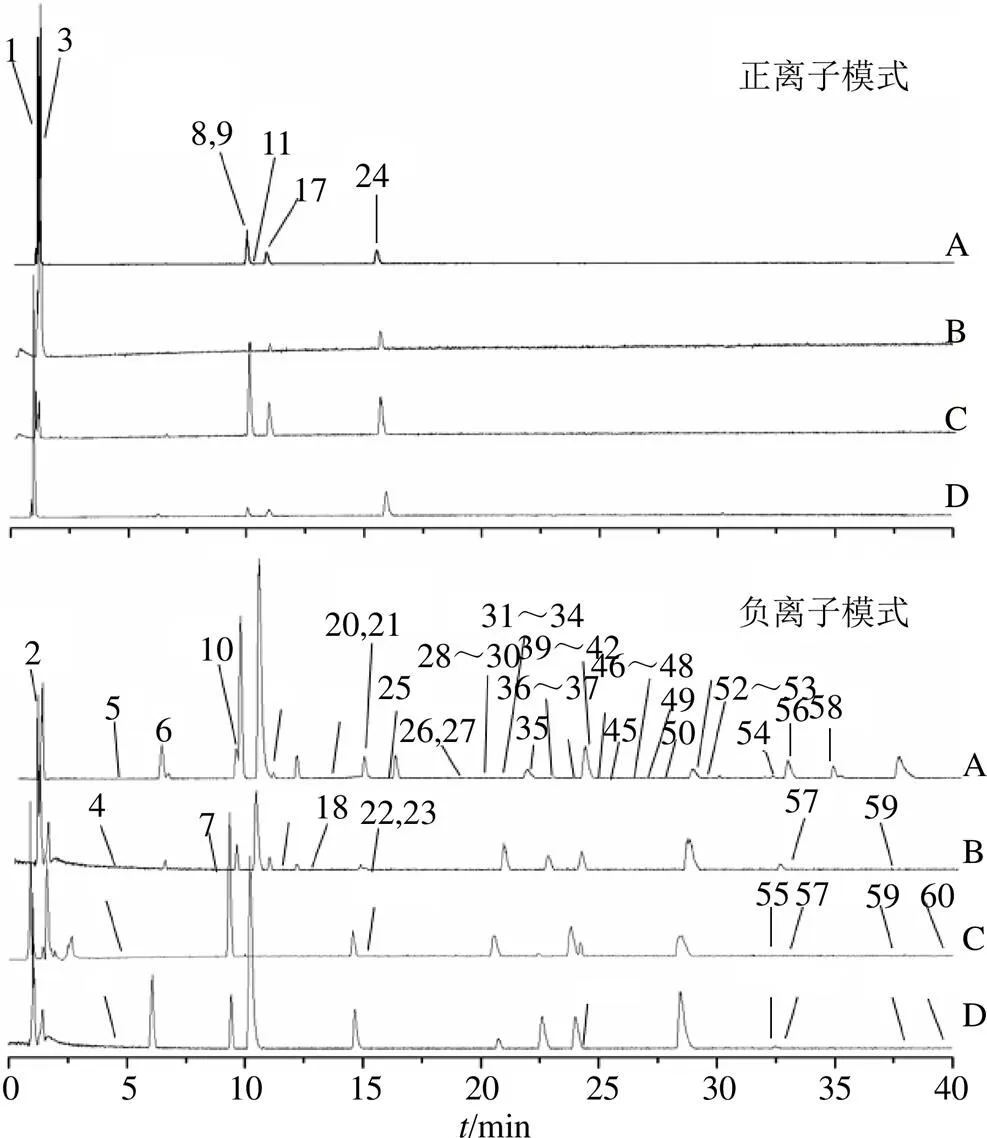
图1 光叶丁公藤(A)、多花丁公藤(B)、凹脉丁公藤(C)、大果飞蛾藤(D)在正离子和负离子模式下的基峰色谱图
3.2 差异成分靶点获取及与RA靶点映射分析
基于“3.1”项下所识别的差异成分,本研究采用Swiss Target Prediction数据库对35个差异成分进行反向预测靶点,共得到715个靶点,去掉未预测到靶点的化合物及重复靶点后,共得到光叶丁公藤及3个潜在代用品的差异成分18个,潜在靶点348个。通过TTD、GeneCards数据库检索“rheumatoid arthritis”,绘图软件将差异成分反向找靶所得到的348个靶点映射到RA相关靶点,即得光叶丁公藤及3个潜在代用品的差异成分抗RA的潜在作用靶点共153个,见图4。
表1 光叶丁公藤及3个潜在代用品化学成分的鉴定
Table 1 Identification of chemical compounds in Erycibe schmidtii and three substitutes
序号tR/min实测值(m/z)理论值(m/z)模式误差(×10−6)分子式碎片离子(m/z)化合物名称归属 10.97144.101 5144.101 9[M+H]+−2.77C7H13NO2126.091 0, 108.080 6包公藤丙素或凹脉丁公藤碱ABCD 21.00267.065 6267.065 2[M-H]−1.50C16H12O4191.054 4, 113.023 8芒柄花素ABC 31.38186.112 8186.112 5[M+H]+1.61C9H15NO3144.101 1, 108.080 1包公藤甲素ACD 44.05315.123 5315.122 7[M-H]−2.54C18H20O5152.010 4, 108.020 9丁公藤黄素EBCD 54.52153.017 8153.018 2[M-H]−−3.27C7H6O4141.911 0, 123.007 43,4-二羟基苯甲酸ABCD 6*6.07353.086 3353.086 8[M-H]−−1.42C16H18O9191.054 8, 179.033 7,173.044 0, 135.044 2新绿原酸ABCD 78.82 529.134 1[M-H]−4.85C26H26O12191.059 2, 179.035 7, 173.044 2, 135.045 23,5-二咖啡酰奎宁酸甲酯或3,4-二咖啡酰奎宁酸甲酯或4,5-二咖啡酰奎宁酸甲酯B 8*9.42 355.102 4[M+H]+−2.82C16H18O9193.049 3, 178.025 9,165.054 3, 133.028 2东莨菪苷ABCD 99.43193.049 7193.049 5[M+H]+−1.55C10H8O4178.025 8, 133.028 6东莨菪内酯同分异构体ABCD 109.51 211.060 1[M-H]−4.26C10H12O5196.038 2, 181.050 4, 163.039 6, 153.055 63-(2,4,5-三羟基苯基)丙酸甲酯ABCD 1110.22 355.102 4[M+H]+−2.82C16H18O9163.038 9, 145.028 5, 135.044 4东莨菪苷同分异构体ABCD 12*10.23353.086 3353.086 8[M-H]−−1.42C16H18O9191.056 0, 173.043 9, 161.022 3, 135.044 0绿原酸ABCD 13*10.24179.033 4179.033 9[M-H]−−2.79C9H8O4135.043 5咖啡酸ABCD 14*10.71177.018 2177.019 3[M-H]−3.87C9H6O4133.028 3, 104.025 3秦皮乙素ABD 15*11.78353.086 3353.086 8[M-H]−−1.42C16H18O9191.054 8, 173.045 2, 161.023 2隐绿原酸ABCD 1611.84529.1366529.134 1[M-H]−4.73C26H26O12367.102 1, 191.056 8, 173.044 53,5-二咖啡酰奎宁酸甲酯或3,4-二咖啡酰奎宁酸甲酯或4,5-二咖啡酰奎宁酸甲酯B 17*11.86163.038 6163.038 8[M+H]+−1.23C9H6O3145.028 2, 135.043 8, 117.033 6伞形花内酯ABCD 1812.47529.1366529.134 1[M-H]−4.73C26H26O12374.498 6, 191.059 5, 173.044 13,5-二咖啡酰奎宁酸甲酯或3,4-二咖啡酰奎宁酸甲酯或4,5-二咖啡酰奎宁酸甲酯B 1913.22337.106 2337.107 1[M-H]−−2.67C20H18O5191.054 8, 173.034 4, 119.049 4刺桐素BACD 2014.57367.101 7367.102 4[M-H]−−1.91C17H20O9191.054 5, 173.043 8, 135.035 6绿原酸甲酯ACD 2114.64665.173 0665.171 2[M-H]−2.71C30H34O17197.045 4, 176.011 3, 121.029 3丁公藤苷A或丁公藤苷B或丁公藤苷CAD 2215.38581.220 0581.222 9[M-H]−−4.99C28H38O13419.167 4, 373.126 7, 233.079 6, 153.054 5南烛木树脂酚-3α-O-β-D-吡喃葡萄糖苷或其同分异构体BC 2315.89581.220 0581.222 9[M-H]−−4.99C28H38O13419.168 8, 373.125 3, 233.079 7, 153.054 8南烛木树脂酚-3α-O-β-D-吡喃葡萄糖苷或其同分异构体BC 24*16.29193.049 7193.049 5[M+H]+−1.55C10H8O4178.025 5, 149.059 1, 137.059 5, 133.028 1东莨菪内酯ABCD 2516.93695.180 4695.181 8[M-H]−−2.01C31H36O18359.096 6, 197.044 1, 173.043 7, 153.053 8丁公藤苷FABCD 2618.89665.173 0665.171 2[M-H]−2.71C30H34O17197.045 5, 191.034 8, 153.055 6, 121.029 3丁公藤苷A或丁公藤苷B或丁公藤苷CAD 2719.08635.159 7635.160 7[M-H]−−1.57C29H32O16191.033 4, 167.033 4, 121.043 3丁公藤苷D或丁公藤苷EAD 2820.61627.189 1627.192 0[M-H]−−4.62C28H36O16447.112 1, 429.102 0, 315.072 3, 197.045 7丁公藤苷N或猫尾木酚苷B或二者同分异构体A 2920.74533.129 5533.129 0[M-H]−0.94C25H26O13371.097 6, 353.086 7, 335.076 8, 197.045 3, 173.045 2, 161.024 1,135.045 05-O-咖啡酰基-4-O-丁香酰奎宁酸或5-O-咖啡酰基-3-O-丁香酰奎宁酸或4-O-咖啡酰基-5-O-丁香酰奎宁酸或5-O-咖啡酰基-4-O-香草酰奎宁酸甲酯ABCD 3020.83635.159 7635.160 7[M-H]−−1.57C29H32O16327.314 5, 191.034 9, 167.034 9, 123.045 0丁公藤苷D或丁公藤苷EA
续表1
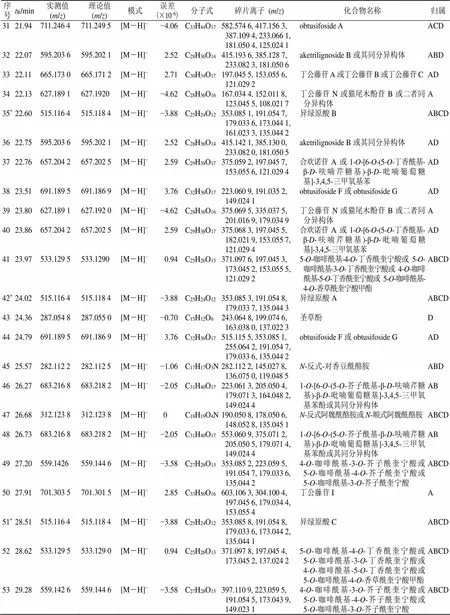
序号tR/min实测值(m/z)理论值(m/z)模式误差(×10−6)分子式碎片离子(m/z)化合物名称归属 3121.94711.246 4711.249 5[M-H]−−4.06C33H44O17582.574 6, 417.156 3, 387.109 4, 233.066 1, 181.050 4, 125.024 1obtusifoside AACD 3222.07595.203 6595.202 1[M-H]−2.52C28H36O14415.193 6, 385.128 7, 233.082 3, 181.050 6aketrilignoside B或其同分异构体ABD 3322.11665.173 0665.171 2[M-H]−2.71C30H34O17197.045 5, 153.055 6, 121.029 2丁公藤苷A或丁公藤苷B或丁公藤苷CAD 3422.13627.189 1627.1920[M-H]−−4.62C28H36O16167.034 4, 152.011 8, 123.045 5, 108.021 7丁公藤苷N或猫尾木酚苷B或二者同分异构体A 35*22.60515.116 4515.118 4[M-H]−−3.88C25H24O12353.085 1, 191.054 7, 179.033 6, 173.044 1, 161.023 3, 135.044 2异绿原酸BABCD 3622.75595.203 6595.202 1[M-H]−2.52C28H36O14415.142 1, 385.130 0, 233.082 0, 181.050 5aketrilignoside B或其同分异构体AD 3722.76657.204 2657.202 5[M-H]−2.59C29H38O17375.059 2, 197.045 7, 153.055 6, 121.029 4合欢诺苷A或1-O-[6-O-(5-O-丁香酰基- β-D-呋喃芹糖基)-β-D-吡喃葡萄糖基]-3,4,5-三甲氧基苯AD 3823.51691.189 5691.186 9[M-H]−3.76C32H36O17223.060 9, 191.035 2, 149.024 1obtusifoside F或obtusifoside GAD 3923.80627.189 1627.192 0[M-H]−−4.62C28H36O16375.069 5, 335.037 5, 201.016 9, 179.034 9丁公藤苷N或猫尾木酚苷B或二者同分异构体A 4023.86657.204 2657.202 5[M-H]−2.59C29H38O17375.068 3, 197.045 5, 182.021 9, 153.055 7, 121.029 4合欢诺苷A或1-O-[6-O-(5-O-丁香酰基- β-D-呋喃芹糖基)-β-D-吡喃葡萄糖基]-3,4,5-三甲氧基苯AD 4123.97533.129 5533.1290[M-H]−0.94C25H26O13371.097 6, 197.045 3, 173.045 2, 153.055 5, 121.029 25-O-咖啡酰基-4-O-丁香酰奎宁酸或5-O-咖啡酰基-3-O-丁香酰奎宁酸或4-O-咖啡酰基-5-O-丁香酰奎宁酸或5-O-咖啡酰基- 4-O-香草酰奎宁酸甲酯ABCD 42*24.02515.116 4515.118 4[M-H]−−3.88C25H24O12353.085 3, 191.054 8, 179.033 7, 135.044 3异绿原酸AABCD 4324.36287.054 8287.055 0[M-H]−−0.70C15H12O6243.064 8, 199.074 6, 163.038 0, 137.022 3圣草酚D 4424.79691.189 5691.186 9[M-H]−3.76C32H36O17515.115 5, 353.085 1, 255.064 2, 191.054 7, 179.033 6, 135.044 2obtusifoside F或obtusifoside GAD 4525.57282.112 2282.112 5[M-H]−−1.06C17H17O3N282.112 2, 145.027 8, 136.075 0, 119.048 5N-反式-对香豆酰酪胺ABD 4626.27683.216 8683.218 2[M-H]−−2.05C31H40O17223.061 3, 205.050 4, 179.071 3, 164.048 2, 149.024 41-O-[6-O-(5-O-芥子酰基-β-D-呋喃芹糖基)-β-D-吡喃葡萄糖基]-3,4,5-三甲氧基苯酚或其同分异构体AB 4726.68312.123 8312.123 8[M-H]−0C18H19O4N190.050 8, 178.050 6, 148.052 8, 135.045 1N-反式阿魏酰酪胺或N-顺式阿魏酰酪胺ABCD 4826.73683.216 8683.218 2[M-H]−−2.05C31H40O17553.060 9, 375.071 2, 205.050 5, 179.071 4, 149.024 41-O-[6-O-(5-O-芥子酰基-β-D-呋喃芹糖基)-β-D-吡喃葡萄糖基]-3,4,5-三甲氧基苯酚或其同分异构体AB 4927.20559.1426559.144 6[M-H]−−3.58C27H28O13353.085 2, 223.059 5, 191.054 7, 179.033 6, 135.044 24-O-咖啡酰基-3-O-芥子酰奎宁酸或5-O-咖啡酰基-4-O-芥子酰奎宁酸或5-O-咖啡酰基-3-O-芥子酰奎宁酸ABCD 5027.91701.303 5701.301 5[M-H]−2.85C33H50O16603.106 3, 304.100 4, 197.045 6, 179.034 4, 153.055 4丁公藤苷IA 51*28.51515.116 4515.118 4[M-H]−−3.88C25H24O12353.085 8, 191.054 8, 179.033 6, 173.044 2, 135.044 1异绿原酸CABCD 5228.62533.129 5533.129 0[M-H]−0.94C25H26O13371.097 8, 197.045 4, 173.045 2, 137.024 25-O-咖啡酰基-4-O-丁香酰奎宁酸或5-O-咖啡酰基-3-O-丁香酰奎宁酸或4-O-咖啡酰基-5-O-丁香酰奎宁酸或5-O-咖啡酰基-4-O-香草酰奎宁酸甲酯ABCD 5329.28559.142 6559.144 6[M-H]−−3.58C27H28O13397.110 9, 223.059 5, 191.054 5, 173.043 9, 149.023 14-O-咖啡酰基-3-O-芥子酰奎宁酸或5-O-咖啡酰基-4-O-芥子酰奎宁酸或5-O-咖啡酰基-3-O-芥子酰奎宁酸ABCD
续表1
*通过对照品对比确认;药材归属:A-光叶丁公藤 B-多花丁公藤 C-凹脉丁公藤 D-大果飞蛾藤
*Confirmed by comparison with the reference substances; Herb attribution: A-B-C-D-
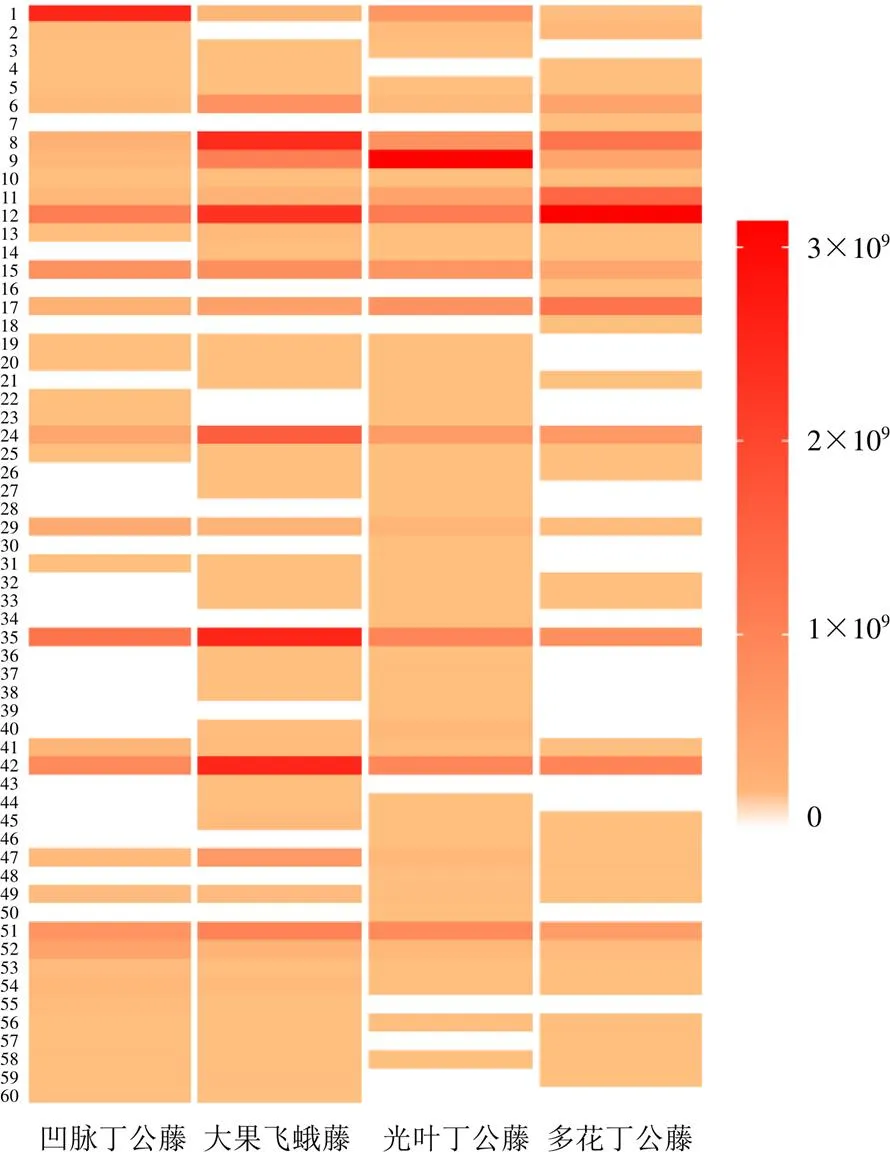
颜色由黄至红代表含量的增大,白色代表该成分在对应药材中未被检出即峰面积为0
3.3 差异成分治疗RA潜在靶点PPI网络构建及潜在靶点拓扑参数分析
将“3.2”项差异成分和RA相映射得到的共同靶点在STRING平台进行PPI网络分析。为对PPI的网络拓扑参数进一步分析,将其导入Cytoscape 3.6.0中重新构建,见图5。其中,碳酸杆酶1(carbonic anhydrase 1,CA1)、钾电压门控通道亚家族A成员3(potassium voltage-gated channel subfamily A member 3,KCNA3)、血清对氧磷酶/芳基酯酶1(serum paraoxonase/arylesterase 1,PON1)等34个靶标节点未与其他靶标节点构成相互作用,因此Cytoscape 3.6.0构建的PPI网络中共显示119个节点,521条边。应用NetworkAnalyzer对其进行分析,利用各节点的度值设置节点大小和颜色,其节点的度值由小变大,则节点形状由小变大,颜色由黄变红,节点越大、颜色越红表示度值越大,说明在网络中越关键;“combined score”设置边的粗细,边越粗,表示“combined score”越大,说明靶点之间的连接关系越紧密。

三角形节点表示差异成分,菱形节点表示共有成分
图4 差异成分抗RA靶点图
在Cytoscape 3.6.0软件中对候选靶点PPI网络的拓扑学参数进行分析,选取度值、接近中心性、中介中心性均大于均值的靶点作为关键靶点(表2)。候选靶点PPI网络分析结果显示,119个潜在靶标节点的度值均值为8.6,接近中心性均值为0.396 9,中介中心性均值为0.014 0,共有23个靶点的3个网络拓扑学参数同时大于相应的均值。这23个靶点包括脾酪氨酸激酶(tyrosine-protein kinase,SYK)、细胞周期蛋白依赖性激酶(cyclin-dependent kinase 1,CDK1;Cyclin-dependent kinase 5,CDK5)、RAC-α丝氨酸/苏氨酸蛋白激酶(RAC-alpha serine/threonine-protein kinase,AKT1)、磷脂酰肌醇4,5-二磷酸3-激酶催化亚单位α亚型(phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit alpha isoform,PIK3CA)、AP-1转录因子(transcriptionfactor AP-1,JUN)、血管内皮生长因子A(vascular endothelial growth factor A,VEGFA)、表皮生长因子受体(epidermal growth factor receptor,EGFR)、基质金属蛋白酶(72 kDa ype IV collagenase,MMP2;matrix metalloproteinase-9,MMP9)、炎症因子白细胞介素-4(interleukin-4,IL-4)等,不仅在网络关系中有重要作用,在差异成分治疗RA的过程中也起到了重要作用,可作为研究光叶丁公藤与3个潜在代用品差异成分治疗RA的关键靶点基因。
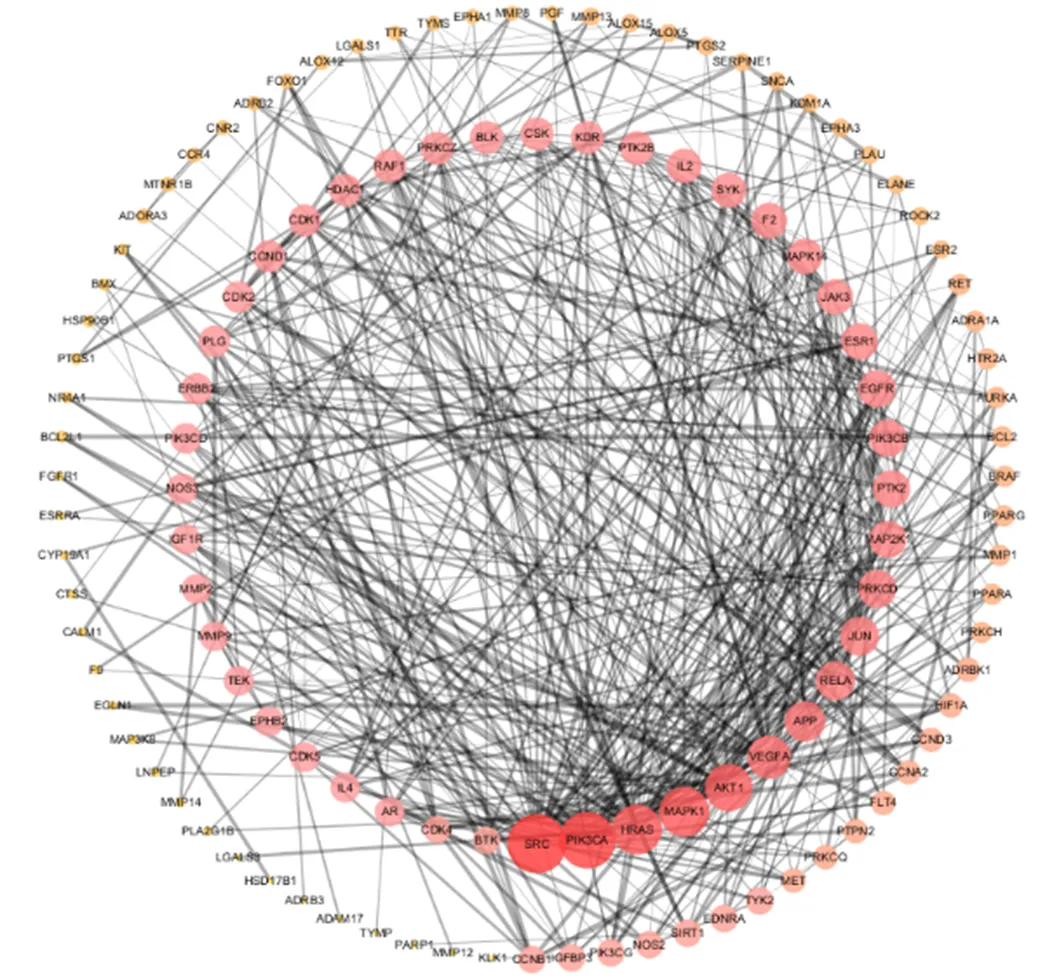
图5 差异成分治疗RA的PPI
表2 差异成分治疗RA的候选靶点拓扑学参数
Table 2 Topology parameters of candidate targets for discriminatory components in treatment of RA
靶点中文全称ID度值接近中心性中介中心性 SRC原癌基因酪氨酸蛋白激酶P12931440.567 30.174 3 PIK3CA磷脂酰肌醇4,5-二磷酸3-激酶催化亚基α亚型P42336410.546 30.119 8 MAPK1丝裂原活化蛋白激酶1P28482330.526 80.106 5 HRAS肉瘤病毒癌基因P01112330.500 00.063 3 AKT1RAC-α丝氨酸/苏氨酸蛋白激酶P31749300.508 60.104 7 VEGFA血管内皮生长因子AP15692250.513 00.098 7 APPβ-淀粉样蛋白前体蛋白P05067220.485 60.129 0 JUNAP-1转录因子P05412200.473 90.037 2 RELA转录因子p65Q04206200.493 70.043 5 PRKCD蛋白激酶Δ型Q05655190.457 40.021 7 EGFR表皮生长因子受体P00533180.468 30.032 2 MAP2K1双特异性丝裂原活化蛋白激酶激酶1Q02750180.466 40.025 2 ESR1雌激素受体P03372170.470 10.062 1 F2凝血酶原P00734160.460 90.042 6 MAPK14丝裂原活化蛋白激酶14Q16539160.450 40.015 7 CCND1G1/S-特异性细胞周蛋白-D1P24385120.412 60.017 4 CDK1细胞周期蛋白依激酶1P06493120.414 00.043 8 ERBB2酪氨酸激酶受体2P04626110.435 40.021 5 PLG纤溶酶原P00747110.400 00.036 4 IL4白细胞介素-4P0511290.402 70.039 4 CDK5细胞周期蛋白依激酶5Q0053590.411 10.020 8 MMP9基质金属蛋白酶9P1478090.408 30.031 4 MMP2基质金属蛋白酶2P0825390.411 10.029 8
3.4 GO生物进程分析和KEGG通路富集分析
为了更系统地深入研究光叶丁公藤及3个潜在代用品差异成分对RA的作用机制,将差异成分治疗RA的153个靶蛋白通过DAVID数据库进行GO分析,共得到GO条目284个,其中生物过程(biological process,BP)201个,分子功能(molecular function,MF)47个,细胞组分(cellular component,CC)36个,BP、MF、CC分别占比70.77%、16.55%、12.68%。以<0.01为筛选条件,选取值最小的排名前10的生物进程,根据富集基因数的大小进行排序并作图,见图6。结果发现,这些蛋白在BP方面主要涉及血管生成(angiogenesis)、固有免疫应答(innate immune response)、细胞增殖的调节(regulation of cell proliferation)、ERK1和ERK2级联的正调节(positive regulation of ERK1 and ERK2 cascade)等;在MF方面主要涉及ATP结合(ATP binding)、蛋白质丝氨酸/苏氨酸激酶活性(protein serine/threonine kinase activity)、丝氨酸型内肽酶活性(serine-type endopeptidase activity)、受体结合(receptor binding)、非跨膜蛋白酪氨酸激酶活性(non-membrane spanning protein tyrosine kinase activity)等;在CC方面主要涉及细胞外外切体(extracellular exosome)、细胞外间隙(extracellular space)、细胞液(cytosol)、细胞外基质(extracellular matrix)、质膜细胞质侧的外源成分(extrinsic component of cytoplasmic side of plasma membrane)等。
利用DAVID数据库平台对差异成分治疗RA的靶蛋白进行KEGG富集分析,共得到了102条通路。除去诸如前列腺癌、大肠癌和胰腺癌等具体疾病或其他无关条目后,以<0.01为筛选条件,选取其中显著性最高的20条通路用R语言软件进行富集分析。见图7,气泡图的横坐标(gene radio)表示每条通路中涉及的核心靶点占该通路总靶点数的比率;气泡大小表示该通路中涉及到的核心靶点数;气泡颜色由绿至红表示值越来越小,即该通路的显著性越高。
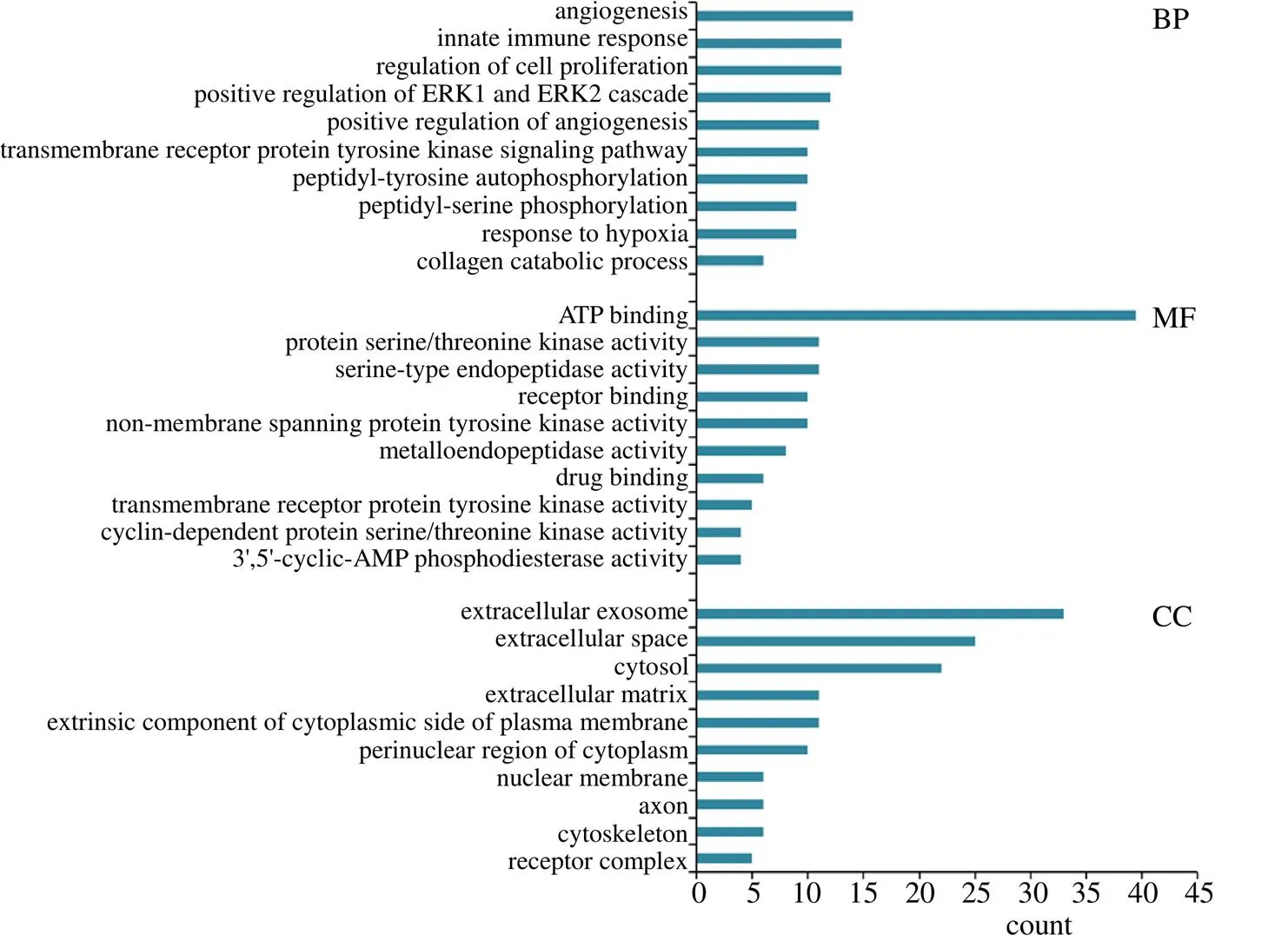
图6 GO功能富集分析

图7 KEGG功能富集分析(前20)
3.5 光叶丁公藤及3个潜在代用品“药材-差异成分-靶点-通路(H-C-T-P)”网络构建及分析
综合GO分析和KEGG通路富集分析的结果,分析筛选出可能具有治疗RA作用的通路,并与差异成分和潜在靶点一一对应,即通路与潜在靶点对应,潜在靶点与差异活性成分对应,差异活性成分与药材对应。将相关数据导入Cytoscape 3.6.0软件,构建光叶丁公藤及3个潜在代用品“药材-差异成分- 靶点-通路”网络图,共有111个节点(20条通路、70个靶点、17个差异成分、4个药材)和455条边。采用Network Analyzer对网络的拓扑参数进行分析,并用Degree Sorted Circle Layout对节点进行布局见图8,其节点的度值越大,节点越大,说明在网络中越关键。
应用NetworkAnalyzer对H-C-T-P网络拓扑参数进行分析,结果如下:70个潜在靶标节点的度值均值为4.1、接近中心性均值为0.349 7、中介中心性均值为0.003 5,共有32个靶点的3个网络拓扑学参数同时大于相应的均值,分别为AKT1、丝裂原活化蛋白激酶(dual specificity mitogen-activated protein kinase kinase 1,MAP2K1;mitogen-activated protein kinase 1,MAPK1;mitogen-activated protein kinase 14,MAPK14)、PIK3CA、PIK3CD、PIK3CG、EGFR、转录因子p65(transcription factor p65,RELA)、JUN、MMP2、胰岛素样生长因子1受体(insulin-like growth factor 1 receptor,IGF1R)、VEGFA、G1/S特异性细胞周期蛋白D1(G1/S-specific cyclin-D1,CCND1)、血管内皮生长因子受体2(vascular endothelial growth factor receptor 2,KDR)、SYK、腺苷受体(adenosine receptor A2a,ADORA2A;adenosine receptor A3,ADORA3)、内皮型一氧化氮合酶(nitric oxide synthase, endothelial,NOS3)、细胞周期蛋白依赖激酶2(cyclin-dependent kinase 2,CDK2)、CDK4、原癌基因酪氨酸蛋白激酶(proto-oncogene tyrosine-protein kinase Src,SRC)、纤溶酶原激活物抑制剂1(plasminogen activator inhibitor 1,SERPINE1)、胎盘生长因子(placenta growth factor,PGF)、MMP9、雌激素受体(estrogen receptor,ESR1; Estrogen receptor beta,ESR2)、钙调素-1(calmodulin-1,CALM1)、凋亡调节因子Bcl-2(apoptosis regulator Bcl-2,BCL2)、IL2、IL4、Bcl-2样蛋白1(Bcl-2-like protein 1,BCL2L1);17个差异活性成分的度值均值为10.6、接近中心性均值为0.372 8、中介中心性均值为0.030 7,共有5个差异成分的3个网络拓扑学参数同时大于相应的均值,分别为化合物2、43、45、14、19;20条通路的度值均值为13.6、接近中心性均值为0.382 5、中介中心性均值为0.026 0,共有5个信号通路的3个网络拓扑学参数同时大于相应的均值,分别为磷酸肌醇3激酶-蛋白激酶B信号通路(PI3K-Akt signaling pathway)、小G蛋白Rap信号通路(Rap1 signaling pathway)、Ras信号通路(Ras signaling pathway)、缺氧诱导因子信号通路(HIF-1 signaling pathway)、雌激素信号通路(Estrogen signaling pathway)。以上分析结果显示,光叶丁公藤及3个潜在代用品的差异成分靶点分布于不同的通路并协同发挥治疗RA作用,同时关键靶点也分布在不同通路上表明富集的RA通路可作为光叶丁公藤及3个潜在代用品差异成分治疗RA的重要通路。
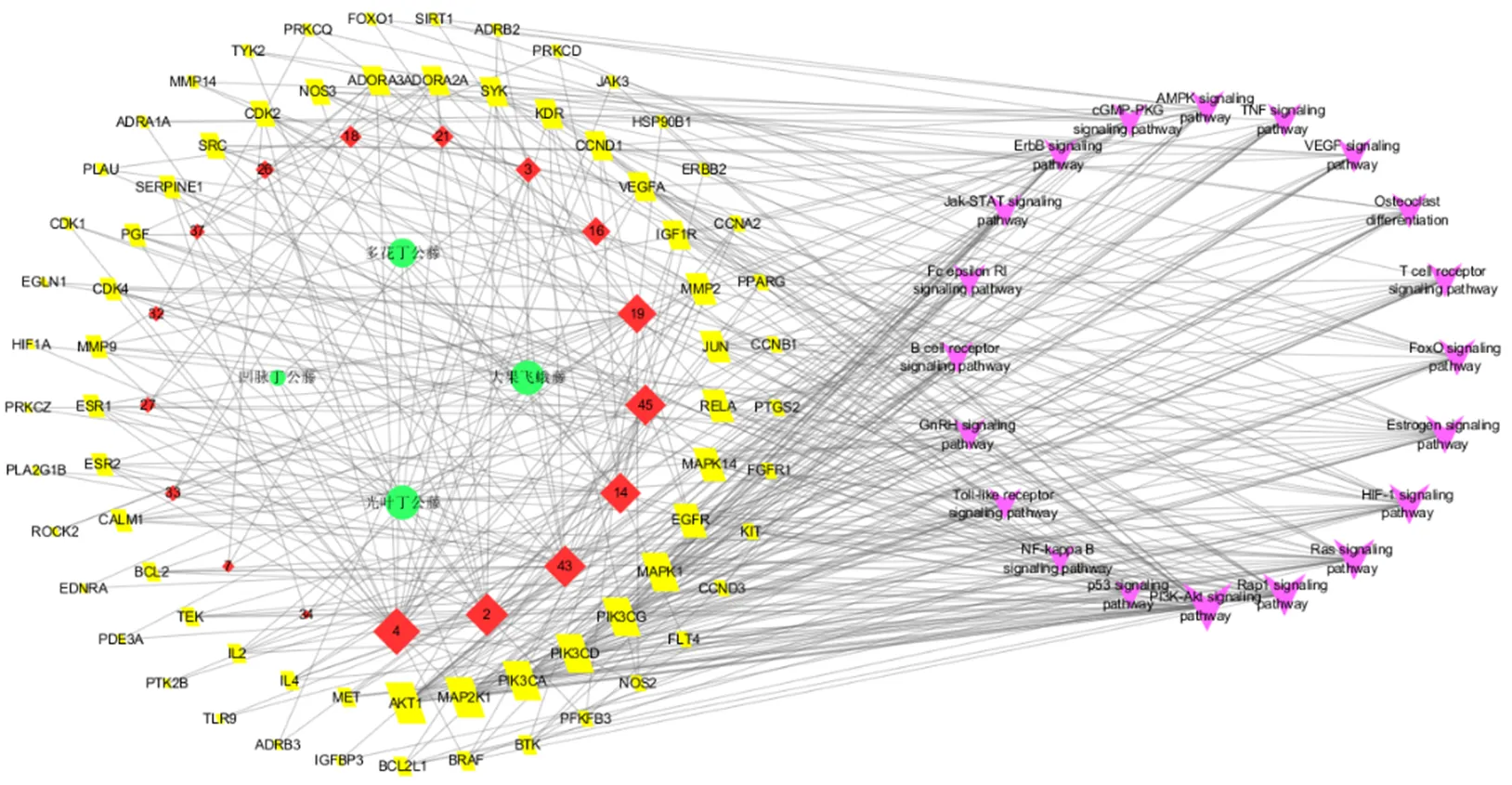
圆形节点表示药材,菱形节点表示差异成分,平行四边形节点表示靶点,“V”形表示信号通路
另外,4个药材所连接的差异活性成分数量分别为光叶丁公藤12个、大果飞蛾藤12个、凹脉丁公藤4个、多花丁公藤8个,可见光叶丁公藤和大果飞蛾藤的化学成分在数量和活性上更为相似。
4 讨论
4.1 差异性成分分析
本研究以光叶丁公藤及3个潜在代用品为研究对象,基于UHPLC-Q Exactive Focus MS/MS平台,共鉴定了60个化学成分,其中差异成分35个,为分析其潜在差异药效成分奠定了物质基础。在35个差异成分中,化合物3为生物碱类,化合物2、4、19、43为黄酮类,化合物7、16、18、20、55、57、59、60为奎尼酸衍生物;化合物14、21、26、27、30、33为香豆素类,化合物22、23、31、32、36、38、44为木脂素类,化合物45为酰胺类;化合物28、34、37、39、40、46、48、50为其他类。结合表1和图2可知,化合物43为大果飞蛾藤专属成分,化合物7、16、18为多花丁公藤专属成分,化合物28、30、39、50为光叶丁公藤专属成分,凹脉丁公藤无专属化合物。专属成分的鉴别将为冯了性风湿跌打药酒、藤络宁胶囊、丁公藤注射液等中成药中丁公藤药材投料品种的快速鉴别提供依据。另外,在35个差异性成分中,光叶丁公藤与多花丁公藤、凹脉丁公藤、大果飞蛾藤分别有6、5、16个共同化合物。因此,理论上可认为大果飞蛾藤与光叶丁公藤在化学成分上更加近似,具有更大的成为丁公藤药材代用品的潜力。
4.2 差异性成分网络药理学研究
通过Cytoscape 3.6.0软件对H-C-T-P网络图进行网络分析发现5个化合物的3个拓扑参数均大于均值。这5个化合物主要存在于光叶丁公藤和大果飞蛾藤中,具有抗炎镇痛、软骨保护、健骨等药理作用。芒柄花素(2)可抑制骨组织MMP-9的合成和分泌、破骨细胞的骨吸收,同时降低肿瘤坏死因子(tumor necrosis factor,TNF)及IL6水平,具有明显的健骨作用[14];圣草酚(43)可通过抑制核因子-κB(nuclear factor-kappa B,NF-κB)信号通路,激活核因子E2相关因子2(nuclear factor E2 related factor 2,Nrf2)/ 血红素氧合酶-1(heme oxygenase-1,HO-1)通路,发挥抑制IL-1β刺激的软骨细胞炎症效应的作用[15];-反式-对香豆酰酪胺(45)和刺桐素B(19)均可抑制脂多糖诱导的RAW 264.7细胞释放一氧化氮,发挥抗炎作用[16];秦皮乙素(14)在小鼠耳肿胀和醋酸扭体实验中表现出显著的抗炎活性和外周镇痛作用[17],能够通过抑制MMP的合成、分泌及其活性,治疗骨关节炎和风湿性关节炎造成的软骨损伤[18]。
将光叶丁公藤及3个潜在代用品差异成分靶点映射到RA疾病靶点并构建网络,获得153个共同靶点。进一步PPI网络分析显示SRC、PIK3CA、MAPK1等23个靶点最为重要,提示其为光叶丁公藤及3个潜在代用品差异成分治疗RA的关键靶点。这些靶点主要包含丝氨酸/苏氨酸/酪氨酸蛋白激酶、丝裂原活化蛋白激酶、细胞周期蛋白依赖性激酶、磷脂酰肌醇-3激酶、炎症因子等。KEGG通路富集分析整合预测结果显示,RA发病机制与关键靶点及其所分布的通路有密切关联,这与光叶丁公藤及3个潜在代用品差异成分及其关键靶点的研究结果和其与RA间关系的相关研究基本一致。
SYK是蛋白酪氨酸激酶(protein-tyrosine kinase,PTK)中的一种,是一种非受体型的酪氨酸激酶[19]。SYK在B细胞受体Fc受体等免疫信号的传导中起重要作用[20]。而B细胞等免疫细胞在RA发病中发挥了关键作用,有研究应用口服型SYK抑制剂fostamatinib(R788)作用于实验小鼠后,可观察到其能够控制类风湿关节炎实验小鼠的滑膜炎、关节炎以及骨侵蚀等,证实滑膜组织中的炎症细胞和SYK水平有关[21]。AKT1是3种密切相关的丝氨酸/苏氨酸蛋白激酶(AKT1、AKT2和AKT3)之一,被称为Akt激酶,它调控代谢、增殖、细胞存活、生长和血管生成等许多过程,如股骨头坏死[22]和维持骨量及骨转化[23-24]等。AKT1还可以促进软骨内骨血管生成和骨化[25],这对修复RA所造成的关节损伤有重要作用。PI3K是肌醇与磷脂酰肌醇(phosphatidylinositol,PI)的重要激酶,PIK3CA、PIK3CD、PIK3CG均为PI3K Ι型催化亚单位。PI3K参与多种信号通路,与Akt共同组成的PI3K/Akt信号通路在RA患者的滑膜组织中PI3K表达水平升高,升高的PI3K促进Akt磷酸化,激活PI3K/Akt信号通路;活化的Akt可磷酸化多种蛋白,介导细胞的生长发育和血管生成的调节[26],甚至引起PIK3CA相关的过度生长现象[27]。已有动物实验证明JUN的表达和骨细胞、成骨细胞的凋亡有关,与原发性骨质疏松关系密切[28],故JUN可能在RA病程发展及转移的过程中起到重要作用。MAPK1、MAPK14等属于丝裂原活化蛋白激酶家族,已证实MAPK1能辅助β-雌二醇通过细胞外调节蛋白激酶(extracellular regulated protein kinases,ERK-1/2)抑制RA滑膜细胞的凋亡和MMF3的释放,是治疗RA的潜在靶点[29];雌激素不仅对性发育和生殖功能重要,而且参与RA的病程发展,阻止健康人向RA发展[30-31]。ESR1为介导骨内雌激素作用的受体,能够促进成骨细胞增殖分化、抑制破骨细胞,但是由于雌激素可能同时具有抗炎和促炎特性,所以ESR1是否会加剧骨关节炎的发生仍有争议[32]。VEGFA是血管生成和成骨的有效耦合所必需的,具有促进骨形成减少骨吸收的作用[33]。RA滑膜衬里层VEGF蛋白表达与血管形态、弯曲血管走行方式及血管密度均呈正相关[34]。EGFR能通过刺激关节软骨细胞的增殖发挥合成代谢的作用[35];CCND1表达抑制会抑制软骨细胞增殖[36];CDKs是一类调控细胞周期及细胞转录的丝氨酸/苏氨酸蛋白激酶,而RA患者化膜细胞异常增殖可能与CDK1、CDK5的过度表达有关[37]。MMPS是一类锌离子依赖的内肽酶,是胞外基质降解过程中主要的蛋白酶,参与关节炎及转移、细胞外基质的破裂等正常生理过程。研究表明绝经后女性血清MMP-2及MMP-9水平随骨密度的降低呈现升高趋势,可成为新的反映骨代谢的指标[38]。另外,IL-4等是临床常见的与炎症相关的靶点,如IL-4可以抑制炎症反应[39]。
KEGG富集的20条通路,经H-C-T-P网络拓扑参数分析表明PI3K-Akt、Rap1、Ras、HIF-1、Estrogen信号通路共5条通路的度值、接近中心性、中介中心性均大于均值。其中PI3K-Akt信号通路与RA滑膜细胞的增殖或凋亡有关[40]。Ras是小分子G蛋白,Rap1为Ras相关蛋白属于Ras癌基因家族成员之一。Ras信号通路可以增强血管内皮生长因子VEGF的表达[41],表明Ras信号通路可能在RA血管生成中起重要作用;有研究表明Rap1的过度表达会增强T细胞的激活和增加IL-2的产生,可能在RA作用机制中起到免疫调节作用[42];另外,在血小板整合素激活及血小板生成过程中Rap1信号通路也发挥重要作用[43]。由于RA患者滑膜细胞大量增殖,其关节腔为缺氧环境,缺氧诱导因子(hypoxia-inducible factor 1,HIF-1)表达增加。HIF-1信号通路可以调节VEGF信号通路,通过上调VEGF而激活NOS,从而促进血管生成与舒张[44]。Estrogen信号通路与骨骼的生长与重构有关[45]。这5条包括了153个共同靶点中的49个,占32%。故推测光叶丁公藤及3个代用品的差异成分可能是通过与RA相关生物通路中的靶蛋白相作用来维持滑膜内环境稳态,降低炎症反应、防止和减少关节的破坏,从而改善患者生活质量。
中国医学科学院药用植物研究所根据“亲缘关系相近的植物类群具有相似的化学成分”的亲缘学理论,寻找出一批药效非常相似的替代品,如国产萝芙木、新疆阿魏等[46-47]。本研究在亲缘学理论指导下开展丁公藤药材的代用品开发,将代用品开发目标定位于丁公藤、光叶丁公藤的近缘品种,研究结果也切实验证了亲缘学理论。综上,珍稀濒危植物光叶丁公藤及3个代用品的差异成分中芒柄花素、圣草酚、-反式-对香豆酰酪胺、刺桐素B及秦皮乙素可能作用于SRC、PIK3CA、MAPK1等23个RA相关靶点,调控包括PI3K-Akt、Rap1、Ras、HIF-1和Estrogen在内的多条信号通路。光叶丁公藤及3个代用品的差异成分通过多靶点、多通路调节免疫、调控炎症因子、抑制滑膜细胞增殖、抑制软骨破坏及促进成骨细胞增殖分化等达到治疗RA的目的。本研究系统阐释了光叶丁公藤及其3个潜在代用品的成分异同,探讨了成分差异在治疗RA方面可能导致的机制差异。本研究为丁公藤药材代用品开发,中成药中正品丁公藤与潜在代用品投料的快速鉴别提供了依据。同时,本研究采用的技术与方法为珍稀濒危中药代用品开发研究提供了新的思路。
利益冲突 所有作者均声明不存在利益冲突
[1] 谢曜联, 汪悦. 艾条灸法治疗类风湿关节炎初探 [J]. 陕西中医, 2014, 35(8): 1054-1055.
[2] Yamamoto A, Fukuda A, Seto H,. Suppression of arthritic bone destruction by adenovirus-mediated dominant-negative Ras gene transfer to synoviocytes and osteoclasts [J]., 2003, 48(9): 2682-2692.
[3] 池里群, 周彬, 高文远, 等. 治疗类风湿性关节炎常用药物的研究进展 [J]. 中国中药杂志, 2014, 39(15): 2851-2858.
[4] 莫永生. 抗类风湿性关节炎药物不良反应及临床研究进展 [J]. 临床合理用药杂志, 2016, 9(9): 172-173.
[5] 世界中医药学会联合会, 中华中医药学会. 国际中医临床实践指南类风湿关节炎 (2019-10-11) [J]. 世界中医药, 2020, 15(20): 3160-3168.
[6] 郑婧, 张贵君. 丁公藤研究进展 [A] //第四届中国中药商品学术大会暨中药鉴定学科教学改革与教材建设研讨会论文集 [C]. 北京: 中国商品学会, 2015: 243-246.
[7] 谭建宁, 高振霞. 丁公藤的研究进展 [J]. 广西科学院学报, 2008, 24(1): 49-52.
[8] 吴立宏, 朱恩圆, 张紫佳, 等. 广西产丁公藤原植物的调查及商品丁公藤主流品种的鉴定 [J]. 中草药, 2005, 36(9): 1398-1400.
[9] 中国科学院中国植物志委员会, 中国植物志. 第64卷[M]. 北京: 科学出版社, 1989: 5-6.
[10] 徐小昆, 陈志永, 廖立平, 等. 丁公藤属植物中东莨菪苷、绿原酸、东莨菪素、异绿原酸A、异绿原酸B和异绿原酸C的含量测定 [J]. 中国中药杂志, 2015, 40(6): 1119-1122.
[11] 杨媛媛, 谢志民, 胡静, 等. 3种丁公藤属植物中8个成分的含量测定 [J]. 中南药学, 2020, 18(6): 982-985.
[12] 胡静, 崔小敏, 杨媛媛, 等. HPLC法同时测定大果飞蛾藤中8种成分 [J]. 中成药, 2020, 42(3): 666-670.
[13] 胡静, 杨媛媛, 任慧, 等. 光叶丁公藤中化学成分的UPLC-Q-Exactive Focus-MS/MS鉴定 [J]. 中国实验方剂学杂志, 2020, 26(18): 124-132.
[14] 喻琴云, 詹晓峰, 黄彦. 刺芒柄花素对去势骨质疏松大鼠P38MAPK/MMP-9信号通路的影响 [J]. 中国临床药理学与治疗学, 2017, 22(12): 1371-1376.
[15] 王永胜, 陈有, 杨广杰, 等. 探讨圣草酚抑制IL-1β介导的人类骨关节软骨细胞炎症 [J]. 医学新知杂志, 2019, 29(5): 489-493.
[16] 孙晶. 茄根的抗炎活性成分及其质量标准研究 [D]. 北京: 北京中医药大学, 2015: 45-46.
[17] Tubaro A, del Negro P, Ragazzi E,. Anti-inflammatory and peripheral analgesic activity of esculetin[J]., 1988, 20(Suppl 5): 83-85.
[18] Yamada H, Watanabe K, Saito T,. Esculetin (dihydroxycoumarin) inhibits the production of matrix metalloproteinases in cartilage explants, and oral administration of its prodrug, CPA-926, suppresses cartilage destruction in rabbit experimental osteoarthritis [J]., 1999, 26(3): 654-662.
[19] Singh R, Masuda E S, Payan D G. Discovery and development of spleen tyrosine kinase (SYK) inhibitors [J]., 2012, 55(8): 3614-3643.
[20] 单保恩, 董青, 王晓玲, 等. 兔抗Syk抗体制备和在乳腺癌诊断中的应用 [J]. 细胞与分子免疫学杂志, 2003, 19(5): 486-488.
[21] Pine P R, Chang B, Schoettler N,. Inflammation and bone erosion are suppressed in models of rheumatoid arthritis following treatment with a novel Syk inhibitor [J]., 2007, 124(3): 244-257.
[22] Vandoorne K, Magland J, Plaks V,. Bone vascularization and trabecular bone formation are mediated by PKB alpha/Akt1 in a gene-dosage-dependent manner:andMRI [J]., 2010, 64(1): 54-64.
[23] Mukherjee A, Larson E A, Klein R F,. Distinct actions of Akt1 on skeletal architecture and function [J]., 2014, 9(3): e93040.
[24] Mukherjee A, Rotwein P. Selective signaling by Akt1 controls osteoblast differentiation and osteoblast- mediated osteoclast development [J]., 2012, 32(2): 490-500.
[25] Ulici V, Hoenselaar K D, Agoston H,. The role of Akt1 in terminal stages of endochondral bone formation: Angiogenesis and ossification [J]., 2009, 45(6): 1133-1145.
[26] Jiao Y C, Ding H M, Huang S S,. Bcl-XL and Mcl-1 upregulation by calreticulin promotes apoptosis resistance of fibroblast-like synoviocytes via activation of PI3K/Akt and STAT3 pathways in rheumatoid arthritis [J]., 2018, 36(5): 841-849.
[27] 殷悦涵, 孙丽颖, 田文. 雷帕霉素在PIK3CA相关的过度生长(PROS) 的临床应用进展 [J]. 中国骨与关节杂志, 2020, 9(11): 858-863.
[28] 林华, 魏海燕, 蒋青, 等. 原发性骨质疏松骨组织c-fos、c-jun和p53基因的表达 [J]. 中国骨肿瘤骨病, 2003(6): 368-371.
[29] Yamaguchi A, Nozawa K, Fujishiro M,. Estrogen inhibits apoptosis and promotes CC motif chemokine ligand 13 expression on synovial fibroblasts in rheumatoid arthritis [J]., 2012, 34(5): 852-857.
[30] 祝静, 晏波, 蒋瑶, 等. 女性类风湿关节炎患者miR-146a、miR-23b的表达及雌激素对其的影响 [J]. 川北医学院学报, 2020, 35(4): 637-641.
[31] 孙莹, 邱新萍, 孙颂歌. Wnt/β-catenin信号通路在类风湿关节炎中的表达[J]. 世界中医药, 2019, 14(3): 761-765.
[32] Ma H C, Wu W Q, Yang X D,. Genetic effects of common polymorphisms in estrogen receptor alpha gene on osteoarthritis: A meta-analysis [J]., 2015, 8(8): 13446-13454.
[33] Clarkin C E, Gerstenfeld L C. VEGF and bone cell signalling: An essential vessel for communication? [J]., 2013, 31(1): 1-11.
[34] 赵绵松, 夏蓉晖, 王玉华, 等. 骨关节炎与类风湿关节炎患者膝关节滑膜中血管内皮生长因子及血管形态的特征 [J]. 北京大学学报: 医学版, 2012, 44(6): 927-931.
[35] Qin L, Beier F. EGFR signaling: Friend or foe for cartilage? [J]., 2019, 3(2): e10177.
[36] Zan P F, Yao J, Wu Z,. Retracted: cyclin D1 gene silencing promotes IL-1β-induced apoptosis in rat chondrocytes [J]., 2018, 119(1): 290-299.
[37] Sekine C, Sugihara T, Miyake S,. Successful treatment of animal models of rheumatoid arthritis with small-molecule cyclin-dependent kinase inhibitors [J]., 2008, 180(3): 1954-1961.
[38] 秦健, 王志辉, 毕仕强. 绝经后妇女血清基质金属蛋白酶-2、-9与骨转换生化指标及骨密度的关系 [J]. 中国骨质疏松杂志, 2009, 15(4): 292-295.
[39] Wojdasiewicz P, Poniatowski Ł A, Szukiewicz D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis [J]., 2014, 2014: 561459.
[40] 于静雅, 陈勇. PI3K-AKT信号通路及其在类风湿性关节炎滑膜细胞增殖和凋亡中的作用 [J]. 细胞与分子免疫学杂志, 2014, 30(12): 1326-1329.
[41] Westenskow P D, Kurihara T, Aguilar E,. Ras pathway inhibition prevents neovascularization by repressing endothelial cell sprouting [J]., 2013, 123(11): 4900-4908.
[42] Katagiri K, Hattori M, Minato N,. Rap1 functions as a key regulator of T-cell and antigen-presenting cell interactions and modulates T-cell responses [J]., 2002, 22(4): 1001-1015.
[43] Lagarrigue F, Paul D S, Gingras A R,. Talin-1 is the principal platelet Rap1 effector of integrin activation [J]., 2020, 136(10): 1180-1190.
[44] 潘书涵, 王永萍, 王茂林, 等. 基于TNF-α-HIF-1α-iNOS-NO信号通路探讨虎杖大黄素干预类风湿关节炎的作用研究 [J]. 中药药理与临床, 2019, 35(4): 62-67.
[45] 李传洁, 李亚男. 雌激素通过信号通路对成骨细胞分化、成熟与凋亡的调控 [J]. 口腔颌面修复学杂志, 2013, 14(6): 361-365.
[46] 王路, 孙睿, 徐萌, 等. 阿魏化学成分、药理作用及毒理研究进展[J]. 世界中医药, 2020, 15(24): 3887-3894.
[47] 肖培根. 植物亲缘关系、化学成分和疗效间的联系性 [J]. 中国药学杂志, 1978, 13(1): 1-5.
Discriminatory compounds and mechanism ofand three potential substitutes against rheumatoid arthritis
HU Jing1, YANG Yuan-yuan2, REN Hui1, CUI Xiao-min1, LI Ning1, QU Tong, CHEN Zhi-yong1
1. Shaanxi Academy of Traditional Chinese Medicine, Xi’an 710061, China 2. Xi’an Institute for Food and Drug Control, Xi’an 710054, China
To clarify the discriminatory compounds and action mechanism ofCraib and three potential substitutes against rheumatoid arthritis (RA).The discriminatory compounds betweenand three potential substitutes were analyzed by UHPLC-Q Exactive Focus MS/MS. Combined with Swiss target prediction and DisGeNET, OMIM, TTD databases, the common targets for discriminatory compounds in the treatment of RA was obtained. The common targets were imported into the STRING database to construct the network diagram of target interaction, and Cytoscape 3.6.0 software was used to screen out the core targets. The GO and KEGG enrichment analysis of the target were carried out by DAVID database and R language software. The network diagram of Herb-Component-Target-Passway (H-C-T-P) was constructed by Cytoscape 3.6.0 software and analyzed to explore the potential mechanism of the discriminatory components ofand three potential substitutes against RA.A total of 60 components were identified fromand three potential substitutes, including 35 discriminatory components, involving 348 targets. There were 18 components related to RA, and there were 153 common targets for the discriminatory components and RA. GO and KEGG pathway enrichment analysis revealed that five active discriminatory components (formononetin, erycibenin E, eriodictyol,---coumaroyltyramine and esculetin) ofand three potential substitutes acted on AKT1, MAP2K1, PIK3CA and other 32 targets, and may regulate five pathways including PI3K-Akt, Rap1, Ras and so on.The present study preliminarily identified the discriminatory components, key targets and core pathways ofand three potential substitutes in the treatment of RA. The number of common chemical components identified inandand the degree of H-C-T-P network diagram were higher than the other two potential substitutes, which indicate thatis more suitable than other two substitutes. This study provides a relatively reliable strategy for the development of endangered traditional Chinese medicine substitutes.
Benth.; discriminatory compounds; network pharmacology; rheumatoid arthritis;Merr.;Merr.et Chun.;Henmsl.; formononetin; erycibenin E;eriodictyol;--- coumaroyltyramine; esculetin
R284.1
A
0253 - 2670(2022)16 - 4958 - 15
10.7501/j.issn.0253-2670.2022.16.005
2021-02-19
国家自然科学基金面上项目(81973419);陕西省重点研发计划一般项目(2022-SF-315);陕西省中医药管理局青年培优项目(2021-PY-003);陕西省中医药管理局项目(2021-ZZ-JC033);陕西省中医药研究院“苗圃计划”一般项目(2021-12)
胡 静(1988—),女,硕士,主管药师,研究方向为中药质量控制与活性成分研究。Tel: (029)85395696 E-mail: huj668@163.com
陈志永(1987—),男,博士,副研究员,研究方向为中药质量控制与活性成分研究。Tel: (029)85395696 E-mail: chenzy0612@163.com
[责任编辑 王文倩]
