水产动物苯丙氨酸研究进展
2022-08-09王连生
吴 迪 王 良 王连生*
(1.中国水产科学研究院黑龙江水产研究所,黑龙江省水生动物病害与免疫重点实验室,哈尔滨 150070;2.潍坊加易加生物科技有限公司,潍坊 261100)
苯丙氨酸(phenylalanine,Phe)是鱼类不能靠自身代谢合成的几种必需氨基酸之一,属芳香族氨基酸。在人体内,Phe能够直接合成重要的神经递质和激素,参与人体供能所必需的糖脂代谢活动[1-2];在植物体内,Phe可对细胞器和组织发育起到直接作用,促进植物细胞的生长与信号转导[3];在医药领域,Phe还可作为抗癌药物的载体将药物分子直接导入癌瘤区,有效抑制癌细胞的分化,减少药物的毒副作用[4];在食品加工领域,Phe能够和糖类等物质发生一系列酰胺化反应,改善食物风味,维持人体日常营养所需的氨基酸平衡[5-7]。然而,Phe在水产动物中的生理活性机制尚不明晰。本文综述了Phe的合成与代谢、水产动物需求量及其在水产养殖中的应用研究进展,以期为Phe在水产动物的营养需求、代谢机制与绿色养殖等方面的进一步研究提供重要基础。
1 Phe的合成与代谢
1.1 Phe的生物合成
Phe的生物合成是一个极其复杂的过程,且只能由植物或微生物合成。Phe的合成起始物质是赤藓糖-4-磷酸(E4P)和磷酸烯醇式丙酮酸(PEP),二者经过一系列氧化、脱氢、脱羧、分子重排与缩合,最终生成莽草酸。莽草酸是Phe合成的前体物质。以莽草酸为起始物,经多种酶作用并最终形成分支酸的过程,称为莽草酸途径[8]。这一途径是芳香族氨基酸合成的共同步骤。随后,作为芳香族氨基酸合成途径的分支点,在分支酸变位酶与预苯酸脱水酶的作用下,先后转变成预苯酸和苯丙酮酸,最后在转氨酶的作用下形成Phe[9](图1)。
1.2 Phe的代谢
Phe在生物体内的主要代谢途径是通过羟化反应生成酪氨酸(tyrosine,Tyr)[10](图2)。Phe可在辅酶四氢生物喋呤的作用下不可逆地转化为Tyr[11]。当体内苯丙氨酸羟化酶发生缺陷时,Phe不能正常转变为Tyr,Phe在转氨基作用下生成苯丙酮酸、苯乙酸等,且随尿排出,俗称苯酮酸尿症(phenyl keronuria,PKU),是最常见的氨基酸代谢缺陷[12-13]。Phe缺乏时,会影响动植物体内Tyr合成,并造成甲状腺素水平下降,影响动植物的正常代谢活动,严重时可导致智力低下、癫痫等问题[14]。
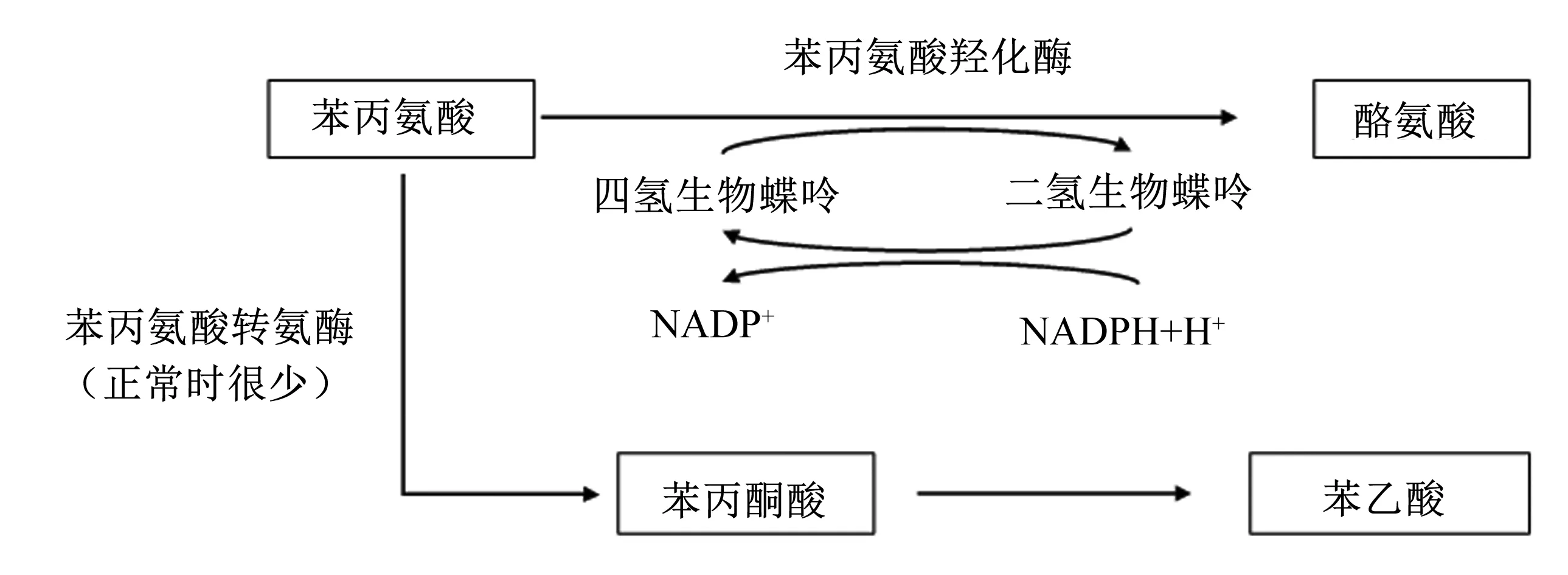
图2 苯丙氨酸的代谢
2 水产动物对Phe的需求量
不同种类水产动物对Phe的需求量见表1。由表可知,水产动物对Phe需求量的范围变化较大。鱼类对Phe的需求量为0.47%~3.66%,对Phe+Tyr的需求量为1.20%~3.80%;虾蟹类对Phe的需求量为1.30%~1.58%。水产动物对Phe的需求量与其种类、大小、饲料类型、饲养管理条件、投喂方法、饲料氨基酸组成、评价指标和统计方法等密切相关。同一品种不同饲养规格的水产动物对饲料中Phe的需求量不同。当草鱼(Ctenopharyngodonidella)体重为255.97 g时,其对Phe的需求量为饲料干重的1.04%[15],而体重为13.21 g的草鱼对Phe的需求量为饲料干重的1.22%~1.27%[16]。当体重为158.78和1.65 g时,尼罗罗非鱼(Oreochromisniloticus)对Phe的需求量分别为1.21%[17]和3.66%[18]。在露斯塔野鲮(Labeorohita)的研究中,体重为0.18 g的露斯塔野鲮对Phe的需求量为1.16%~1.22%[19]。体重为0.92 g的麦瑞加拉鲮(Cirrhinusmrigala)对Phe的需求量为1.75%[20]。当体重为0.58 g时,遮目鱼(Chanoschanos)对Phe的需求量为1.26%~1.90%[21];当体重达到8.00 g时,其对Phe的需求量为1.12%~1.69%[22]。在日本鳗鲡(Anguillajaponica)对Phe需求量的研究中,黑子鳗为1.89%,幼鳗为1.76%,成鳗为1.60%[23],以上需求量均低于NRC(2011)中日本鳗鲡成鳗在正常饲养水平下的必需氨基酸推荐值。评价指标不同,所得水产动物对Phe的需求量也存在一定差异。在草鱼的研究中表明,以增重率、饲料效率和蛋白质沉积率为评价指标,通过折线回归分析模型得出,体重为13.21 g的草鱼对Phe的需求量分别为1.27%、1.22%和1.26%[16]。Kim等[24]以红鲷(Pagrusmajor)为研究对象,评价了Phe二肽形式作为一种新型氨基酸来源的效果,结果表明,二肽Phe对红鲷幼鱼的利用与自由形态一样有效,对其生长性能和全身氨基酸组成均无不良影响。Baker[25]在对不同统计方法进行比较分析后得出,通常条件下由折线法计算得出的氨基酸需求量往往比非线性方法估计得出的氨基酸需求量低。

表1 水产动物苯丙氨酸需求量(以饲料百分比计)
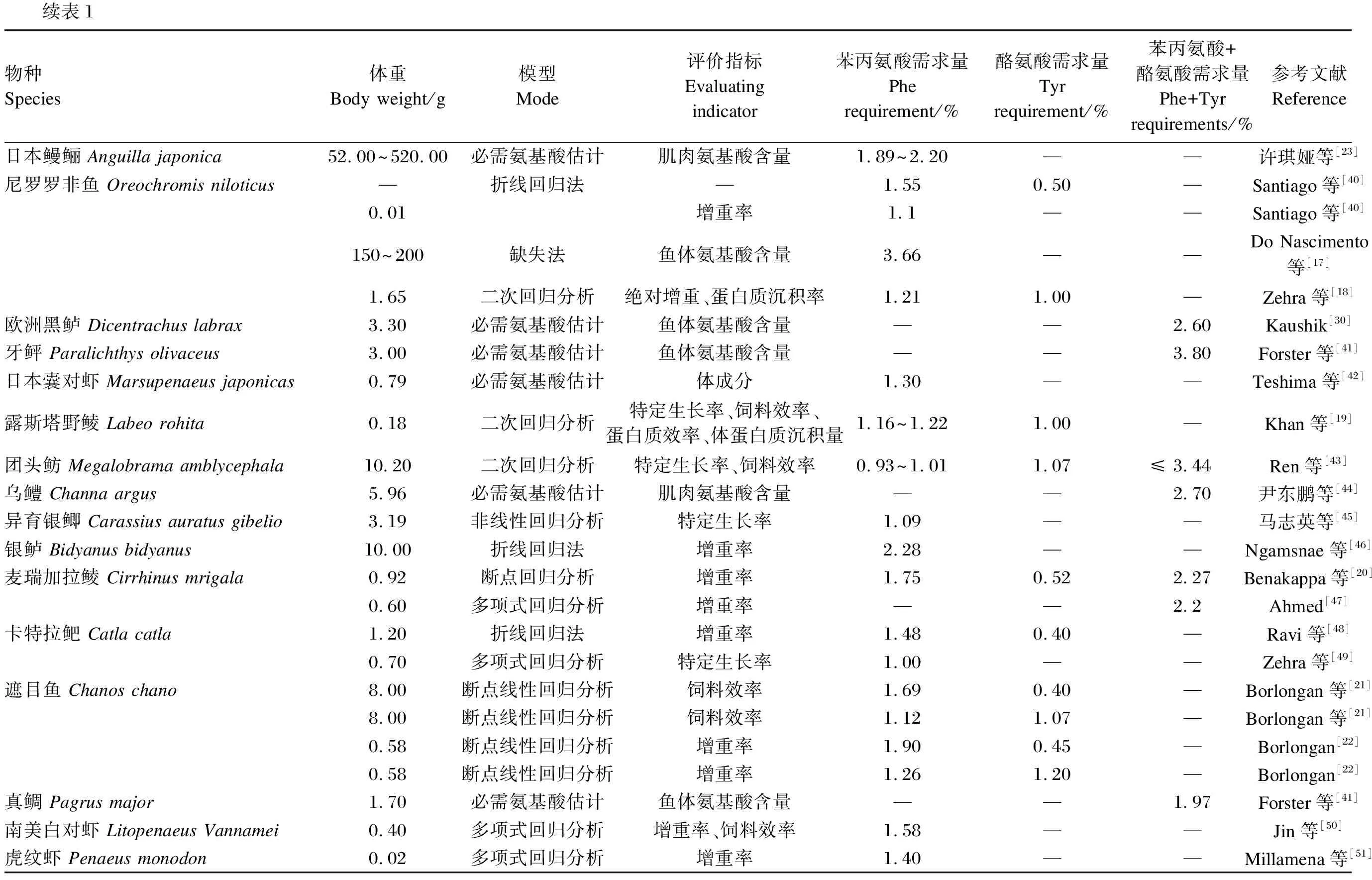
续表1物种Species体重Body weight/g模型Mode评价指标Evaluating indicator苯丙氨酸需求量Phe requirement/% 酪氨酸需求量Tyr requirement/% 苯丙氨酸+酪氨酸需求量Phe+Tyr requirements/% 参考文献Reference日本鳗鲡 Anguilla japonica52.00~520.00必需氨基酸估计肌肉氨基酸含量1.89~2.20——许琪娅等[23]尼罗罗非鱼 Oreochromis niloticus—折线回归法—1.550.50—Santiago等[40]0.01增重率1.1——Santiago等[40]150~200缺失法鱼体氨基酸含量3.66——Do Nascimento等[17]1.65二次回归分析绝对增重、蛋白质沉积率1.211.00—Zehra等[18]欧洲黑鲈 Dicentrachus labrax3.30必需氨基酸估计鱼体氨基酸含量——2.60Kaushik[30]牙鲆 Paralichthys olivaceus3.00必需氨基酸估计鱼体氨基酸含量——3.80Forster等[41]日本囊对虾 Marsupenaeus japonicas0.79必需氨基酸估计体成分1.30——Teshima等[42]露斯塔野鲮 Labeo rohita0.18二次回归分析特定生长率、饲料效率、蛋白质效率、体蛋白质沉积量1.16~1.221.00—Khan等[19]团头鲂 Megalobrama amblycephala10.20二次回归分析特定生长率、饲料效率0.93~1.011.07≤ 3.44Ren等[43]乌鳢 Channa argus 5.96必需氨基酸估计肌肉氨基酸含量——2.70尹东鹏等[44]异育银鲫 Carassius auratus gibelio3.19非线性回归分析特定生长率1.09——马志英等[45]银鲈 Bidyanus bidyanus10.00折线回归法增重率2.28——Ngamsnae等[46]麦瑞加拉鲮 Cirrhinus mrigala0.92断点回归分析增重率1.750.522.27Benakappa等[20]0.60多项式回归分析增重率——2.2Ahmed[47]卡特拉鲃 Catla catla1.20折线回归法增重率1.480.40—Ravi等[48]0.70多项式回归分析特定生长率1.00——Zehra等[49]遮目鱼 Chanos chano8.00断点线性回归分析饲料效率1.690.40—Borlongan等[21]8.00断点线性回归分析饲料效率1.121.07—Borlongan等[21]0.58断点线性回归分析增重率1.900.45—Borlongan[22]0.58断点线性回归分析增重率1.261.20—Borlongan[22]真鲷 Pagrus major1.70必需氨基酸估计鱼体氨基酸含量——1.97Forster等[41]南美白对虾 Litopenaeus Vannamei0.40多项式回归分析增重率、饲料效率1.58——Jin等[50]虎纹虾 Penaeus monodon0.02多项式回归分析增重率1.40——Millamena等[51]
根据Kaushik等[52]的研究方法,将以上水产动物的Phe需求量进行Meta分析(图3)。将增重率进行标准化相应处理后,对水产动物Phe需求量与增重率进行Meta分析,得到二次多项式回归分析模型,即y饲料=-35.89x2+120.59x+0.677 9,R2=0.971 2;y饲料蛋白质=-3.952x2+40.957x-3.066 4,R2=0.939 1。以增重率为评价指标,经计算得到水产动物对Phe的需求量为占饲料的1.68%,占饲料蛋白质的5.18%。
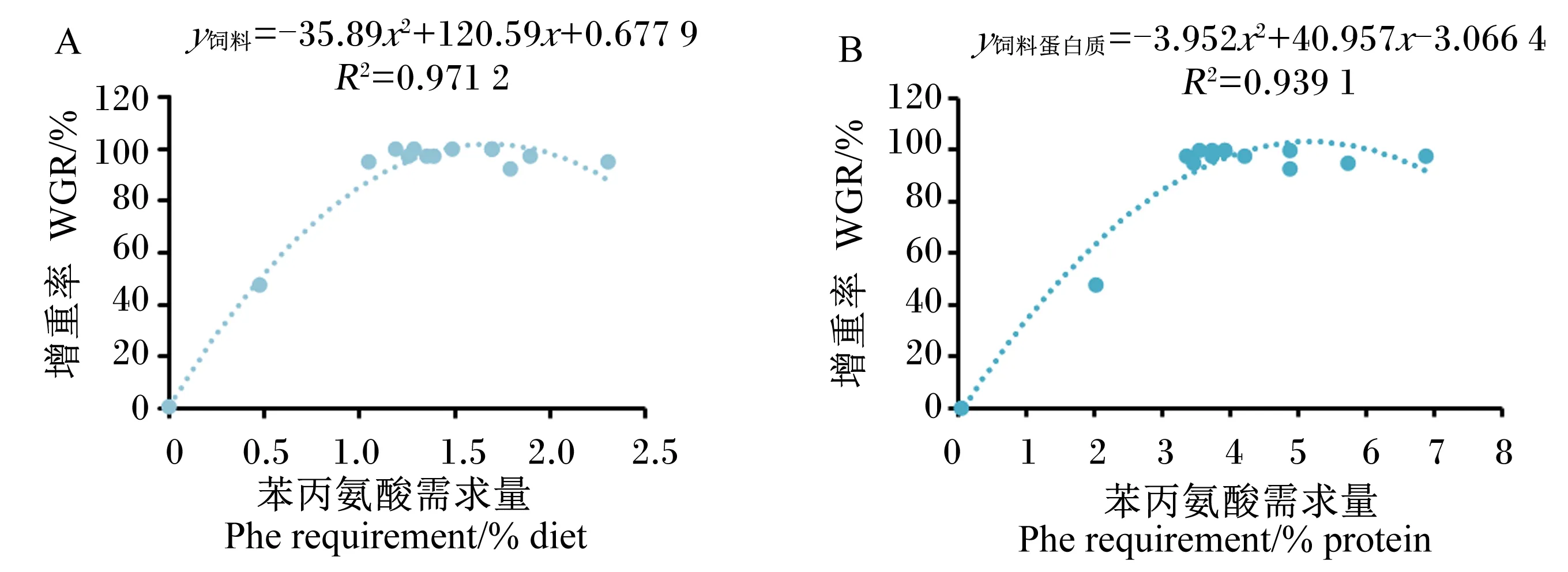
图3 水产动物苯丙氨酸需求量Meta分析
3 Phe与其他营养素的相互作用
Phe是Tyr的前体,Tyr参与多种分子的合成,且在代谢、生长、应激反应和色素沉着等方面发挥关键作用。由于Phe可以转化为Tyr,饲料中Tyr水平对Phe的需求量影响极大。研究表明,当饲料中Tyr水平为1.33%时,虹鳟(Oncorhynchusmykiss)幼鱼对Phe的需求量为0.7%;Tyr对Phe的节约替代率约为53%(以重量计),即虹鳟对Tyr和Phe的总需求量为1.5%[36]。以增重率为评价指标,当饲料中Tyr水平为0.45%时,遮目鱼对Phe的需求量为1.90%,而当饲料中Tyr水平为1.20%时,遮目鱼对Phe的需求量则下降到1.26%[22]。Khan等[19]研究表明,当露斯塔野鲮幼鱼饲料中Tyr水平为1.0%时,其对Phe的需求量为1.22%;而当饲料中缺乏Tyr时,露斯塔野鲮对Phe的需求量为2.5%。此外,在日本鳗鲡的研究中发现,当饲料中缺乏Tyr时,日本鳗鲡对Phe的需求量是2.2%,而当饲料中Tyr水平增加至2%时,日本鳗鲡对Phe的需求量是1.2%[23]。由此可见,饲料中的Tyr对Phe存在一定的节约作用。
此外,Saavedra等[53]研究表明,添加Phe和Tyr的银鲷(Diplodussargus)幼鱼对温度胁迫的存活率显著提高;同时,在饲料中补充Phe和Tyr有助于减少应激引起的骨骼畸形和死亡率。综上所述,饲料中Tyr和Phe的补充对银鲷幼鱼的生长和存活具有一定的积极作用;在温度胁迫条件下,Phe降低了银鲷幼鱼脊椎椎体的压缩率,Tyr增加了银鲷幼鱼的抵抗力,这也意味着其他代谢过程对氨基酸的需求可能高于生长所需的氨基酸。
4 Phe对水产动物蛋白质代谢及健康的影响
4.1 Phe对水产动物蛋白质沉积率的影响
Phe作为一种必需氨基酸,对水产动物的生长与代谢有着积极的促进作用。研究表明,饲料中适量的Phe可显著提高虹鳟[36]和银鲈(Bidyanusbidyanus)[46]等鱼类的生长性能。而Phe的过量或缺乏都将影响水产动物对氨基酸的吸收利用,破坏饲料的氨基酸平衡,阻碍水产动物对蛋白质的合成,进而抑制其生长[45]。在对草鱼[16]、麦瑞加拉鲮[54]、吉富罗非鱼[37]、团头鲂(Megalobramaamblycephala)[43]、卡特拉鲃(Catlacatla)[48-49]和露斯塔野鲮[19]的研究中发现,饲料Phe的过量或缺乏都将导致饲料系数的升高,引起水产动物生长性能的下降。Zehra等[18]在尼罗罗非鱼的研究中发现,增重率和蛋白质沉积率随饲料中Phe水平的增加而增加,并在1.15%时有最大值,进一步增加Phe水平则导致尼罗罗非鱼增重率和蛋白质沉积率显著下降。
值得注意的是,水产动物的种类和饲料Tyr水平等条件的不同都将导致不同的结果。孙丽慧等[16]研究表明,当饲料中Phe水平从0.82%提高到1.20%时,草鱼增重率、蛋白质沉积率和饲料效率显著升高,继续增加Phe水平后,草鱼生长性能则无显著变化。该结果与在遮目鱼[22]和大磷大马哈鱼(Oncorhynchustshawytscha)[29]等上的研究结果相似。然而,Ahmed[47]在研究麦瑞加拉鲮时发现,在适当范围内降低饲料Phe水平不会引起麦瑞加拉鲮生长性能的下降;但当饲料Phe水平过量时,则会显著降低麦瑞加拉鲮的增重率与蛋白质沉积率等生长性能指标。这可能意味着Phe过量时的抑制能力要强于缺乏时。马志英等[45]研究表明,当饲料Tyr水平为1.04%且Phe不足时,会引起异育银鲫(Carassiusauratusgibelio)幼鱼的生长性能的下降和饲料系数的升高;当提高饲料中Phe水平后,其生长性能随着饲料中Phe水平的提高而升高,超过一定范围后则导致异育银鲫生长性能显著下降。饲料中过高水平的Phe可能导致苯丙酮酸等代谢产物大量生成,并在水产动物体内聚积而产生毒性,对生长有一定的负面作用,抑制水产动物的生长[55-56],但具体机制需进一步研究。
4.2 Phe对水产动物体成分的影响
国内外关于饲料Phe水平对水产动物体成分的影响已有一些报道。Xiao等[57]研究发现,饲料Phe水平为1.30%的杂交罗非鱼(Oreochromisniloticus×Oreochromisaureus)的全鱼蛋白质和脂肪含量显著高于Phe水平为0.43%的杂交罗非鱼,而全鱼水分含量则呈下降趋势。Kim等[36]研究表明,饲料Phe水平可显著提高虹鳟全鱼粗蛋白质和粗脂肪含量。Khan等[19]对露斯塔野鲮体成分的研究表明,饲料中适宜水平的Phe可显著增加露斯塔野鲮全鱼的粗脂肪和粗灰分含量。Ren等[43]在团头鲂饲料中添加0.57%~2.04%的Phe后发现,饲料Phe水平可显著提高团头鲂全鱼水分、粗蛋白质和粗脂肪含量,但对全鱼粗灰分含量无显著影响。Phe作为一种生酮氨基酸,可随饲料Phe水平的升高而分解,为脂肪等生物大分子的合成提供足够的碳骨架,并导致全鱼粗脂肪含量的增加[58]。与露斯塔野鲮一样,吉富罗非鱼全鱼粗灰分含量随饲料Phe水平的升高而升高,当Phe水平达到1.72%后,全鱼粗灰分含量显著下降。而马志英等[45]研究则认为,饲料Phe水平对异育银鲫的全鱼体成分无显著影响。孙丽慧等[16]发现,草鱼全鱼粗蛋白质含量随饲料Phe水平的升高显著升高,而水分、粗脂肪和粗灰分含量则不随饲料Phe水平的变化而变化。由以上结果可知,不同试验的研究结果不尽相同,除与水产动物的种类有关外,还与不同生长阶段、饲料Phe水平等因素有关。
4.3 Phe对水产动物血清生化指标的影响
血清生化指标是一种判断水产动物健康程度的重要手段。研究表明,饲料中添加不同水平Phe对水产动物血清葡萄糖、胆固醇、甘油三酯含量以及谷丙转氨酶和谷草转氨酶活性等指标均可产生显著影响。Yaghoubi等[59]研究发现,当饲料中Tyr水平为0.85%时,Phe缺乏可引起体重为4.7 g的银黑鲷(Sparidentexhasta)血浆补体3(C3)、补体4(C4)含量和溶菌酶活性降低,同时引起总免疫球蛋白含量和血红细胞总数显著降低,而对其血浆总蛋白含量无显著影响。Xiao等[57]在进行饲粮中不同水平Phe(0.43%~1.91%)对杂交罗非鱼幼鱼[体重(5.63±0.04) g]血浆生化指标的影响时发现,饲粮中添加Phe可显著降低血浆谷丙转氨酶和谷草转氨酶活性,提高溶菌酶、过氧化氢酶活性和总蛋白含量。蒋明等[37]研究发现,吉富罗非鱼血清谷丙转氨酶活性随饲料Phe水平的升高先上升后下降。马志英等[45]研究表明,异育银鲫血清葡萄糖的含量在Phe水平为1.26%时达到最大值,这与饲喂不同Phe水平饲料的虹鳟血清中葡萄糖含量变化规律[60]一致;此外,当饲料Phe水平为0.73%~1.67%时,异育银鲫血清甘油三酯含量先升高后下降,且在Phe水平为1.33%时有最大值;同时,异育银鲫血清甘油三酯含量随饲料Phe水平的升高而在整体上呈下降趋势,且在Phe水平为0.89%时有最小值。Phe能够调节水产动物血清生化指标的可能原因是:当水产动物体内氨基酸维持平衡后,可充分进行蛋白质的合成,此时动物体的氧化供能则主要依靠调动总胆固醇和甘油三酯等能量物质[37]。此外,当饲料中Phe水平超出维持生长所需时,过量的氨基酸可直接参与代谢供能,也可转化为糖类或脂肪储存供能[45]。
4.4 Phe对水产动物肠道健康的影响
水产动物的肠道消化酶活性决定了其对营养物质的吸收能力。研究显示,随着饲料中Phe水平的上升,吉富罗非鱼肠道蛋白酶、脂肪酶活性呈先升高后下降的趋势,且在Phe水平为1.51%时有最大值,而肠道淀粉酶活性的变化则不显著。此外,Phe还可改善草鱼和团头鲂肠道溶菌酶、酸性磷酸酶活性和C3含量[61]。Li等[15]研究表明,饲料Phe水平为1.15%时,草鱼肠道谷胱甘肽含量和谷胱甘肽还原酶活性有最大值。Phe提高水产动物肠道消化酶活性可能是通过促进胆囊收缩素的分泌并调节胰腺消化酶的分泌实现的[62]。Feng等[63]评估了饲料Phe水平对草鱼肠道免疫反应、紧密连接蛋白基因表达水平和抗氧化相关信号通路基因表达的影响,结果表明,Phe缺乏或过量均降低了肠道溶菌酶、酸性磷酸酶活性和C3含量;当饲料Phe水平为1.15%时,可显著上调闭合蛋白-1(Occludin-1)、闭锁小带蛋白-1(ZO-1)和密封蛋白-c(Claudin-c)等紧密连接蛋白的基因表达水平,同时还可显著降低白细胞介素-8(IL-8)、肿瘤坏死因子-α(TNF-α)和核因子-κB p65(NF-κBp65)的基因表达水平。此外,当Phe水平为1.68%时,草鱼肠道超氧化物歧化酶(SOD)的基因表达水平显著降低;当Phe水平为0.91%时,草鱼肠道核因子E2相关因子2(Nrf2)的基因表达水平显著增加。综上所述,Phe改善鱼类的肠道免疫状态可能是通过调控哺乳动物雷帕霉素靶蛋白(mTOR)、NF-κB和Nrf2信号通路等途径实现的[61]。
5 小结与展望
水产养殖业绿色发展是渔业可持续发展的必然选择。饲料中添加适量的Phe不仅可以促进水产动物生长,还可改善水产动物的健康状态。目前对水产动物Phe的研究主要集中在其需求量、血清生化指标、消化酶活性和肠道健康等几个方面,且研究对象的规格主要集中在鱼苗和鱼种上。然而,由于我国水产养殖存在地理分布广泛、养殖模式丰富、设施类型复杂和养殖种类多样等特殊性,水产动物在不同生长阶段的Phe营养需求数据库还需要进一步完善。此外,关于Phe的消化吸收、转运、代谢和与其他氨基酸相互作用的分子机制的研究还需不断深入,这不仅可以为水产动物氨基酸的利用提供参考,还可进一步推动我国水产养殖业的绿色健康发展。
