紫茎泽兰入侵对哀牢山森林植物物种多样性的影响
2022-08-12姜倪皓张诗函
姜倪皓,曹 尘,张诗函
1.楚雄师范学院 资源环境与化学学院,云南楚雄 675000;2.楚雄师范学院 教育学院,云南楚雄 675000
紫茎泽兰(Ageratina adenophora)是多年生丛生型半灌木状草本植物,是广泛传播的世界性害草之一,是我国外来入侵物种中危害最严重的植物之一[1-4]。紫茎泽兰不仅与本地种争夺水分、阳光、养分、生存空间,紫茎泽兰还分泌化感物质抑制本地生物的生长,紫茎泽兰入侵严重影响生物群落的结构与功能,打破了群落内物种共存的生态格局[5-7]。
哀牢山位于云贵高原、横断山脉和西藏高原之间,是云南重要的气候与生态类型的分界线,该区域拥有典型的亚热带森林生态系统,生物多样性较丰富,植物区系地理成分复杂[8]。笔者在哀牢山东麓的野外调查中发现,紫茎泽兰早已入侵该区域,处于入侵过程的各个阶段的种群均有发现,紫茎泽兰在该区域呈迅速扩散蔓延的趋势,这将对该地区的生物多样性造成严重威胁。研究紫茎泽兰入侵对哀牢山森林植物物种多样性的影响对植物入侵的防控极为重要。
目前,有关紫茎泽兰对哀牢山亚热带森林群落物种多样性影响的定量调查研究较少。本研究拟通过探讨紫茎泽兰入侵对哀牢山森林群落植物物种多样性的影响,为系统评估紫茎泽兰入侵产生的环境影响和对外来入侵植物的防控提供科学参考。
1 研究区域概况
本研究区域选择在双柏县爱尼山乡安龙哨森林管护点所辖区域,地理坐标为:101°56′E~101°57′E,24°44′N~24°45′N,属于北亚热带高原季风气候,年平均降雨量927 mm,年平均气温15℃。土壤类型以紫色土和红壤为主。样地的森林植被以亚热带常绿阔叶林为主。样地长期维持自然生长发育状态,对紫茎泽兰入侵也未采取任何防治措施。
2 研究方法
2.1 样地设置与调查
本研究选择位于哀牢山东麓的双柏县爱尼山乡的林区为研究区,以林区典型林分为研究对象。在紫茎泽兰分布较多、范围较大的区域内设置样方,设置18个立地类型相似的试验样方(20m×20 m),以紫茎泽兰入侵的样地为试验样方,以紫茎泽兰未入侵的样地为对照样方,对各样方内的乔木层、灌木层、草本层的植物进行统计分析。乔木调查:种类、总株数、树高、胸径、盖度和频度等。灌木层调查:种类、株数、株高度、盖度和频度等。草本层调查:种类、株高、个体数、盖度和频度等。
2.2 重要值计算
本研究选用重要值反映物种在群落中的优势程度,计算公式:
乔木层重要值=(相对盖度+相对高度+相对多度)÷3
灌木层和草本层重要值=(相对密度+相对盖度+相对高度+相对频度)÷4
2.3 物种多样性计算:
Simpson 多样性指数:

Shannon-Wiener 指数:

Pielou均匀度指数:

以上公式中[9-10],Pi为样地中第i物种的重要值,S为物种数。
2.4 数据分析
采用SPSS 19.0软件对野外调查数据进行T检验、回归分析和相关关系检验。
3 结果与分析
3.1 紫茎泽兰入侵与群落特征的关系
群落乔木层盖度和物种丰富度与紫茎泽兰平均株高呈显著负相关关系(P=0.030,P=0.043)(表1)。紫茎泽兰平均株高与灌木层的盖度呈极显著负相关的关系(P=0.009)。灌木层物种丰度、草本层盖度、草本层物种丰度与紫茎泽兰高度和盖度无显著相关。
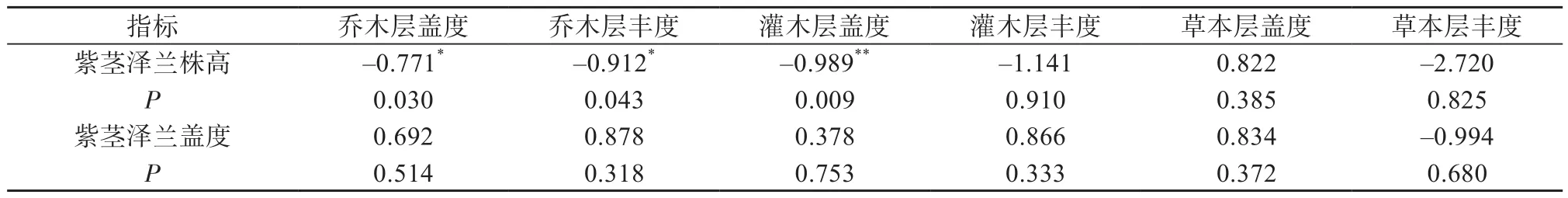
表1 群落植物生活型与紫茎泽兰特征值的相关性
紫茎泽兰的入侵受群落乔木层和灌木层的影响较为显著。乔木层的盖度、乔木层物种丰度以及灌木层盖度增加,紫茎泽兰的平均株高则明显受到抑制。
3.2 重要值分析
在入侵样方的乔木层中,共有物种15个,其中云南樟的重要值最高(重要值0.133))。在试验样方的灌木层中,共有物种4个,重要值最高的物种分别为云南杨梅(0.114)。在试验样方的草本层中,共有物种4个,重要值最高的物种为紫茎泽兰(0.396)。
而在对照样方的乔木层中,共统计到物种5个物种,其中重要值最高的为红花槭(0.126)。灌木层中,共统计到物种7个,重要值最高的物种为野牡丹(0.129)。草本层中,共统计到物种14个,重要值最高的物种分别为里白 (0.284)。
入侵样方中物种重要值集中度高,而对照样方灌木层和草本层的重要值较分散,说明入侵对灌木层和草本层的乡土植物的优势地位起到削减作用,紫茎泽兰入侵影响森林群落物种格局。
3.3 入侵对植物物种多样性的影响
对照样方内灌木层和草本层的Simpson、Shannon-Wiener和Pielou指数均显著高于入侵样方(表3),其中灌木层该指数比入侵样方高128.70%,而草本层高27.34%。对照样方内灌木层和草本层的Shannon-Wiener 指数都显著高于入侵样方,其中灌木层高163.90%,草本层则高出58.77%。对照样方内灌木层和草本层的Pielou指数分别高出试验样方163.90%(P<0.05)和3.58%(P<0.50)。紫茎泽兰入侵对群落整体水平上生物多样性的负效应在灌木层中最高,其次是草本层。将对照样方和入侵样方数据合并,将森林植物群落物种多样性指数、均匀度指数与紫茎泽兰重要值进行拟合(表3)。当紫茎泽兰重要值低于3.27,群落植物多样性指数和均匀度指数都随紫茎泽兰重要值的增加而增加, 而紫茎泽兰重要值高于3.27时, 各项指数都随重要值的增加而减小(图1)。

图1 紫茎泽兰的重要值对群落物种多样性的影响

表3 紫茎泽兰重要值与群落物种多样性指数回归方程
4 讨论
高的资源获取有助于入侵植物的成功入侵[12],光照是影响植物生长发育的重要环境因素[13-14],草本层盖度和乔木层盖度是影响紫茎泽兰生长的限制因子[15]。本研究中,乔木层的盖度、物种丰度以及灌木层盖度和紫茎泽兰的平均株高呈显著负相关的关系,但是草本层相关特征值对紫茎泽兰的平均株高和盖度影响较小。本研究中,乔木层组成物种较对照样方丰富,郁闭度较高,此时光照可能是紫茎泽兰入侵的主导因子,因此常呈成半灌木状的紫茎泽兰与本地灌木层争夺生存空间和资源[16],而与草本植物的竞争可能相对较小。基于本研究结果,对该区域已出现紫茎泽兰危害的林地,可以通过营林来控制该入侵种的扩张。
通常情况下,多样性指数越高,群落的均匀度和丰富度越高,稳定性越好[17]。本研究中,紫茎泽兰入侵的试验样方的灌木层和草本层物种多样性指数和均匀度指数均显著下降。以往研究认为,入侵群落的物种多样性指数和均匀度指数与紫茎泽兰的重要值间都是负相关关系[18]。本研究发现,紫茎泽兰重要值低于0.327时,群落植物多样性指数和均匀度指数都随紫茎泽兰重要值的增加而增加,而紫茎泽兰重要值高于0.327时,各项指数都随重要值的增加而减小。基于前人的研究,推测出因为紫茎泽兰的抗逆性较强,当紫茎泽兰入侵时, 它能改变其生境,进而改善土壤的理化性质,增强物种的多样性。该阶段群落中生长所需资源相对充足,紫茎泽兰与其他灌木层和草本层植物无明显竞争,此时紫茎泽兰对群落灌木层和草本层植物物种多样性产生正向影响。当紫茎泽兰重要值高于0.327时,紫茎泽兰繁殖需要消耗土壤中大量的水分和营养物质,便造成其他灌木层和草本层植物生长所需的水分与营养物质供应不足,最终对其他灌木层和草本层植物物种多样性产生负面影响。此外,可能还与紫茎泽兰的化感作用有关, 紫茎泽兰根系释放的化感物质抑制其他植物的生长,化感作用还改变了入侵生境土壤养分的供求平衡,改变土壤环境,进而加剧该入侵物种的扩散,森林群落的物种多样性下降,最终紫茎泽兰在林下形成单一优势群落。

表2 物种多样性指数比较
5 结论
本研究分析了紫茎泽兰入侵对哀牢山东麓森林群落植物物种多样性的影响。乔木层的盖度、乔木层物种丰度以及灌木层盖度均与紫茎泽兰的平均株高呈显著负相关,该区域紫茎泽兰入侵受森林群落乔木层和灌木层的影响较为显著。紫茎泽兰入侵严重危害群落内灌木层和草本层植物的生存。紫茎泽兰可以对乡土草本和灌木植物物种多样性产生正向和负向双方面的影响, 当紫茎泽兰重要值低于0.327时,随着重要值的增加,物种多样性指数和均匀度指数逐步增加, 但当其重要值高于0.327时,物种多样性指数和均匀度指数随紫茎泽兰重要值的增加而迅速下降。
