烟酸的生物学功能及其在畜禽生产中的应用
2022-08-09梅文亮许兰娇瞿明仁
龙 凡 梅文亮 许兰娇 瞿明仁
(江西农业大学动物科学技术学院,江西省动物营养重点实验室,营养饲料开发工程研究中心,南昌 330045)
烟酸(NA)作为动物机体所必需的维生素之一,具有抗炎、抗氧化、调节脂质代谢[1]、缓解应激(如瘤胃酸应激、热应激、断奶应激[2])等多种生物学功能。烟酸作为辅酶Ⅰ(NAD)和辅酶Ⅱ(NADP)的前体,既在机体碳水化合物、蛋白质和脂类代谢中扮演着重要的角色,又能参与体内能量代谢的调节[3-4]。烟酸缺乏会导致动物生长性能下降、免疫力降低,不利于动物的健康生长。动物可以通过采食饲粮、体内过量色氨酸生物合成、微生物(特别是反刍动物瘤胃微生物)合成等途径获得生长发育所需的烟酸[5]。但是随着动物饲养集约化程度的提高,以及在一些特殊情况(应激状态、高精料饲粮等)下,单靠以上途经获得的烟酸不能满足动物的生产需要,必须在饲粮中额外添加烟酸才能最大限度地发挥其生产潜力,提高饲养效果。本文首先介绍了烟酸的结构和缺乏症,然后重点综述烟酸的生物学功能及其在畜禽生产中的应用,以期为烟酸在畜禽生产中的应用提供参考。
1 烟酸概述
烟酸又称尼克酸,属于水溶性维生素,化学名为吡啶-3-甲酸(C6H5NO2),其化学结构式如图1所示。其结构特征为C5位置带有1个羧基的吡啶环,因此结构简单,理化性质稳定。烟酸在自然界分布甚广,动物中以烟酰胺(NAM)为主,植物中以烟酸为主,但谷物中多是结合型烟酸,不易被吸收利用。动物缺乏烟酸会导致机体代谢紊乱、采食量下降、动物产品质量下降等问题,最终危害动物的生长发育,影响经济效益。王灿宇等[6]用800 mg/kg烟酸的饲粮饲喂夏南牛,结果表明对照组夏南牛的平均日增重、养分消化率低于烟酸组,料重比显著高于烟酸组,由此得出缺乏烟酸会降低夏南牛的生长性能。此外,有研究表明,饲粮中缺乏烟酸会增加肉仔鸡胫骨短促病的发生率,从而使肉仔鸡采食发生严重障碍,导致生长速度下降,甚至死亡[7]。当动物体内烟酸含量过高时,易对机体造成负面影响,甚至危及生命。因此,在实际饲养过程中应该合理利用烟酸。
2 烟酸的生物学功能
2.1 抗炎
烟酸能提高机体的抗炎能力,对维持肠道健康有重要作用。近年来,G蛋白偶联受体109A(GPR109A)被认为与调节微生物群、抑制肠道炎症和维持肠道屏障完整性有关[8]。Singh等[9]以野生型和Niacr1-/-小鼠为试验动物开展试验,结果发现,烟酸以GPR109A依赖性方式增强小鼠结肠巨噬细胞和树突状细胞的抗炎特性,并使它们能够诱导调节性T细胞(Treg)和T细胞[能产生抗炎细胞因子白细胞介素-10(IL-10)]的分化;烟酸还能上调结肠上皮细胞中白细胞介素-18(IL-18)的表达,有利于保护结肠免受炎症和癌变。Salem等[10]研究发现,烟酸以GPR109A依赖性方式,逆转碘乙酰胺诱导的结肠炎小鼠的肿瘤坏死因子-α(TNF-α)、血管内皮生长因子(VEGF)、血管抑制素和内皮抑素水平的升高,使结肠中IL-10水平正常化,从而改善病理性血管生成并缓解结肠炎。Guo等[11]的研究表明,烟酸能够减少小鼠体内的促炎介质,保护乳腺中血乳屏障的完整性;在脂多糖(LPS)诱导的乳腺上皮细胞中,烟酸能够抑制LPS诱导的炎症反应和紧密连接蛋白的下调。此外,烟酸可以将巨噬细胞的极化从促炎状态转变为抗炎状态[12],同时,烟酸可以通过GPR109A减少磷酸化核转录因子-κB(p-NF-κB)的核易位,而核易位的减少则会降低巨噬细胞中促炎细胞因子白细胞介素-1β(IL-1β)、白细胞介素-6(IL-6)的表达,改善血管通透性[13]。Liu等[14]研究发现,饲粮添加烟酸能显著提高断奶仔猪十二指肠转化生长因子-β(TGF-β)、空肠IL-10的蛋白表达,降低回肠白细胞介素-8(IL-8)的蛋白表达,从而减轻肠道炎症;增加结肠中多尔氏菌(Dorea)和乳酸杆菌的相对丰度,降低消化球菌的相对丰度,从而对肠道健康起保护作用。Si等[15]研究发现,烟酸可以减少氧化低密度脂蛋白(oxLDL)刺激的人脐静脉内皮细胞(HUVECs)和人单核细胞THP-1巨噬细胞中炎症细胞因子TNF-α和IL-6的表达,抑制核转录因子-κB(NF-κB)p65和Notch1的蛋白表达,从而抑制炎症反应。此外,烟酸可减少TNF-α诱导的促炎趋化因子单核细胞趋化蛋白-1(MCP-1)、fractalkine和RANTES基因的表达和分泌的增加,增加动脉粥样硬化保护激素脂联素的mRNA表达水平[16]。烟酸可以抑制NF-κB信号通路,导致分离的人单核细胞和视网膜色素上皮细胞中促炎细胞因子和趋化因子TNF-α、IL-6和MCP-1的分泌减少[17-18]。Feng等[19]研究发现,幼龄草鱼缺乏烟酸会降低肠道中溶菌酶(LA)、酸性磷酸酶(ACP)的活性以及补体3(C3)的含量,下调草鱼肠道内肝脏表达抗菌肽-2(LEAP-2)、IL-10、TGF-β1和NF-κB抑制因子α(IκBα)的mRNA表达水平,上调肠道TNF-α、IL-1β、干扰素-γ2(IFN-γ2)、IL-8、IκB激酶(IKK)α、IKKβ和IKKγ的mRNA表达水平,下调封闭蛋白(Claudin)b、Claudin3、Claudinc、闭锁蛋白(Occludin)、闭锁小带蛋白-1(ZO-1)、Claudin15等的mRNA表达水平,降低草鱼肠道黏膜免疫能力和肠道生理功能。
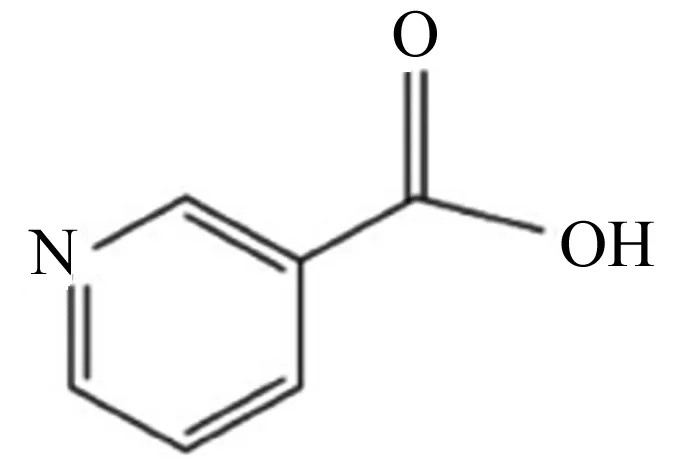
图1 烟酸的化学结构式
2.2 抗氧化
烟酸具有抗氧化特性。段晨磊等[20]研究发现,烟酸可以提高4周龄五龙鹅血清和肝脏谷胱甘肽过氧化物酶(GSH-Px)活性和总抗氧化能力(T-AOC),降低肝脏丙二醛(MDA)和过氧化氢(H2O2)含量;还可以提高16周龄五龙鹅血清总超氧化物歧化酶(T-SOD)和肝脏T-SOD、GSH-Px、过氧化氢酶(CAT)活性,降低血清和肝脏MDA含量。蒋亚军[21]对中国荷斯坦奶牛的十二指肠持续灌注烟酸,结果发现,烟酸可以促进泌乳奶牛血清中T-SOD、GSH-Px等抗氧化酶活性的提高,降低硫代巴比土酸反应物(TBARS)和MDA含量,从而减少脂质过氧化物反应。有研究表明,烟酸缺乏会增加肠道中活性氧(ROS)和MDA含量,降低抗氧化酶铜/锌超氧化物歧化酶(Cu/Zn-SOD)、GSH-Px、锰超氧化物歧化酶(MnSOD)、CAT、谷胱甘肽-S-转移酶(GST)和谷胱甘肽还原酶(GR)在幼鱼肠道中的活性,对草鱼抗氧化系统产生不利影响[19]。
核因子E2相关因子2(Nrf2)是存在于细胞质中的转录因子。在生理条件下,Keap1与Nrf2结合,并在细胞质中隔离Nrf2。在氧化应激或存在Nrf2激活化合物的情况下,Nrf2基因表达量上调,Nrf2易位到细胞核并调节各种抗氧化反应元件(AREs)基因的表达,包括血红素加氧酶-1(HO-1)、NAD(P)H醌脱氢酶1(NQO1)、GST,因此,Nrf2的激活被认为是对抗氧化应激损害效应的保护机制之一[22-23]。Wu等[24]发现烟酸可以减少肺缺血再灌注(I/R)损伤的大鼠的MDA含量,降低肺部氧化应激;降低TNF-α的含量,缓解肺部炎症;提高沉默信息调节因子1(SIRT1)的含量,缓解肺部氧化应激和炎症。Hu等[25]研究发现,用烟酸预处理被对乙酰氨基酚(APAP)攻击的BALB/c小鼠可以降低血清中丙氨酸氨基转移酶(ALT)和天冬氨酸氨基转移酶(AST)活性,并减轻组织病理学损伤;提高肝脏中抗氧化酶SOD、CAT的活性和谷胱甘肽(GSH)的含量,降低MDA含量,从而减轻脂质过氧化和氧化应激;同时,烟酸上调肝脏中SIRT1、Nrf2、NQO1的蛋白表达水平并下调Keap1的蛋白表达水平。烟酸还可以抑制甲基汞(MeHg)中毒引起的雄性Wistar大鼠线粒体氧化应激[26]。Chen等[27]研究发现,烟酸可以抑制大鼠肺氧化应激和炎症,防止胰腺缺血再灌注(PIR)诱导的急性胰腺炎和肺屏障功能障碍,同时恢复SIRT1的mRNA表达。另外,有研究表明,烟酸及其主要代谢物烟酰胺能够显著降低LPS处理的人成熟巨噬细胞中ROS、一氧化氮(NO)和一氧化氮合酶2(NOS2)的表达,还能使巨噬细胞极化倾向于抗炎的M2巨噬细胞,从而发挥抗氧化和抗炎作用[28]。
2.3 调节脂质代谢
烟酸有良好的降脂作用,可以降低血脂异常和糖尿病患者血清中总胆固醇(TC)、甘油三酯(TG)、低密度脂蛋白胆固醇(LDL-C)的含量,提高高密度脂蛋白胆固醇(HDL-C)的含量,改善血脂状况[29-31]。Galescu等[32]研究表明,烟酸能显著降低肥胖儿童血清游离脂肪酸(FFA)含量并增加生长激素(GH)含量,有利于其生长发育。杨玲[33]的研究表明,烟酸能减少大鼠肝细胞(BRL3A)内TG的合成;能够通过激活蛋白激酶A(PKA),提高其磷酸化水平而降低碳水化合物反应元件结合蛋白(ChREBP)表达水平及入核效率,从而降低乙酰辅酶A羧化酶(ACC)、脂肪酸合成酶(FAS)、甘油二酯酰基转移酶(DGAT2)蛋白表达水平,减轻脂肪变性程度。Wang等[34]研究表明,0.5 mmol/L烟酸通过GPR109A激活AMP依赖的蛋白激酶(AMPK)/ACC信号通路并抑制AMPK/哺乳动物雷帕霉素靶蛋白(mTOR)/核糖体S6K蛋白激酶(S6K)信号通路,从而减少牛乳腺上皮细胞(BMECs)中乳脂的合成以及FAS和固醇调节元件结合蛋白1(SREBP1)蛋白的表达。有研究表明,烟酸能通过GPR109A介导的蛋白激酶C(PKC)-细胞外信号调节激酶1/2(ERK1/2)-AMPK信号通路抑制C57BL/6小鼠的肝脏脂肪从头生成(DNL)来改善肝脏脂肪变性[35]。Zimmer等[36]发现,烟酸和二十碳五烯酸的组合可以抑制小鼠肝细胞中SREBP1的活性,减少TG的沉积。
烟酸能通过与GPR109A结合,降低PKA、腺苷酸环化酶(AC)、激素敏感酯酶(HSL)等酶的活性,从而抑制TG分解并降低血液FFA含量[37-38];烟酸在肝脏中通过抑制DGAT2活性来减少低密度脂蛋白(LDL)、极低密度脂蛋白(VLDL)和TG的合成与分泌[1]。有研究表明,烟酸诱导的肝脏FFA供应减少可以抑制肝脏中过氧化物酶体增殖物激活受体γ辅活化因子-1β(PGC-1β)和载脂蛋白C3(APOC3)的表达,这将减少VLDL的合成和分泌,并增加其周转率,从而达到降血脂作用[39]。此外,烟酸可通过蛋白激酶B(AKT)/叉头框转录因子1(FoxO1)信号通路调控一系列能量代谢和细胞信号通路基因表达[40],还可通过肾上腺素、脂联素、瘦素、胰岛素等激素内分泌系统来调控动物体内糖、脂代谢,从而促进脂肪细胞分化和脂质沉积[41-43]。
2.4 改善肉品质
肉品质是个复杂的性状,难以用单个指标去衡量,一般评价肉品质的决定性指标包括肉色、嫩度、多汁性、风味、营养价值等,而这些指标与肌肉组织中肌纤维类型和肌内脂肪沉积密切相关。目前,根据肌球蛋白重链亚型(MyHC)可以将肌纤维分成4种:慢速氧化型(Ⅰ型)、快速氧化型(Ⅱa型)、中间型(Ⅱx型)和快速糖酵解型(Ⅱb型)[44]。王勇峰等[45]研究了不同部位新疆褐牛肌纤维特性与肉品质的差异,结果表明,Ⅰ型、Ⅱa型和Ⅱb型肌纤维面积、直径与剪切力、硬度和蒸煮损失成正比;Ⅰ型肌纤维比例与硬度、剪切力成反比,Ⅱb型肌纤维比例与剪切力成正比,所以增加Ⅰ型肌纤维比例、减少Ⅱb型肌纤维比例均有利于提高牛肉嫩度。此外,有研究发现,Ⅱb型肌纤维含量高的肌肉在贮藏过程中,肌纤维易发生收缩和分离,断裂程度更严重,导致肌肉失水严重,持水力低[46]。Khan等[47]在11周龄杂交猪饲粮中添加750 mg/kg烟酸后发现,烟酸能够提高猪的背最长肌、股四头肌和腓肠肌的Ⅰ型肌纤维百分率,降低Ⅱ型肌纤维百分率,促进Ⅱ型肌纤维向Ⅰ型转化,影响肉品质;显著提高猪背最长肌中纤维转化的关键调节因子(PGC-1β)、线粒体脂肪酸分解代谢[肉毒碱脂酰转移酶(CACT)、脂肪酸转运蛋白1(FATP1)、有机阳离子转运体2(OCTN2)]、三羧酸循环[琥珀酸脱氢酶复合体亚单位A(SDHA)]、氧化磷酸化[细胞色素c氧化酶亚基4/1(COX4/1)、细胞色素c氧化酶亚基6A1(COX6A1)]和产热[解偶联蛋白3(UCP3)]相关基因的表达水平,而这些基因都在Ⅰ型肌纤维中大量表达,表明烟酸能够诱导背最长肌的Ⅰ型肌纤维含量增加,从而改善肉品质。Khan等[48]研究发现,烟酸可以显著提高绵羊背最长肌(LD)、半膜肌(SM)和半腱肌(ST)中的Ⅰ型肌纤维的数量百分比,降低Ⅱ型肌纤维的数量百分比,还能提高半膜肌和半腱肌中的纤维转化调节因子过氧化物酶体增殖激活受体δ(PPARδ)、过氧化物酶体增殖物激活受体γ辅活化因子-1α(PGC-1α)和PGC-1β的mRNA表达水平以及参与线粒体脂肪酸摄取[肉碱棕榈酰转移酶1B(CPT1B)、肉碱-酰基肉碱转位酶(SLC25A20)]、三羧酸循环(SDHA)、线粒体呼吸链[细胞色素c氧化酶亚基5A(COX5A)、COX6A1]和血管生成[血管内皮生长因子A(VEGFA)]等基因的mRNA表达水平,说明烟酸能够诱导肌肉纤维从Ⅱ型转变为Ⅰ型,增强骨骼肌利用脂肪酸的能力。
肌内脂肪(IMF)又称大理石花纹脂肪,存在于肌肉内部,与肉品质密切相关。有研究表明,IMF含量增加,肌肉中的不饱和脂肪酸(UFA)、醛类、醇类和碳氢化合物等风味化合物水平增加,其风味、多汁性、嫩度更好[49]。IMF的沉积包括脂肪的合成、分解与转运,在这个过程中,起关键性作用的是与脂肪代谢相关的一些酶、激素、基因和通路。AMPK、过氧化物酶体增殖物激活受体(PPARs)、脂肪酸结合蛋白(FABPs)、脂滴包被蛋白1(PLIN1)等调控因子在脂肪代谢的调控上发挥着重要作用[50-51]。研究表明,烟酸能够激活AMPK[35];而Luo等[52]研究发现,AMPKα磷酸化能够抑制乙酰辅酶A羧化酶2(ACC2)的活性并提高肉碱棕榈酰转移酶1(CPT1)的活性,促进脂肪酸的氧化,还能上调FAS和硬脂酰辅酶A去饱和酶(SCD)的mRNA表达水平,增加肥育猪骨骼肌脂肪生成能力,从而增加IMF的积累和单不饱和脂肪酸(MUFA)的合成,这可能是烟酸促进IMF沉积的原因之一。Zhang等[53]研究表明,烟酸能够显著提高五龙鹅胸肌的pH和持水能力,降低亮度(L*)值和剪切力,并且极显著增加IMF含量和肌间脂肪带的宽度,从而提高五龙鹅的肉品质,改善其胴体性状。有研究表明,烟酸可以提高NADPH合成酶如葡萄糖-6-磷酸脱氢酶(G6PDH)和异柠檬酸脱氢酶(ICDH)在脂肪酸从头合成过程中的活性,同时促进脂肪细胞分化和前脂肪细胞脂滴形成,从而增加肉牛背最长肌IMF的沉积[43,54]。
2.5 缓解奶牛乳腺炎
目前,奶牛乳腺炎严重阻碍了全球奶业发展[55]。病原体入侵乳腺后产生大量的内毒素(主要成分是LPS),它能通过与Toll样受体4(TLR4)结合激活NF-κB,刺激促炎细胞因子(如IL-6和IL-1β)的表达,并诱导强烈的免疫反应[56-57],还能触发BMECs或巨噬细胞的免疫反应,从而释放大量的促炎因子,破坏BMECs,导致乳腺炎[58-59]。大量研究表明,乳腺炎严重影响奶牛的产奶量和奶品质[60],因此缓解乳腺炎是提高奶牛产奶量和奶品质的重要手段。Guo等[61]研究表明,烟酸能显著提高乳腺的先天免疫水平,降低奶牛乳汁和血清中IL-6、TNF-α和IL-1β的含量;可以激活GPR109A并磷酸化AMPK,促进Nrf2核导入和自噬,从而减轻LPS诱导的BMECs炎症反应,缓解奶牛的乳腺炎。此外,巩倩[62]的研究也表明,烟酸能够降低奶牛血清和乳汁中IL-6、TNF-α和IL-1β的含量,提高免疫力,还能通过GPR109A/AMPK/Nrf2信号通路激活自噬过程而起到抗炎作用,减轻奶牛乳腺炎的程度。
2.6 其他功能
烟酸能够促进动物的正常生长发育,是动物生命过程中不可缺少的维生素之一。烟酸是NAD和NADP的前体,而NAD和NADP作为重要辅酶,在氧化还原过程中起传递氢和电子的作用,参与细胞中氧化磷酸化、糖酵解和DNA修复等过程[3]。有效的NAD+/NADH的比值能够调节糖酵解、三羧酸循环、脂肪酸氧化和脂肪酸合成等关键酶的活性,这些酶是细胞新陈代谢的重要途径[63]。NAD+的可用性降低会阻止线粒体去乙酰化酶(Sirt3-5)的最佳活性,而Sirt3-5对NAD+的依赖使它们能够充当代谢传感器,感知细胞的代谢状态并将这些信息转化为线粒体调节,而线粒体去乙酰化酶的活性与预防与年龄相关的神经病变、胰岛素抵抗和心脏病等有关[64]。因此,烟酸对动物机体营养物质代谢、能量代谢以及健康等方面有重要作用。
烟酸除以上功能外,还有一些其他作用,如:1)提高肥胖动物内皮细胞(EC)的血管生成功能[65];2)提高饲料转化率,提高畜禽的生产性能[66-67];3)通过影响瘤胃微生物蛋白(MCP)、氨态氮(NH3-N)和总挥发性脂肪酸(TVFA)的含量来改善瘤胃发酵[68]。此外,烟酸在提高动物抗应激能力方面也起着至关重要的作用。烟酸可以通过引起血管舒张,加快血液向皮肤表面流动,从而促进组织散热,缓解机体热应激[69]。
3 烟酸在畜禽生产中的应用
3.1 在家禽生产中的应用
Jiang等[67]研究了饲粮中不同添加水平的烟酸对3种基因型鸡[爱拔益加(AA)肉鸡、IMF上选系北京油鸡(BJY+IMF)和IMF下选系北京油鸡(BJY-IMF)]生长性能、胴体性状和肉品质的影响,研究发现,饲粮添加60 mg/kg烟酸时,3种基因型鸡的平均日增重、平均日采食量、肌间脂肪带宽度、皮下脂肪(包括皮肤)厚度和腹脂率均有增加趋势,同时饲喂玉米-豆粕型饲粮的AA肉鸡的烟酸适宜添加量为60 mg/kg,能最大限度地提高平均日增重和肉品产量,并降低胸肌滴水损失。刘政等[70]用含60、80、100、120 mg/kg烟酸的饲粮饲喂AA肉仔鸡,研究发现,不同浓度的烟酸均能降低肉仔鸡血清TC和LDL-C含量,提高血清葡萄糖(GLU)和HDL-C含量,且随着烟酸浓度的提高,血清TG含量先上升后下降,从而改善血脂状况;同时,不同浓度的烟酸均对肉鸡血清SOD、谷胱甘肽过氧化物酶(GSH-Px)和CAT活性有提高作用,能提高肉仔鸡的抗氧化性能。Zhang等[53]研究表明,烟酸能够以线性方式增加五龙鹅血清脂肪酶活性和肝脏脂蛋白脂肪酶mRNA表达水平,并对血清TG含量和苹果酸脱氢酶(MDH)活性产生影响,从而调节脂质代谢。刘晨龙等[71]研究表明,饲粮中添加40 mg/kg烟酸可以降低产蛋期种鹅的料蛋比,提高蛋壳厚度和蛋壳强度,改善蛋品质,还能降低血清中TG、TC、LDL-C含量,提高血清中HDL-C含量,调节脂质代谢。Adebowale等[72]研究发现,饲粮中添加180 mg/kg烟酸降低了火鸡雏鸡的采食量和血清ALT、ALP活性与LDL、TG和TC含量,提高了火鸡的增重、代谢能和血清高密度脂蛋白(HDL)含量,因此,在不影响火鸡增重的情况下,添加烟酸可以降低火鸡的脂肪含量,提高火鸡的抗应激能力。王静等[73]研究发现,饲粮含80 mg/kg烟酸组罗斯308肉仔鸡的活重和屠体重相比对照组增加了7.87%和7.50%;同时,烟酸显著提高了胸肌和腿肌的IMP含量,促进了脂肪沉积;显著提高了肉色评分,降低胸肌和腿肌的滴水损失率和剪切力,从而改善了肉品质。
综上可知,在家禽饲粮中添加烟酸能够改善家禽的血液指标,提高机体抗氧化能力,调节脂质代谢,改善肉品质和胴体性状,并提高其生长性能。
3.2 在猪生产中的应用
Almubarak等[74]研究表明,烟酸能显著提高猪卵母细胞的GSH-Px活性,降低ROS含量,并对卵丘细胞和卵母细胞中与能量和氧化状态(SIRT1)、抗凋亡[B淋巴细胞瘤-2(Bcl-2)]、促凋亡[Bcl-2相关X蛋白(Bax)]和脂质代谢[ACC和含patatin样磷脂酶结构域蛋白2(PNPLA2)]相关的基因的表达产生积极影响,保护机体免受氧化应激,改善早期胚胎发育能力;提高囊胚总细胞数、卵裂和囊胚率,促进胚胎发育。此外,Pollard等[75]研究也表明,烟酸能够促进猪卵母细胞的体外成熟(IVM),促进囊胚的形成,对胚胎发育起积极作用。Feng等[76]研究表明,烟酸可显著降低断奶仔猪的腹泻指数和体重损失;改善空肠绒毛高度和肠道形态评分;显著增加空肠抗菌肽[猪β防御素2(pBD2)和PR39]、ZO-1和Occludin的mRNA表达;改善结肠内容物中微生物菌群组成和乙酸盐浓度;显著降低结肠内容物中丙酸盐/乙酸盐和丁酸盐/乙酸盐的比率;显著增加SIRT1的蛋白表达并抑制组蛋白脱乙酰基酶7(HDAC7)的蛋白表达。Liu等[14]研究发现,烟酸能显著增加断奶仔猪十二指肠的绒毛高度以及绒毛高度与隐窝深度的比值,同时降低隐窝深度,从而增强肠道对营养物质的吸收能力,改善肠道上皮细胞的分泌功能。Real等[77]在玉米-豆粕型基础饲粮中添加烟酸,结果显示烟酸能够提高育肥猪的平均日采食量,改善生长性能;能提高背最长肌的pH24 h,显著降低胴体收缩率,改善肉色评分,提高肉品质。
综上可知,在猪生产中应用烟酸能够提高早期胚胎的发育能力,改善肠道结构,提高机体免疫能力,提高肉品质和生长性能。
3.3 在反刍动物生产中的应用
3.3.1 奶牛
Gaowa等[78]选取12头经产荷斯坦奶牛,在饲粮中分别添加0、20 g/d的过瘤胃烟酸(RPN),研究表明,烟酸能够显著提高产犊后前3周内奶牛的干物质采食量(DMI)和产奶量;显著提高血液中神经肽Y(NPY)和食欲肽A(OXA)的含量,提高产后荷斯坦奶牛的采食欲望。Aragona等[79]研究表明,在产前荷斯坦奶牛饲粮中添加烟酸能够增加初乳中免疫球蛋白G(IgG)和其他营养物质(如蛋白质、脂肪等)的含量,为犊牛提供更高质量的初乳,从而减少犊牛发病率和死亡率;能够提高尿囊素和总嘌呤衍生物的含量,表明烟酸能增加MCP的含量;还能够提高犊牛的饲料转化率,其中添加32 g/d烟酸时提高犊牛生产性能的效果最好。此外,有研究表明,产前给母牛补饲48 g/d烟酸,能够将初乳中IgG的含量从73.8 g/L增加到86.8 g/L,增加了18%,很大程度上改善了初乳质量[80]。Guo等[81]研究发现,烟酸能够显著降低泌乳荷斯坦奶牛血清中热休克蛋白70(HSP70)、促肾上腺皮质激素(ACTH)、IL-6含量和乳酸脱氢酶(LDH)活性,缓解热应激;显著提高奶牛的产奶量、能量校正乳(ECM)产量和DMI,提高乳脂水平,改善奶牛的生产性能和乳品质。陈志远等[82]发现,以围产期奶牛为试验动物,在饲粮中添加20 g/(头·d)过瘤胃烟酸能够显著提高奶牛产奶量,减少血清中β-羟丁酸(BHBA)和非酯化脂肪酸(NEFA)的含量,并减轻能量负平衡的程度,抑制脂质动员,减少患酮病的风险。Zeitz等[83]研究发现,烟酸可以降低泌乳早期奶牛血浆中BHBA含量,降低肝脏中酮生成调节因子成纤维细胞生长因子21(FGF21)的基因表达水平,从而降低患酮血症的风险。
综上可知,在奶牛生产中应用烟酸能够提高奶牛的产奶量和乳品质,预防奶牛酮血症发生,缓解热应激。
3.3.2 肉牛
杨艳等[68]体外研究发现,烟酸可以提高锦江牛瘤胃液的pH以及MCP、VFA、丙酸含量和总产气量,降低乙酸/丙酸比值,改善瘤胃的发酵功能,其中以精粗比为6∶4时添加400 mg/kg烟酸以及精粗比为7∶3、8∶2时添加800 mg/kg烟酸的效果较好。Luo等[84]发现,高精料饲粮会诱导亚急性瘤胃酸中毒(SARA),降低牛瘤胃细菌群落的α多样性,而添加800 mg/kg的烟酸能够提高瘤胃液pH,降低普雷沃氏菌属(Prevotella)的相对丰度,增加解琥珀酸菌属(Succiniclasticum)、醋弧菌属(Acetivibrio)和密螺旋体属(Treponema)的相对丰度,改变瘤胃微生物组成,缓解牛的SARA症状。此外,在高精料饲粮中添加烟酸可以增加锦江牛瘤胃NAD+浓度,降低NADH浓度,重新抑制LDH活性,减少乳酸和丙酮酸蓄积,并且能改善固、液相菌群功能,缓解瘤胃酸中毒[85]。王雪莹[66]的研究表明,热应激期在饲粮中添加烟酸能够显著提高牦牛的DMI、平均日增重以及粗脂肪和粗蛋白质的消化率,提高生长性能。Yang等[54]选取24月龄的公牛,在基础饲粮中添加1 000 mg/kg的烟酸,结果表明,饲粮中添加烟酸能够增加背最长肌的IMF含量、眼肌面积、大理石花纹评分、红度(a*)值和色饱和度(C*)值,降低胴体脂肪(不包括IMF)含量,促进肌肉内脂肪沉积,提高肉品质,改善胴体性状;增加血清中HDL-C含量,降低LDL-C、TG、NEFA、TC和糖化血清蛋白(GSP)的含量,调节血清代谢物的组成,改善脂质代谢。此外,烟酸还可以减轻小牛长途运输带来的不利影响[86-87]。
综上可知,在肉牛生产中应用烟酸能够改善肉牛的瘤胃发酵功能,改变瘤胃菌群组成,促进脂肪沉积,缓解应激并提高生产性能。
3.3.3 羊
Luo等[88]研究表明,20~80 mmol/L烟酸可减轻丁酸盐诱导的绵羊瘤胃上皮细胞损伤并促进增殖,40 mmmol/L烟酸呈现出最佳效果,而较高浓度(100 mmol/L)的烟酸会导致低细胞活力;40 mmol/L烟酸能显著降低细胞内ROS含量并增加细胞内T-AOC,缓解丁酸盐诱导的氧化应激;40 mmol/L烟酸能够使凋亡细胞的比例降低,下调细胞凋亡相关基因半胱氨酸天冬氨酸蛋白酶-3(Caspase-3)和p53的mRNA表达水平并上调Bcl-2和Bax的mRNA表达水平,保护瘤胃上皮细胞免受丁酸盐诱导的细胞凋亡。茹婷[89]的研究表明,添加烟酸对围产后期母羊的乳糖率和乳脂率有显著影响,还可以增加母羊围产后期血液中GLU的含量;添加4 g/(只·d)烟酸可显著降低母羊血液中NEFA含量,减轻能量负平衡(NEB);可以通过胰岛素信号转导,调节AKT/磷酸肌醇-3激酶(PI3K)-FoxO1通路,调控糖异生。刘秀梅[90]研究了不同剂量烟酸对羊肝细胞的作用,结果表明,烟酸可以提高羊肝细胞的增殖率;可以显著提高肝细胞糖异生关键酶磷酸烯醇式丙酮酸羧激酶(PEPCK)、丙酮酸羧化酶(PC)的活性,且烟酸浓度为10-4mol/L时效果最强;还能影响肝脏糖异生[如葡萄糖-6-磷酸酶(G6P)、PEPCK、PC]及P53-SIRT6-FoxO1轴基因的mRNA表达水平。
综上可知,在羊生产中应用烟酸能够提高乳品质,缓解细胞损伤并促进细胞增殖。
4 小 结
烟酸是畜禽生长的必需维生素,具有多种生理功能。它不仅可以作为饲料添加剂提高畜禽的抗炎、抗氧化能力,促进生长,改善肉品质和胴体性状,提高泌乳性能,还可以起到药物的效果,治疗血脂异常、乳腺炎等疾病,从而提高畜禽的抗病能力,减少死亡率,最终提高养殖效益。然而,烟酸发挥生物学功能的具体作用机理尚未充分阐明,在这些过程中涉及的酶、激素、转录因子和信号通路也需要进一步地研究并阐明。此外,关于烟酸对不同性别、不同生长阶段、不同品种的动物的作用效果以及烟酸在不同动物中的添加量和添加形式的研究较少,因此,这可以作为今后的研究方向之一。
